
Молекулярные механизмы фоторецепции кратко
Обновлено: 07.07.2024
восприятие света одноклеточными организмами или специализированными образованиями (фоторецепторами), содержащими светочувствительные пигменты. Ф. – одно из основных фотобиологических явлений, в котором свет выступает как источник информации. В отличие от Фотосинтеза, где энергия света используется для химической работы, в Ф. она несёт триггерную, информативную функцию, запуская сложную цепь молекулярных, мембранных и клеточных процессов. Эти процессы обеспечивают сравнительно простые формы Ф., к которым относят Фототропизм – изменение ориентации по отношению к источнику света прикрепленных животных и растений; Фототаксис – направленное движение к свету или от света свободно движущихся организмов; фотокинезис – ненаправленное увеличение или уменьшение подвижности организма в ответ на изменения уровня освещённости. Сложная и высшая форма Ф. – Зрение, осуществляемое специальными органами различной степени совершенства.
В эволюционном и сравнительно-физиологических аспектах исследование Ф. представляет большой интерес. У простейших примитивная фоторецепторная система состоит из глазного пятнышка и жгутика, т. е. рецептора и эффектора. У хламидомонады светочувствительное глазное пятнышко связано с хлоропластом (См. Хлоропласты), у эвглены – непосредственно со жгутиком. Диффузная световая чувствительность (без участия фоторецепторов) свойственна большинству беспозвоночных животных и некоторым позвоночным (отдельные виды рыб, земноводных), у некоторых она обеспечивается специальными клеточными органеллами – хроматофорами (См. Хроматофоры). Неспециализированные светочувствительные элементы могут быть разбросаны по всему телу или сконцентрированы на его поверхности и в глубине.
Зрительная Ф. совершается в фоторецепторах. Стигмы и глазки простейших, а также глазки кишечнополостных, плоских и кольчатых червей, членистоногих можно рассматривать как простейшие формы органов зрения. У моллюсков структура и функция фоторецепторной системы более сложна (у осьминога и каракатицы она, например, вполне сравнима с глазом позвоночных). Высокоспециализированные фоторецепторы в сложном (фасеточном) глазу членистоногих и в камерном глазу позвоночных образуют наиболее совершенные органы зрения. Первичные процессы зрения общие у всех животных и совершаются в светочувствительной фоторецепторной мембране зрительной клетки. Состав и молекулярная организация мембран у позвоночных и беспозвоночных в основном одинаковы. Различия, как правило, касаются способов упаковки мембран в световоспринимающих частях различных фоторецепторов. Основной светочувствительный элемент фоторецепторной мембраны – зрительный пигмент (типичный и хорошо изученный представитель – Родопсин).
В сравнительно-биохимическом аспекте исключительный интерес представляет тот факт, что производное β-каротина – Ретиналь служит хромофором всех без исключения зрительных пигментов; более того, из всех его возможных изомеров только 11-Цис-форма способна быть хромофорной частью молекулы зрительного пигмента. Т. о., удачно найденное однажды молекулярно-биохимическое решение в механизме Ф. оказалось филогенетически закрепленным. Белковая часть молекул зрительного пигмента видоспецифична. Специфичностью белка определяются, по-видимому, и различия в спектральной чувствительности колбочковых клеток в сетчатке глаза при цветовом зрении (См. Цветовое зрение). Физико-химический механизм Ф. основан на реакции фотоизомеризации ретиналя из 11-Цис-формы полностью в Транс-форму. Вследствие этой фотореакции изменяются структура (конформация) белковой части молекулы зрительного пигмента и функцией, свойства фоторецепторной мембраны. В результате в зрительной клетке происходит перемещение ионов и, возможно, изменение скоростей некоторых ферментативных реакций. Фотоиндуцированные изменения в молекуле зрительного пигмента и фоторецепторной мембране приводят в конечном счёте к возникновению в рецепторной клетке зрительного сигнала – распространяющегося фоторецепторного электрического потенциала.
Лит.: Проссер Л., Браун Ф., Сравнительная физиология животных, пер. с англ., М., 1967, гл. 12; Физиология сенсорных систем, ч. 1, Л., 1971, с. 88–119 (Руководство по физиологии); Handbook of sensory physiology, v. 7/1–v. 7/2, В., 1972.
Большая советская энциклопедия. — М.: Советская энциклопедия . 1969—1978 .
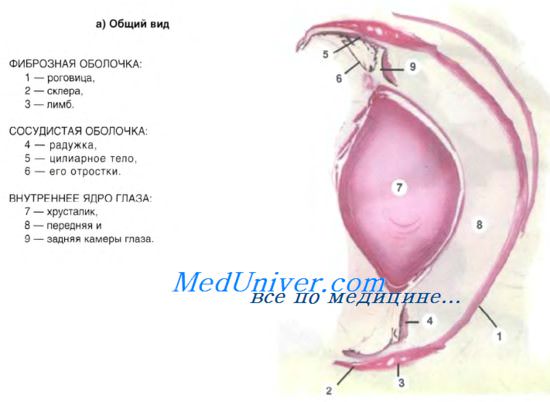
Механизм фоторецепции. Диоптрический аппарат глаза. Роговица.
Механизм фоторецепции связан с распадом молекул родопсина и йодопсина при действии световой энергии. Это запускает цепь биохимических реакций, которые сопровождаются изменением проницаемости мембран в палочках и колбочках и возникновением потенциала действия. После распада зрительного пигмента следует его ресинтез, что происходит в темноте и при наличии витамина А. Недостаток в пище витамина А может приводить к нарушению сумеречного зрения (куриная слепота). Цветовая слепота (дальтонизм) объясняется генетически обусловленным отсутствием в сетчатке одного или нескольких типов колбочек.
Возбуждение нейросенсорной клетки передается посредством центрального отростка на 2-й биполярный нейрон. Тела биполярных нейронов лежат во внутреннем ядерном слое сетчатки. В этом слое, кроме биполярных нейронов, находятся ассоциативные нейроны еще двух типов: горизонтальные и амакринные. Биполярные нейроны соединяют палочковидные и колбочковидные зрительные клетки с нейронами ганглионарного слоя. При этом колбочковидные клетки контактируют с биполярными нейронами в соотношении 1:1, тогда как с одной биполярной клеткой образуют соединения несколько палочковидных клеток.
Горизонтальные нервные клетки имеют много дендритов, с помощью которых контактируют с центральными отростками фоторецепторных клеток. Аксон горизонтальных клеток также вступает в контакт с синаптическими структурами между рецепторной и биполярной клетками. Здесь возникают множественные синапсы своеобразного типа. Передача импульсов через такой синапс и далее с помощью горизонтальных клеток может вызывать эффект латерального торможения, что увеличивает контрастность изображения объекта. Сходную роль выполняют амакринные нейроны, расположенные на уровне внутреннего сетчатого слоя. У амакринных нейронов нет аксона, но есть разветвленные дендриты. Тело нейрона играет роль синаптической поверхности.
Ганглионарные клетки образуют слой такого же названия. Это наиболее крупные нервные клетки сетчатки. Они составляют 3-й компонент нейронной цепи. Аксоны этих клеток дают слой нервных волокон, формирующих зрительный нерв.
Поддерживающие элементы в сетчатке представлены глиальными клетками (мюллеровыми волокнами) и астроцитами. Мюллеровы волокна — это крупные нейроглиальные клетки с отростками, которые располагаются вертикально по всей толщине сетчатки, оплетают нейроны сетчатки, выполняя поддерживающую и трофическую функции. Ядра клеток располагаются на уровне внутреннего ядерного слоя. Наружные отростки клеток заканчиваются многочисленными цитоплазматически-ми выростами (микроворсинками), которые формируют наружную пограничную мембрану, а внутренние — завершаются на границе со стекловидным телом (формируя внутренюю пограничную мембрану).
В сетчатке есть желтое пятно с центральной ямкой. Это — место наилучшего видения. Здесь много колбочковых нейронов. Имеется также слепое пятно, которое соответствует месту выхода зрительного нерва.
Диоптрический аппарат глаза. Роговица.
Прозрачная часть наружной фиброзной оболочки глаза называется роговицей. Ее строма развивается из мезенхимы. Передняя и задняя поверхности роговицы покрыты эпителиальными тканями. Передний эпителий роговицы — многослойный плоский неороговевающий эпителий, лежащий на базальной мембране, — является продолжением эпителия конъюнктивы. Эпителий пронизан многочисленными нервными окончаниями, увлажняется секретом слезных желез. Под эпителием располагается передняя пограничная пластинка толщиной 6-9 мкм, представляющая собой наружную часть стромы роговицы. Она участвует в защите глаза от травмы и проникновения бактерий. Ее повреждение затрудняет последующую регенерацию роговицы.
Собственное вещество роговицы состоит из параллельно лежащих коллагеновых волокон, образующих пластинки. Между ними определяется аморфное вещество, богатое гликозаминогликанами, и небольшое число отростчатых плоских клеток фибробластического дифферона. Компоненты основного вещества обеспечивают прозрачность стромы роговицы. Собственное вещество роговицы продолжается в склеру — плотную непрозрачную соединительнотканную оболочку глазного яблока, выполняющую опорную роль. Переход прозрачной роговицы в непрозрачную склеру происходит в области лимба. На границе между стромой роговицы и задним эпителием роговицы находится задняя пограничная пластинка (десцеметова мембрана) с упорядоченным расположением фибрилл.
Заднюю поверхность роговицы выстилает эпителий, состоящий из одного слоя плоских полигональных клеток нейроглиального происхождения. Эпителий участвует в питании роговицы (путем диффузии веществ из жидкости передней камеры глаза). При травме роговицы проявляет слабую регенераторную способность.
Воспалительные процессы в роговице, сопровождающиеся врастанием в нее отсутствующих в норме кровеносных сосудов, могут приводить к помутнению ее (бельмо) и полной слепоте. В настоящее время успешно проводится гомотрансплантация роговицы. Это объясняется тем, что роговица лишена кровеносных сосудов и сюда не поступают клетки крови, участвующие в отторжении трансплантата.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Исходя из общих принципов физики, биофизики и биологии, легко прийти к заключению, что первичный акт фоторецепции должен состоять в поглощении света молекулами некоего пигмента. Человек и высшие животные воспринимают свет обычной интенсивности в области примерно от 4000 до 7000 А. Ультрафиолетовый свет поглощается прозрачными тканями глаза. Инфракрасные лучи не воспринимаются сетчаткой. Если бы они воспринимались, то теплокровные животные ощущали бы сильную фоновую инфракрасную радиацию, препятствующую рецеп
ции информативных сигналов. Следовательно, пигмент или пигменты фоторецепторных клеток должны поглощать свет в видимой области спектра, т. е. быть окрашенными. Поскольку речь идет о молекулах органических соединений, отсюда следует, что это должны быть молекулы с достаточно протяженной системой л-связей — при малом числе сопряженных связей поглощается лишь ультрафиолетовое излучение.
Действительно, еще в 1933 г. Уолд открыл присутствие витамина А в зрительном пурпуре в сетчатке [107]. Структура витамина А была установлена несколько раньше [108]. Витамин А (точнее, витамин Ai), или ретинолі, имеет следующую структуру:
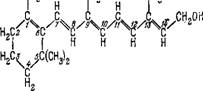
Брутто-формула ретинолаї С19Н27СН2ОН. Эта молекула представляет собой почти не измененную половину молекулы каротина (см. стр. 339). Каротиноиды являются источником витамина А, отсутствие которого приводит к слепоте.
В сетчатке позвоночных фигурирует и вторая форма витамина А—витамин А2, или ретинол2; структура которого отличается от приведенной наличием двойной связи между углеро - дами 2 и 3. Соответственно, брутто-формула ретинола2 есть С19Н25СН2ОН. Ретинолы — это спирты. В фоторецепторах ретинолы трансформированы посредством алкогольдегидрогеназы в альдегиды — в ретинальї и ретиНаль2 — и связаны с фосфолипи - дами и белками. В ретиналях конечная группа молекулы есть не =СН—СН2ОН, а =СН—СН=0, и брутто-формулы ретиналяї и ретиналя2 суть СідН27СНО и С19Н25СНО. Число сопряженных л-связей в ретиналях на одну больше, чем в ретинолах.
Таким образом, ретинали служат хромофорными группами зрительных пигментов, содержащих липопротеины, именуемые о псинами. Опсины колбочек и палочек разнятся. Соответственно возникают четыре основных вида зрительного пигмента, охарактеризованные в табл. 7.5 [109].
Хромофорные свойства ретиналей определяются протяженной системой сопряженных л-связей. Обращает на себя внимание химическая общность пигментов фотосинтеза — каротинои - дов (см. стр. 340) и зрительных хромофоров. Можно думать об
общности их эволюционного происхождения. Имеются бактерии, лишенные, конечно, зрения, но содержащие бактериородопсин, весьма сходный со зрительным пигментом.
Максимум поглощения А,, А
Ретинолі ^ФН> Ретиналь] ДФН-Н
( +Опсин I палочек
I +Опсин к колбочек
Ретинол* ^ФН> РрТИНЯЛЬо ДФН-Н
І +Опсин I палочек
I +Опсин к колбочек
Исходя из общих принципов, можно снова заключить, что вслед за поглощением света в фоторецепторе должна возникать некая фотохимическая реакция. В родопсине ретиналь образует шиффово основание в результате взаимодействия альдегидной группы ретиналя с амино-группой опсина [110, 111], а именно с е-ЫНг-группой лизила [109, 112]:
С,9Н27СН=0 + H2N—опсин —> C,9H27CH=N— опснн + Н20.
Бычнй родопсин имеет молекулярный вес около 40 000. Если считать молекулу сферической, то ее диаметр составляет 40 А.
Под действием света ретиналь отщепляется от родопсина, аналогичные процессы протекают и в трех других случаях (см. табл. 7.5). Однако этим фотохимическое превращение не исчерпывается. Одновременно происходит фотоизомеризация ретиналя.
Комплекс опсина с ретиналем возникает за счет CH = N - связи и ряда слабых взаимодействий хромофора с белком. Эти взаимодействия оптимизируются в результате структурного соответствия ретиналя и опсина. Ретиналь может существовать в виде ряда изомеров, так как двойная связь С = С образует транс- или і
В 11-ц«£-форме СНз-группа у Ci3 и водород у Сю очень сближены, и между ними возникает сильное отталкивание. Благодаря этому молекула несколько скручена, в ней нарушено плоское расположение я-связей. Тем самым уменьшена энергия сопряжения, и 11-цис-форма должна быть менее стабильной, чем полная транс-форма. Однако в комплексе с опсином проигрыш в энергии сопряжения перекрывается выигрышем в энергии взаимодействия с белком, наибольшей для 11-цмс-формы.
При выцветании пигмента на свету ретиналь отщепляется от опсина и изомеризуется в наиболее устойчивую сплошную
К Алногалйдегифоееназа, і
11-цис-решнол с > ' 1 к J Сплошной транс-ретинол Рис. 7.28. Цикл изомеризации родопсина.
Транс-фориу. Цикл цис—гране-изомеризации является обязательной частью любой известной фоторецепторной системы [109]. Этот цикл показан на рис. 7.28 [109].
В растворе ретиналь образует под действием света смесь изомеров, состав которой зависит от длины волны света и полярности растворителя [114, 115]. Полярный растворитель — этанол — стабилизует 11-цис-конфигурацию. Ее содержание в таком растворе ретиналя составляет 25—30% [116].
Установлено, что единственное действие света на ретиналь состоит именно в изомеризации 1 \-цис -> сплошной транс. Отщепление ретиналя от опсина должно рассматриваться как следствие этого процесса [117].
В фоторецепции мы встречаемся не с ротамеризацией, не с конформационной перестройкой ретиналя, а с цис.—транс-изомеризацией— повороты происходят вокруг двойных, а не единичных связей. Различие между ротамеризацией и цис—транс - изомеризацией имеет количественный характер. В первом случае для конформационного превращения нужно преодолеть барьер порядка нескольких ккал/моль (в этане 3 ккал/моль), во втором, при переходе 11-цис-ретиналя в полностью грамс-форму — около 25 ккал/моль, причем значение частотного предэкспонен - циального множителя составляет от 107 до 10" с-1 в зависимости от растворителя [118]. Соответственно изомеры ретиналя устойчивы в растворе. Разности их энергий не велики — порядка 1—3 ккал/моль.
При возбуждении молекулы поглощенным светом происходит перестройка я-электронной оболочки, и барьер внутреннего вращения снижается до величины, соизмеримой с къТ. Время жизни молекулы в синглетном возбужденном состоянии 10~9 с, время, нужное для поворота, 10-11—Ю-12 с. Квантовый выход для реакции изомеризации 1 \-цис - у полностью транс составляет 0,5—0,7 [119]. Энергия кванта света более чем достаточна для изомеризации— кванту с к 5000 А отвечает энергия 58 ккал/моль.
Значительный интерес представляет детальный механизм изомеризации. Нужно установить, через какое возбужденное состояние— синглетное или триплетное — проходит изомеризация под действием света. В теоретических работах показано, что энергетический барьер для переходов в триплетном состоянии существенно ниже, чем в синглетном [120—122]. Окончательный ответ на вопрос о роли возбужденных триплетных состояний при изомеризации хромофора в родопсине был получен в работах Розенфельда и сотрудников [123]. В этих работах был применен метод лазерного фотолиза и использована система веществ-доноров, позволявшая в результате светового возбуждения создавать высокую концентрацию триплетных состояний шиффова основания ретиналя. Действительно, оказалось, что эффективность изомеризации в триплетном состоянии очень высока. Однако время жизни триплетных состояний шиффова основания ретиналя практически такое же, как для триплетных состояний ретиналя. Это означает, что отсутствие триплетного поглощения, установленное для шиффовых оснований ретиналя [124], объясняется не малым временем жизни триплетов, а малой эффективностью интеркомбинационной конверсии. При непосредственном фотолизе этих соединений триплеты не обнаруживаются [123].
По-видимому, триплетное состояние не играет роли при изомеризации хромофора в зрительном пигменте.
Действуя светом разных длин волн на раствор родопсина в эквимолярной смеси глицерина с водой при температуре—190 °С, можно обратимо смещать равновесие изомеров, что находит свое выражение в сдвигах полосы поглощения [109]. На основе такого рода исследований Уолд построил схему последовательных превращений родопсина, показанную на рис. 7.29. Под действием
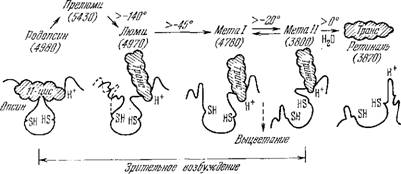
Рис. 7.29. Стадии выцветания родопсина.
Ретиналь заштрихован. В скобках указаны величины в А; температуры измеряются
В градусах Цельсия.
Света ретиналь, находящийся в комплексе с опсином, изомери- зуется, образуется прелюмиродопсин. При этом нарушается структурное соответствие ретиналя и опсина, и последний вместе со связанным ретиналем испытывает конформациониое превращение. Глобула опсина постепенно раскрывается, возникают стадии, обозначаемые как люмиродопсин и метародопсины. На заключительной стадии шиффово основание гидролизуется, и ретиналь отщепляется от опсина. При раскрытии опсина экспонируются новые химические группы, в частности, две SH-группы и группа, связывающая протон, с рК около 6,6 (вероятно, имида - зольная). Эти факты установлены с помощью биохимических и физико-химических методов [125—127].
Поглощая второй квант света, ретиналь может вновь перейти в 11-цмс-форму. Пигмент не только выцветает на свету, но может и регенерироваться. Соответствующая схема представлена на рис. 7.30 [109].
Таким образом, фоторецепция сводится к изомеризации хромофора, за которой следует изменение структуры липопро - теида — опсина. Конформациониое превращение опсина приводит в конечном счете к возникновению нервного импульса.
В сетчатке головоногих моллюсков наряду с родопсином содержится второй фоторецепторный комплекс — ретинохром. В отличие от родопсина, ретиналь в ретинохроме находится в сплошной гране-форме, изомеризующейся в 11-цнс-форму при воздействии оранжевым светом. Белковая часть ретинохрома может давать комплексы и с другими изомерами ретиналя. При освещении все они дают 11-цис-форму, отщепляющуюся от белка. В этом смысле поведение ретинохрома противоположно поведению родопсина. Можно думать, что ретинохром катализирует
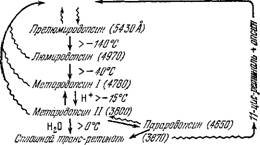
Рис. 7.30. Промежуточные стадии при выцветании и регенерации родопсина
Волнистые линии—действие света, прямые—темиовое превращение.
Изомеризацию ретиналя и, тем самым, способствует реконструкции выцветшего родопсина [128].
В работе [129] было показано, что выцветание и регенерация родопсина in vitro зависят от частоты коротких вспышек лазерного освещения. Поглощается лишь свет, поляризация которого отвечает определенной ориентации молекул родопсина. Тем самым существенно соотношение между частотой вспышек и временем поворота молекул родопсина в растворе.
Ценная информация об электронных переходах в ретинале получена в исследованиях флуоресценции (см., например, [130]). Изучение флуоресценции родопсина, внешних сегментов палочек как целостных систем и модельного соединения — N-рети- нил—опсина позволило выявить особенности переноса энергии между соседними молекулами пигмента [131]. С этими явлениями связан ряд интересных и важных проблем биомолекулярной оптики.
Различные формы зрительных пигментов характеризуются различными спектрами поглощения. Эти' различия не сводятся к указанным в табл. 7.5. Полоса поглощения хромофора существенно зависит от его взаимодействия с опсином и, следова-
Пигментная система фоторецепторов сходна с пигментной системой фотосинтеза в том отношении, что в обоих случаях у ограниченного числа пигментов встречаются самые разные спектральные чувствительности, что объясняется разнообразием их взаимодействий с белками.
В данной главе рассматриваются лишь взаимодействие каро- тиноидов со светом, их роль в фотосинтезе и фоторецепции. Однако этим не исчерпывается участие каротиноидов в биологических процессах. Способность каротиноидов к цис-транс-язоыери - зации существенна для метаболизма. Наличие большого числа - ненасыщенных двойных связей определяет легкую окисляемость этих соединений и, следовательно, участие в окислительно-восстановительных реакциях. Соответственно каротиноиды функционируют в темновой биологии. Они фигурируют в ряде тканей животных, в частности, в нейронах моллюсков, в тканях воспроизводящих органов как беспозвоночных, так и позвоночных (в том числе и у человека). Каротиноиды функциональны в окислительном метаболизме [133, 134]. Если биохимическая эволюция приводит к синтезу той или иной группы органических соединений, то они используются в живой природе многообразно— везде, где это оптимизирует существование организма.
Факты и гипотезы, кратко изложенные в настоящем параграфе, ставят ряд физических задач, еще далеких от своего решения. Структура опсина, а значит, и родопсина изучена недостаточно. Поэтому пока невозможно построение детализированной модели родопсина и его превращений на свету. Значительный интерес представила бы квантовая химия этих явлений, еще не построенная.
Наибольший интерес представляет установление связи между событиями, происходящими в родопсине, и возникновением электрических потенциалов в последовательных нейронах сетчатки, завершающееся посылкой нервного импульса в мозг. Об этом почти ничего не известно. Далее необходимо изучить строение и свойства мембран фоторецепторных клеток. Очевидно, что без такого рассмотрения нельзя даже подойти к решению указанных вопросов. Дальнейшие подробности, относящиеся к фоторецепции на молекулярном уровне, см. в [109, 135].
Начальный этап фототрансдукции связан с абсорбцией света специальными зрительными пигментами, расположенными на мембранах внутриклеточных дисков (в палочках) либо инвагинациях плазматической мембраны (в колбочках) (Рис. 37А). Пигментом палочек является родопсин,
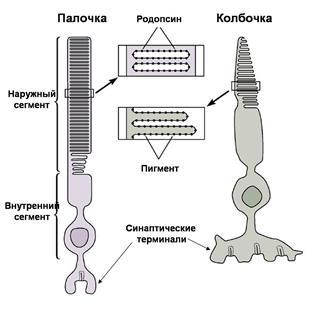 |
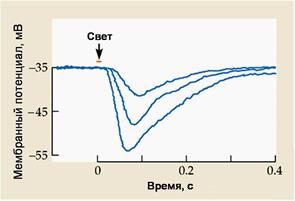
Рис. 37. Фоторецепция
А – Строение фоторецепторов. Палочки и колбочки состоят из наружного сегмента, содержащего пигмент, внутреннего сегмента и синаптической терминали, высвобождающей глутамат на клетки второго порядка. Б – Воздействие света вызывает гиперполяризационный рецепторный потенциал, амплитуда которого зависит от интенсивности света. В – Фототрансдукция в рецепторах сетчатки показана на примере палочки. В отсутствии света цГМФ-зависимые катионные каналы под действием высокой концентрации цГМФ находятся в открытом состоянии. При попадании света на фоторецептор (показан желтой стрелкой) происходит активация зрительного пигмента (родопсина в палочках), встроенного в мембраны дисков наружного сегмента фоторецептора. Активированный пигмент стимулирует G-белок (трансдуцин в палочках), который активирует цГМФ-фосфодиэстеразу. Этот фермент расщепляет цГМФ до 5’-ГМФ, что приводит к снижению внутриклеточной концентрации цГМФ и, следственно, закрытию цГМФ-зависимых катионных каналов. Уменьшение входящего тока приводит к гиперполяризации мембраны фоторецептора.
который состоит из белка опсина и светопоглощающей части – 11-цис ретиналя, производного витамина А. Под действием света 11-цис ретиналь превращается в транс-ретиналь. Каждый из трех видов колбочек имеет пигмент, предназначенный для поглощения определенной части спектра видимого света – красной, зеленой или синей. Пигменты колбочек структурно очень сходны с родопсином палочек; отличие заключается лишь в структуре белковой части (опсин), которая и определяет светочувствительность пигмента.
Установлено, что связующим звеном между процессом активации зрительных пигментов и изменением проницаемости мембраны фоторецептора является вторичный посредник - циклический гуанозинмонофосфат (цГМФ) В покое (в темноте) в цитоплазме содержится высокая концентрация цГМФ, что поддерживает цГМФ-чувствительные катионные каналы мембраны фоторецептора в открытом состоянии. Падение света на фоторецепторы приводит к снижению концентрации цГМФ, что уменьшает долю открытых каналов. На рисунке 37 В показан внутриклеточный каскад фототрансдукции.

Рис. 38. Передача сигналов между клетками сетчатки
Под действием света происходит гиперполяризация мембраны колбочки, что приводит к угнетению выброса глутамата в синапсах на биполярных клетках. ON- и OFF- биполярные клетки по-разному реагируют на уменьшение выброса глутамата. Так, ON-клетки отвечают деполяризацией мембраны и увеличением собственного выброса глутамата в синапсе с ганглиозной клеткой. Далее, в ганглиозной клетке возникает генераторный потенциал и залп потенциалов действия. Напротив, OFF-клетки отвечают гиперполяризацией и уменьшение секреции глутамата в синапсе с ганглиозной клеткой. Соответственно, происходит гиперполяризация ганглиозной клетки и прекращение генерации потенциалов действия.
ток. В OFF-клетках глутамат активирует каналы, проводящие ионы натрия.
Биполярные клетки образуют глутаматные синапсы с ганглиозными клетками. Деполяризация ON-клетки под действием света будет приводить к возбуждению ганглиозной клетки (генераторный потенциал) и генерации в ней потенциалов действия, тогда как гиперполяризация OFF-клетки при действии света будет вызывать ингибирование ганглиозной клетки (Рис. 38). Помимо фоторецепторов, биполярных и ганглиозных клеток, в сетчатке есть горизонтальные и амакринные клетки, которые участвуют в обработке и передаче сенсорной информации.
Ключевое значение в развитии представлений о зрительной рецепции сыграли работы трех выдающихся ученых. Это швед Рагнар Гранит, открывший механизм цветовосприятия, американец Халдан Хартлайн, расшифровавший процессы кодирования зрительной информации, и американец Джордж Уолд, который открыл фотохимические реакции, лежащие в основе зрительного восприятия. В 1967 году они были удостоены Нобелевской премии (рис. 39).
Рис. 39. Лауреаты Нобелевской премии 1967 года Рангар Гранит, Халдан Кеффлер Хартлайн, Джордж Уолд
Начальный этап фототрансдукции связан с абсорбцией света специальными зрительными пигментами, расположенными на мембранах внутриклеточных дисков (в палочках) либо инвагинациях плазматической мембраны (в колбочках) (Рис. 37А). Пигментом палочек является родопсин,
| |
Рис. 37. Фоторецепция
А – Строение фоторецепторов. Палочки и колбочки состоят из наружного сегмента, содержащего пигмент, внутреннего сегмента и синаптической терминали, высвобождающей глутамат на клетки второго порядка. Б – Воздействие света вызывает гиперполяризационный рецепторный потенциал, амплитуда которого зависит от интенсивности света. В – Фототрансдукция в рецепторах сетчатки показана на примере палочки. В отсутствии света цГМФ-зависимые катионные каналы под действием высокой концентрации цГМФ находятся в открытом состоянии. При попадании света на фоторецептор (показан желтой стрелкой) происходит активация зрительного пигмента (родопсина в палочках), встроенного в мембраны дисков наружного сегмента фоторецептора. Активированный пигмент стимулирует G-белок (трансдуцин в палочках), который активирует цГМФ-фосфодиэстеразу. Этот фермент расщепляет цГМФ до 5’-ГМФ, что приводит к снижению внутриклеточной концентрации цГМФ и, следственно, закрытию цГМФ-зависимых катионных каналов. Уменьшение входящего тока приводит к гиперполяризации мембраны фоторецептора.
который состоит из белка опсина и светопоглощающей части – 11-цис ретиналя, производного витамина А. Под действием света 11-цис ретиналь превращается в транс-ретиналь. Каждый из трех видов колбочек имеет пигмент, предназначенный для поглощения определенной части спектра видимого света – красной, зеленой или синей. Пигменты колбочек структурно очень сходны с родопсином палочек; отличие заключается лишь в структуре белковой части (опсин), которая и определяет светочувствительность пигмента.
Установлено, что связующим звеном между процессом активации зрительных пигментов и изменением проницаемости мембраны фоторецептора является вторичный посредник - циклический гуанозинмонофосфат (цГМФ) В покое (в темноте) в цитоплазме содержится высокая концентрация цГМФ, что поддерживает цГМФ-чувствительные катионные каналы мембраны фоторецептора в открытом состоянии. Падение света на фоторецепторы приводит к снижению концентрации цГМФ, что уменьшает долю открытых каналов. На рисунке 37 В показан внутриклеточный каскад фототрансдукции.
Рис. 38. Передача сигналов между клетками сетчатки
Под действием света происходит гиперполяризация мембраны колбочки, что приводит к угнетению выброса глутамата в синапсах на биполярных клетках. ON- и OFF- биполярные клетки по-разному реагируют на уменьшение выброса глутамата. Так, ON-клетки отвечают деполяризацией мембраны и увеличением собственного выброса глутамата в синапсе с ганглиозной клеткой. Далее, в ганглиозной клетке возникает генераторный потенциал и залп потенциалов действия. Напротив, OFF-клетки отвечают гиперполяризацией и уменьшение секреции глутамата в синапсе с ганглиозной клеткой. Соответственно, происходит гиперполяризация ганглиозной клетки и прекращение генерации потенциалов действия.
ток. В OFF-клетках глутамат активирует каналы, проводящие ионы натрия.
Биполярные клетки образуют глутаматные синапсы с ганглиозными клетками. Деполяризация ON-клетки под действием света будет приводить к возбуждению ганглиозной клетки (генераторный потенциал) и генерации в ней потенциалов действия, тогда как гиперполяризация OFF-клетки при действии света будет вызывать ингибирование ганглиозной клетки (Рис. 38). Помимо фоторецепторов, биполярных и ганглиозных клеток, в сетчатке есть горизонтальные и амакринные клетки, которые участвуют в обработке и передаче сенсорной информации.
Ключевое значение в развитии представлений о зрительной рецепции сыграли работы трех выдающихся ученых. Это швед Рагнар Гранит, открывший механизм цветовосприятия, американец Халдан Хартлайн, расшифровавший процессы кодирования зрительной информации, и американец Джордж Уолд, который открыл фотохимические реакции, лежащие в основе зрительного восприятия. В 1967 году они были удостоены Нобелевской премии (рис. 39).
Рис. 39. Лауреаты Нобелевской премии 1967 года Рангар Гранит, Халдан Кеффлер Хартлайн, Джордж Уолд
Читайте также: