Монофилия это в биологии кратко
Обновлено: 30.06.2024
(от моно. и греч. phylon — племя, род, вид), происхождение группы организмов от общего предка; один из осн. принципов эволюции органич. мира. Классич. понимание М. подразумевает возникновение таксона любого ранга от единств, родоначального вида на основе дивергенции или адаптивной радиации, при этом филогенез изображается в виде родословного древа. Широкое распространение в филогенезе разл. групп организмов явлений эволюц. параллелизма затрудняет практич. использование классич. концепции М. в систематике, поскольку границу между предковым и дочерним таксонами зачастую пересекают неск. параллельно эволюционирующих филетич. линий, общий предок к-рых существовал на более ранних этапах филогенеза. Этот частный случай М. наз. иногда параф илией. Дж. Симпсон (1960) предложил рассматривать таксон как монофилетический, если он происходит одним или неск. корнями от одного таксона более низкого ранга (т. е. тип — от класса, класс — от отряда, отряд — от семейства и т. д.). По этому определению, напр., класс млекопитающих, возникший неск. линиями от отряда терапсид, является монофилетическим. (см. ПОЛИФИЛИЯ).
Смотреть значение Монофилия в других словарях
Монофилия — (моно- + греч. phylon вид, род) концепция, согласно которой человек происходит от одного вида ископаемых наземных человекообразных обезьян.
Большой медицинский словарь
Монофилия — (от моно. и греч. phylon - племя - род), происхождение группыорганизмов от единого общего предка (не от одной пары родителей, а отоднородной группы особей). Наглядное выражение.
Большой энциклопедический словарь
Монофили́я — (Моно- + греч. phylon вид, род)
концепция, согласно которой человек происходит от одного вида ископаемых наземных человекообразных обезьян.
Медицинская энциклопедия
МОНОФИЛИЯ — МОНОФИЛИЯ (от моно. и греч. phylon — племя, род), происхождение группы организмов от общего предка; один из основных принципов эволюции органического мира.
Экологический словарь
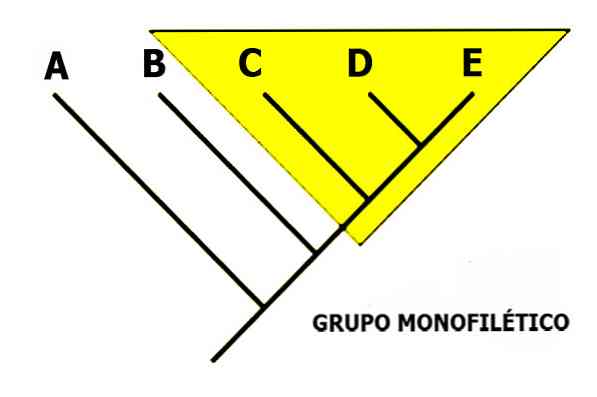
монофилетическая группа представляет собой группу видов, которые связаны друг с другом через историю уникального происхождения, то есть родовой вид и все его потомки.
Этот термин тогда обозначает естественную группу. Он противопоставляет термины полифилетический и парафилетический. Последние определяют искусственные группы как неполные (парафилетические) или для включения потомков разных предков (полифилетические).
Некоторые авторы утверждают, что, будучи единственными естественными группировками, должны приниматься только те, что относятся к морфильным группам. Эта точка зрения, однако, не единодушно разделяется всеми таксономистами и систематиками. Например, численная таксономия не дифференцирует моно, пара или полифилетические таксоны.
- 1 Классификация организмов
- 2 Таксономические школы
- 2.1 Числовая или фенетическая таксономия
- 2.2 Эволюционная таксономия
- 2.3 Филогенетическая или кладистическая таксономия
- 3.1 Расхождения
- 5.1 Кладограмма
- 5.2 Фенограмма
- 5.3 Филограмма или филетическое дерево
Классификация организмов
Таксономия - это наука, которая отвечает за классификацию живых существ. Согласно этому, организмы должны быть сгруппированы в таксоны, которые являются взаимоисключающими.
Эти таксоны, в свою очередь, сгруппированы в таксоны более высоких уровней, также взаимоисключающие для каждого из этих уровней или таксономических категорий.
В каждом таксоне организмы имеют атрибуты (признаки), на которых основаны таксономисты, чтобы указать их связь с другими организмами и тем самым разграничить биологические таксоны.
Существуют разные подходы (или школы) для оценки и взвешивания сходств (или различий), существующих между этими персонажами, и принятия соответствующих решений..
Таксономические школы
В настоящее время существует три основных таксономических школы:
Числовая или фенетическая таксономия
Предложено Р.Р. Сокаль и П.Х.А. Снит в 1963 году. Он основан на сходстве или несходстве наблюдаемых признаков, без учета предыдущих гипотез об их филогении, для классификации организмов..
Эволюционная таксономия
Это также известно как традиционная или дарвиновская таксономия. Он использует филогенетические отношения, отношения родитель-потомок (серийное потомство), а также степень эволюционных изменений для классификации организмов..
Позволяет группам быть исключенными из их родительских таксонов, считая действительными парафилетические таксоны.
Филогенетическая или кладистическая таксономия
Предложенный Вилли Хеннигом в 1966 году в его книге под названием Филогенетическая систематика. Он основан на общих производных сходствах (гомологиях) или синапоморфиях для установления эволюционных отношений между организмами.
Это основа большинства современных систем биологической классификации и стремление группировать организмы по их эволюционным связям. Только признает, насколько действительны монофилетические группы.
Противоречие между школами
В настоящее время феноменическая таксономия в строгом смысле слова используется очень немногими таксономистами, однако ее инструменты часто используются любой из двух других таксономических школ..
По словам Дэмиена Оберта, практика систематической таксономии слишком затруднена из-за глубоких различий в основах этой дисциплины.
расхождения
Существуют расхождения в типе информации, которая должна быть включена или исключена в адекватной классификации живых существ. Хотя две основные школы систематики признают эволюцию, они имеют противоположные идеи.
Кладизм утверждает, что классификация должна отражать только порядок, в котором разветвления линий происходят на древе жизни.
Эволюционизм, со своей стороны, утверждает, что степень модификации, отраженная как длина ветвей, также должна быть принята во внимание. Согласно этой школе, указанная длина будет отражать макроэволюционные разрывы.
Школа кладистов утверждает, что ни один потомок группы, содержащей их предков, не должен быть исключен. С другой стороны, эволюционная таксономия явно требует, чтобы потомки, сильно отличающиеся от своих предков, были включены в отдельные группы..
Наконец, мы можем сделать вывод, что, если мы хотим провести анализ для классификации одного или нескольких таксонов и использовать постулаты трех школ в отдельности, результаты, вероятно, будут различными.
Некоторые основные понятия
Чтобы правильно понять концепцию монофилетики, мы должны использовать определенную базовую терминологию, согласно кладистической школе, среди них:
характерлюбой наблюдаемый признак в организме, различные проявления которого называются состояниями, например, наличие волос, перьев или чешуи; географическое распределение; поведение и т. д..
Состояние персонажа: каждый из способов, которым этот персонаж может быть представлен, примитивный или производный. Например, двуногая ходьба людей является производным состоянием (персонажем), а не смещением в 4 конечностях (состояние или наследственный характер) других гоминидов..
Плезиоморфный характер: примитивный или наследственный характер, общий для всей монофилетической группы.
symplesiomorphy: плезиоморфия, разделяемая двумя или более таксонами.
Производный или апоморфный характерэто то, что возникает из предкового состояния, то есть происходит в результате трансформации персонажа в исследуемой группе. Это начало новой клады.
аутапоморфией: производный не общий символ. Он присутствует только в таксоне и часто используется в микротаксономии для дифференциации видов..
синапоморфией: апоморфия или характер, общий для двух или более видов или таксонов.
Clado (monofiletico): группа, включающая исконный вид и всех его потомков.
Гомология: состояние сходства из-за наличия общего предка.
Гомологический характер: похожие символы или с другими атрибутами, но они происходят от общего предка.
аналогия: разработка подобных структур, которые выполняют одну и ту же функцию, но их эмбриональное происхождение отличается.
homoplasy: ложное сходство, обусловленное присутствием персонажей от разных предков. Происходит путем конвергенции, параллелизма или обращения.
конвергенция: является синонимом аналогии.
параллелизм: независимая эволюция одного и того же состояния персонажа из одного и того же состояния наследственного характера.
реверсия: апоморфия, которая впоследствии теряется (возвращается к плезиоморфному состоянию) в одном из таксонов монофилетической группы.
Графические представления по таксономическим школам
кладограмма
Кладограмма является характерной схемой кладистической школы. В них выражены генеалогические филогенетические отношения, которые должны быть естественными или монофилетическими, то есть они включают общего предка и его потомков.
![]()
phenogram
Фенограммы - это диаграммы, используемые фенетической таксономией для выражения классификаций организмов. Этот тип анализа принимает три типа таксонов: монофилетические, парафилетические и полифилетические..
Хотя эти диаграммы относительно похожи на кладограммы, они не выражают филогенетические отношения, а скорее сходство или очевидное различие между организмами.
Филограмма или филетическое дерево
Филогенетические классификации, предложенные эволюционной или классической таксономической школой, используют филетические деревья. Эти диаграммы выражают генеалогические отношения предков-потомков и принимают два типа таксонов: монофилетические и парафилетические..
![]()
На этом филогенетическом древе группа завропсид , состоящая из рептилий , птиц и их общего предка, считается монофилетической .
Резюме
Определения
Монофилия в том смысле, который был наиболее популяризирован и в основном используется сегодня, был определен Вилли Хеннигом . Монофилетическая группа обозначает набор таксонов, имеющих общую синапоморфию , то есть группу, состоящую из одного предкового вида, а также всех его потомков.
Код филогенетической номенклатуры ( PhyloCode ) предлагает три типа критериев для определения таксона по набору всех потомков данной точки в рассматриваемом филогенетическом дереве : апоморфия , узел и ветвь .
Точность определения варьируется в зависимости от следующих определений:
- Согласно Филокоду , группа Короны определяется узлом: группа сохраняет только предковый вид и всех его потомков. Это самый эксклюзивный.
- Группа, определяемая в основании ветвью: группа сохраняет всех предков той же линии после кладогенеза и всех ее потомков. Это самый инклюзивный.
- Группа, определяемая синапоморфией : группа сохраняет первого предка, который приобрел синапоморфию, что позволяет определить группу и всех ее потомков. Это было бы наиболее справедливым в теории, хотя и труднодоступным на практике.
Эти определения не исключают друг друга и не являются абсолютными. Все они соответствуют основному принципу монофилии.
Происхождение термина
![]()
Монофилетическое генеалогическое древо организмов по Геккелю (1866).
Щелкните миниатюру, чтобы увеличить ее.
Голофилия и полемика
Разницу между различными терминами можно резюмировать в следующей таблице (адаптированной из Dubois 1986):
Сравнение сроков
Синтетические систематики Кладистские систематики Концепция 1 Голофилия Монофилия Концепция 2 Парафилия Парафилия Концепция 3 Полифилия Полифилия Концепция 4 (концепции 1 + 2) Монофилия Ø Концепция 5 (концепты 2 + 3) Гетерофилия Ø Полифилия
Математические свойства
В теории графов холофилия равна парафилии степени 0, а монофилия равна полифилии степени 1.
Эволюционное значение
Наличие гомологичных признаков, общих для разных таксонов, позволяет сгруппировать их под одним названием, что отражает монофилетический характер группы. Например, наличие или отсутствие ядра в клетках из эукариот или прокариот соответственно. Присутствие в группе определенных (то есть исключительных для этой группы) производных (гомологичных) признаков - или синапоморфий - позволяет индуцировать монофилию этой группы. Эукариоты образуют монофилетическую группу, потому что наличие ядра является таким производным признаком, в то время как его отсутствие является наследственной характеристикой группы прокариот (это симплезиоморфия ), поэтому последние являются парафилетическими.
Проблема монофилии — пример того, как накопление новых данных заставляет ограничить сферу приложимости некоторых, еще недавно казавшихся всеобщими принципов.
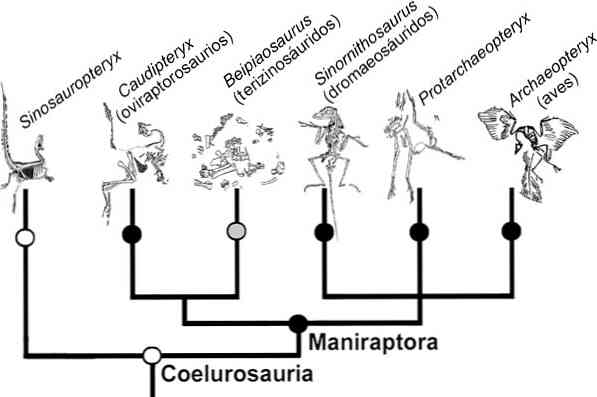
Другой пример. Класс млекопитающих характеризуется определенным набором основных признаков и свойств, отличающих их от рептилий. Этот набор признаков (постоянная теплокровность, живорождение, вскармливание детенышей молоком, развитие мозга, особенности локомоции, строение органа слуха и другие особенности) возник, как показывают палеонтологические данные, не у одного вида или группы близких видов, а в разных стволах одного пучка филогенетически близких групп рептилий, причем не одновременно (см. рис. 17.2). Тем не менее весь класс млекопитающих надо считать монофилетической группой; он возник от одного таксона того же ранга (от класса рептилий).
Сейчас много примеров того, как по мере накопления данных по морфологии, палеонтологии, особенностям образа жизни отдельных групп их приходится разделять на самостоятельные. Так было с отрядом грызунов (Glires), в который еще недавно включали в качестве подотряда зайцеобразных. Теперь нет сомнения, что сходство зайцеобразных с грызунами внешнее, конвергентное, и ныне они выделены в самостоятельные отряды Lagomorpha и Rodentia, имеющие разное происхождение. Еще недавно в один отряд хищных птиц объединяли соколов и сов — две совершенно различные по происхождению группы птиц. Как только выясняется полифилетический характер таксонов, их следует разделять.
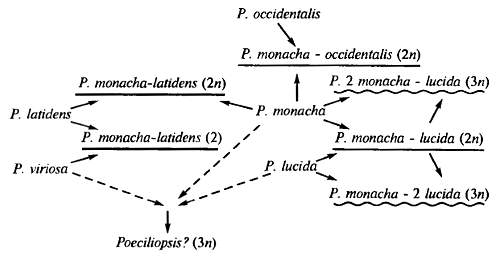
Однако проблема монофилии не исчерпывается признанием концепций широкой монофилии (происхождением одного таксона от другого такого же систематического ранга). В гл. 13 приводились некоторые из многочисленных примеров гибридогенного видообразования. Многие виды ириса, табака, полыни, костра, мятлика и других растений имеют гибридогенное происхождение (путем аллополиплоидии — гибридизации с последующим удвоением числа хромосом). Это означает, что на микроэволюционном уровне полифилия возможна. Возможность возникновения нового рода посредством гибридизации, т. е. полифилетически, доказана экспериментально (см. рис. 6.28). Сейчас известен ряд природных межродовых гибридогенных форм. Среди них рябинокизильник (Sorbocotaneaster), распространенный на юге Якутии, ячмень-клинэлимусовый гибрид (Hordeum x Clinelymus), распространенный на Памире, гибрид алычи и абрикоса — в Молдове.
Интересными примерами гибридогенного видообразования служат некоторые виды амфибий (из группы европейских зеленых лягушек) и рептилий (кавказские скальные ящерицы). В последнем случае видообразованию способствует возникновение партеногенетических форм.
![]()
Рис. 20.3. Сетчатая эволюция в гибридогенном комплексе диплоидно-триплоидных, одно- и двуполых рыб рода Poecciliopsis. Однополые формы гибридного происхождения не подчеркнуты, диплоидные гибридогенные подчеркнуты, триплолидные гиногенетические подчеркнуты волнистой линией. В скобках указана степень полиплоидности однополых форм (по Б.В. Васильеву, 1985)
Другим фактором, заставляющим по-иному взглянуть на всеобщность и обязательность проявления принципа монофилии в возникновении природных таксонов, является существование огромной и разнообразной группы лишайников. Известно, что лишайники — своеобразные комплексные организмы, которые состоят из зеленых водорослей и грибов аско- или базидиомицетов. Образующие лишайники грибы не могут жить без водорослей. Существует около 10 000 видов лишайников, выделяемые как особый класс Lichenes. При этом возникает своеобразная ситуация: одновременно существуют филогенетические системы, включающие порознь водоросли, с одной стороны, а аско- и базидиомицеты — с другой. В то же время существует особая, самостоятельная филогенетическая система для лишайников. В целом как систематическая группа организмов они имеют полифилетическое происхождение.
При обсуждении проблемы полифилии приходится вспомнить и гипотезу о симбиогенном происхождении основных групп современных организмов путем последовательного объединения аэробных бактерий и микоплазм (см. гл. 5). Это позволяет говорить о полифилетическом происхождении типов и царств живой природы.
Итак, разнообразие эволюционных путей живой природы оказывается большим, чем это предполагалось в 50-е гг. XX века: в некоторых специальных случаях в процессе эволюции могли на разных уровнях вторично возникать полифилетические таксоны.
Читайте также: