
Микротрубочки это в биологии кратко
Обновлено: 05.07.2024
Общая характеристика микротрубочек. К обязательным компонентам цитоскелета относятся микротрубочки (рис. 265), нитчатые неветвящиеся структуры, толщиной 25 нм, состоящие из белков-тубулинов и ассоциированных с ними белков. Тубулины при полимеризации образуют полые трубки (микротрубочки), длина которых может достигать нескольких мкм, а самые длинные микротрубочки встречаются в составе аксонемы хвостов спермиев.
Микротрубочки располагаются в цитоплазме интерфазных клеток поодиночке, небольшими рыхлыми пучками, или в виде плотноупакованных образований в составе центриолей, базальных телец в ресничках и жгутиках. При делении клеток большая часть микротрубочек клетки входит в состав веретена деления.
По строению микротрубочки представляют собой длинные полые цилиндры с внешним диаметром 25 нм (рис. 266). Стенка микротрубочек состоит из полимеризованных молекул белка тубулина. При полимеризации молекулы тубулина образуют 13 продольных протофиламентов, которые скручиваются в полую трубку (рис. 267). Размер мономера тубулина составляет около 5 нм, равного толщине стенки микротрубочки, в поперечном сечении которой видны 13 глобулярных молекул.
Молекула тубулина представляет собой гетеродимер, состоящий из двух разных субъедниц, из a–тубулина и b– тубулина, которые при ассоциации образуют собственно белок тубулин, изначально поляризованный. Обе субъединицы мономера тубулина связаны с ГТФ, однако на a-субъдинице ГТФ не подвергается гидролизу, в отличие от ГТФ на b-субъединице, где при полимеризации происходит гидролиз ГТФ до ГДФ. При полимеризации молекулы тубулина объединяются таким образом, что с b-субъединицей одного белка ассоциирует a–субъединица следующего белка и т.д. Следовательно, отдельные протофибриллы возникают как полярные нити, и соответственно вся микротрубочка тоже является полярной структурой, имеющей быстро растущий (+)-конец и медленно растущий (-) конец (рис. 268).
При достаточной концентрации белка полимеризация происходит спонтанно. Но при спонтанной полимеризации тубулинов происходит гидролиз одной молекулы ГТФ, связанной с b-тубулином. Во время наращивания длины микротрубочки связывание тубулинов происходит с большей скоростью на растущем (+)-конце. Но при недостаточной концентрации тубулина микротрубочки могут разбираться с обоих концов. Разборке микротрубочек способствует понижение температуры и наличие ионов Са ++.
Микротрубочки являются очень динамичными структурами, которые могут достаточно быстро возникать и разбираться. В составе выделенных микротрубочек обнаруживаются ассоциированные с ними дополнительные белки, т.н. МАР-белки (МАР- microtubule accessory proteins). Эти белки, стабилизируя микротрубочки, ускоряют процесс полимеризации тубулина (рис. 269).
Роль цитоплазматических микротрубочек сводится к выполнению двух функций: скелетной и двигательной. Скелетная, каркасная, роль заключается в том, что расположение микротрубочек в цитоплазме стабилизирует форму клетки; при растворении микротрубочек клетки, имевшие сложную форму, стремятся приобрести форму шара. Двигательная роль микротрубочек заключается не только в том, что они создают упорядоченную, векторную, систему движения. Микротрубочки цитоплазмы в ассоциации со специфическими ассоциированными моторными белками образуют АТФ-азные комплексы, способные приводить в движение клеточные компоненты.
Практически во всех эукариотических клетках в гиалоплазме можно видеть длинные неветвящиеся микротрубочки. В больших количествах они обнаруживаются в цитоплазматических отростках нервных клеток, в отростках меланоцитов, амеб и других изменяющих свою форму клетках (рис. 270). Они могут быть выделены сами или же можно выделить их образующие белки: это те же тубулины со всеми их свойствами.
Центры организации микротрубочек. Рост микротрубочек цитоплазмы происходит полярно: наращивается (+)-конец микротрубочки. Время жизни микротрубочек очень коротка, поэтому постоянно происходит образование новых микротрубочек. Процесс начала полимеризации тубулинов, нуклеация, происходит в четко ограниченных участках клетки, в т.н. центрах организации микротрубочек (ЦОМТ). В зонах ЦОМТ происходит закладка коротких микротрубочек, обращенных своими (-)-концами к ЦОМТ. Считается, что в зонах ЦОМТ (--)-концы заблокированы специальными белками, предотвращающими или ограничивающими деполимеризацию тубулинов. Поэтому при достаточном количестве свободного тубулина будет происходить наращивание длины микротрубочек, отходящих от ЦОМТ. В качестве ЦОМТ в клетках животных участвуют главным образом клеточные центры, содержащие центриоли, о чем будет сказано далее. Кроме того в качестве ЦОМТ может служить ядерная зона, и во время митоза полюса веретена деления.

Одним из назначений микротрубочек цитоплазмы заключается в создании эластичного, но одновременно устойчивого внутриклеточного скелета, необходимого для поддержания формы клетки. У дисковидных по форме эритроцитов амфибий по периферии клетки лежит жгут циркулярно уложенных микротрубочек; пучки микротрубочек характерны для различных выростов цитоплазмы (аксоподии простейших, аксоны нервных клеток и т.д.).
Роль микротрубочек заключается в образовании каркаса для поддержания клеточного тела, для стабилизации и укрепления клеточных выростов. Кроме того, микротрубочки участвуют в процессах роста клеток. Так, у растений в процессе растяжения клеток, когда за счет увеличения центральной вакуоли происходит значительный рост объема клеток, большие количества микротрубочек появляются в периферических слоях цитоплазмы. В этом случае микротрубочки, так же как и растущая в это время клеточная стенка, как бы армируют, механически укрепляют цитоплазму.
Создавая внутриклеточный скелет, микротрубочки являются факторами ориентированного движения внутриклеточных компонентов, задавая своим расположением пространства для направленных потоков разных веществ и для перемещения крупных структур. Так, в случае меланофоров (клетки, содержащие пигмент меланин) рыб при росте клеточных отростков гранулы пигмента передвигаются вдоль пучков микротрубочек.
В аксонах живых нервных клеток можно наблюдать перемещение различных мелких вакуолей и гранул, которые двигаются как от тела клетки к нервному окончанию (антероградный транспорт), так и в противоположном направлении (ретроградный транспорт).
Были выделены белки, ответственные за движение вакуолей. Один из них кинезин, белок с молекулярным весом около 300 тыс.
Существует целое семейство кинезинов. Так, цитозольные кинезины участвуют в транспорте по микротрубочкам везикул, лизосом и других мембраных органелл. Многие из кинезинов связываются специфически со своими грузами. Так некоторые участвуют в переносе только митохондрий, другие – только синаптических пузырьков. Кинезины связываются с мембранами через мембранные белковые комплексы – кинектины. Кинезины веретена деления участвуют в образовании этой структуры и в расхождении хромосом.
За ретроградный транспорт в аксоне отвечает другой белок – цитоплазматический динеин (рис. 275). Он состоит из двух тяжелых цепей – головок, взаимодействующих с микротрубочками, нескольких промежуточных и легких цепей, которые связываются с мембранными вакуолями. Цитоплазматический динеин является моторным белком, переносящим грузы к минус-концу микротрубочек. Динеины также делятся на два класса: цитозольные – участвующие в переносе вакуолей и хромосом, и аксонемные – отвечающие за движение ресничек и жгутиков.
Цитоплазматические динеины и кинезины были обнаружены практически во всех типах клеток животных и растений.
Таким образом, и в цитоплазме движение осуществляется по принципу скользящих нитей, только вдоль микротрубочек перемещаются не нити, а короткие молекулы – движетели, связанные с перемещающимися клеточными компонентами. Сходство с актомиозиновым комплексом этой системы внутриклеточного транспорта заключается в том, что образуется двойной комплекс (микротрубочка + движетель), обладающий высокой АТФ-азной активностью.
Как видно, микротрубочки образуют в клетке радиально расходящиеся поляризованные фибриллы, (+)-концы которых направлены от центра клетки к периферии. Наличие же (+) и (-)-направленных моторные белков (кинезинов и динеинов) создает возможность для переноса в клетке её компонентов как от периферии к центру (эндоцитозные вакуоли, рециклизация вакуолей ЭР и аппарата Гольджи и др), так и от центра к периферии (вакуоли ЭР, лизосомы, секреторные вакуоли и др) (рис. 276). Такая полярность транспорта создается за счет организации системы микротрубочек, возникающих в центрах их организации, в клеточном центре.
Общая характеристика микротрубочек. К обязательным компонентам цитоскелета относятся микротрубочки (рис. 265), нитчатые неветвящиеся структуры, толщиной 25 нм, состоящие из белков-тубулинов и ассоциированных с ними белков. Тубулины при полимеризации образуют полые трубки (микротрубочки), длина которых может достигать нескольких мкм, а самые длинные микротрубочки встречаются в составе аксонемы хвостов спермиев.
Микротрубочки располагаются в цитоплазме интерфазных клеток поодиночке, небольшими рыхлыми пучками, или в виде плотноупакованных образований в составе центриолей, базальных телец в ресничках и жгутиках. При делении клеток большая часть микротрубочек клетки входит в состав веретена деления.
По строению микротрубочки представляют собой длинные полые цилиндры с внешним диаметром 25 нм (рис. 266). Стенка микротрубочек состоит из полимеризованных молекул белка тубулина. При полимеризации молекулы тубулина образуют 13 продольных протофиламентов, которые скручиваются в полую трубку (рис. 267). Размер мономера тубулина составляет около 5 нм, равного толщине стенки микротрубочки, в поперечном сечении которой видны 13 глобулярных молекул.
Молекула тубулина представляет собой гетеродимер, состоящий из двух разных субъедниц, из a–тубулина и b– тубулина, которые при ассоциации образуют собственно белок тубулин, изначально поляризованный. Обе субъединицы мономера тубулина связаны с ГТФ, однако на a-субъдинице ГТФ не подвергается гидролизу, в отличие от ГТФ на b-субъединице, где при полимеризации происходит гидролиз ГТФ до ГДФ. При полимеризации молекулы тубулина объединяются таким образом, что с b-субъединицей одного белка ассоциирует a–субъединица следующего белка и т.д. Следовательно, отдельные протофибриллы возникают как полярные нити, и соответственно вся микротрубочка тоже является полярной структурой, имеющей быстро растущий (+)-конец и медленно растущий (-) конец (рис. 268).
При достаточной концентрации белка полимеризация происходит спонтанно. Но при спонтанной полимеризации тубулинов происходит гидролиз одной молекулы ГТФ, связанной с b-тубулином. Во время наращивания длины микротрубочки связывание тубулинов происходит с большей скоростью на растущем (+)-конце. Но при недостаточной концентрации тубулина микротрубочки могут разбираться с обоих концов. Разборке микротрубочек способствует понижение температуры и наличие ионов Са ++.
Микротрубочки являются очень динамичными структурами, которые могут достаточно быстро возникать и разбираться. В составе выделенных микротрубочек обнаруживаются ассоциированные с ними дополнительные белки, т.н. МАР-белки (МАР- microtubule accessory proteins). Эти белки, стабилизируя микротрубочки, ускоряют процесс полимеризации тубулина (рис. 269).
Роль цитоплазматических микротрубочек сводится к выполнению двух функций: скелетной и двигательной. Скелетная, каркасная, роль заключается в том, что расположение микротрубочек в цитоплазме стабилизирует форму клетки; при растворении микротрубочек клетки, имевшие сложную форму, стремятся приобрести форму шара. Двигательная роль микротрубочек заключается не только в том, что они создают упорядоченную, векторную, систему движения. Микротрубочки цитоплазмы в ассоциации со специфическими ассоциированными моторными белками образуют АТФ-азные комплексы, способные приводить в движение клеточные компоненты.
Практически во всех эукариотических клетках в гиалоплазме можно видеть длинные неветвящиеся микротрубочки. В больших количествах они обнаруживаются в цитоплазматических отростках нервных клеток, в отростках меланоцитов, амеб и других изменяющих свою форму клетках (рис. 270). Они могут быть выделены сами или же можно выделить их образующие белки: это те же тубулины со всеми их свойствами.
Центры организации микротрубочек. Рост микротрубочек цитоплазмы происходит полярно: наращивается (+)-конец микротрубочки. Время жизни микротрубочек очень коротка, поэтому постоянно происходит образование новых микротрубочек. Процесс начала полимеризации тубулинов, нуклеация, происходит в четко ограниченных участках клетки, в т.н. центрах организации микротрубочек (ЦОМТ). В зонах ЦОМТ происходит закладка коротких микротрубочек, обращенных своими (-)-концами к ЦОМТ. Считается, что в зонах ЦОМТ (--)-концы заблокированы специальными белками, предотвращающими или ограничивающими деполимеризацию тубулинов. Поэтому при достаточном количестве свободного тубулина будет происходить наращивание длины микротрубочек, отходящих от ЦОМТ. В качестве ЦОМТ в клетках животных участвуют главным образом клеточные центры, содержащие центриоли, о чем будет сказано далее. Кроме того в качестве ЦОМТ может служить ядерная зона, и во время митоза полюса веретена деления.
Одним из назначений микротрубочек цитоплазмы заключается в создании эластичного, но одновременно устойчивого внутриклеточного скелета, необходимого для поддержания формы клетки. У дисковидных по форме эритроцитов амфибий по периферии клетки лежит жгут циркулярно уложенных микротрубочек; пучки микротрубочек характерны для различных выростов цитоплазмы (аксоподии простейших, аксоны нервных клеток и т.д.).
Роль микротрубочек заключается в образовании каркаса для поддержания клеточного тела, для стабилизации и укрепления клеточных выростов. Кроме того, микротрубочки участвуют в процессах роста клеток. Так, у растений в процессе растяжения клеток, когда за счет увеличения центральной вакуоли происходит значительный рост объема клеток, большие количества микротрубочек появляются в периферических слоях цитоплазмы. В этом случае микротрубочки, так же как и растущая в это время клеточная стенка, как бы армируют, механически укрепляют цитоплазму.
Создавая внутриклеточный скелет, микротрубочки являются факторами ориентированного движения внутриклеточных компонентов, задавая своим расположением пространства для направленных потоков разных веществ и для перемещения крупных структур. Так, в случае меланофоров (клетки, содержащие пигмент меланин) рыб при росте клеточных отростков гранулы пигмента передвигаются вдоль пучков микротрубочек.
В аксонах живых нервных клеток можно наблюдать перемещение различных мелких вакуолей и гранул, которые двигаются как от тела клетки к нервному окончанию (антероградный транспорт), так и в противоположном направлении (ретроградный транспорт).
Были выделены белки, ответственные за движение вакуолей. Один из них кинезин, белок с молекулярным весом около 300 тыс.
Существует целое семейство кинезинов. Так, цитозольные кинезины участвуют в транспорте по микротрубочкам везикул, лизосом и других мембраных органелл. Многие из кинезинов связываются специфически со своими грузами. Так некоторые участвуют в переносе только митохондрий, другие – только синаптических пузырьков. Кинезины связываются с мембранами через мембранные белковые комплексы – кинектины. Кинезины веретена деления участвуют в образовании этой структуры и в расхождении хромосом.
За ретроградный транспорт в аксоне отвечает другой белок – цитоплазматический динеин (рис. 275). Он состоит из двух тяжелых цепей – головок, взаимодействующих с микротрубочками, нескольких промежуточных и легких цепей, которые связываются с мембранными вакуолями. Цитоплазматический динеин является моторным белком, переносящим грузы к минус-концу микротрубочек. Динеины также делятся на два класса: цитозольные – участвующие в переносе вакуолей и хромосом, и аксонемные – отвечающие за движение ресничек и жгутиков.
Цитоплазматические динеины и кинезины были обнаружены практически во всех типах клеток животных и растений.
Таким образом, и в цитоплазме движение осуществляется по принципу скользящих нитей, только вдоль микротрубочек перемещаются не нити, а короткие молекулы – движетели, связанные с перемещающимися клеточными компонентами. Сходство с актомиозиновым комплексом этой системы внутриклеточного транспорта заключается в том, что образуется двойной комплекс (микротрубочка + движетель), обладающий высокой АТФ-азной активностью.
Как видно, микротрубочки образуют в клетке радиально расходящиеся поляризованные фибриллы, (+)-концы которых направлены от центра клетки к периферии. Наличие же (+) и (-)-направленных моторные белков (кинезинов и динеинов) создает возможность для переноса в клетке её компонентов как от периферии к центру (эндоцитозные вакуоли, рециклизация вакуолей ЭР и аппарата Гольджи и др), так и от центра к периферии (вакуоли ЭР, лизосомы, секреторные вакуоли и др) (рис. 276). Такая полярность транспорта создается за счет организации системы микротрубочек, возникающих в центрах их организации, в клеточном центре.
С помощью электронного микроскопа в цитоплазме эукариот можно увидеть фибриллярную сеть, функции которой связаны с движением внутриклеточного содержимого, перемещением самой клетки, а также в совокупности с другими структурами поддерживается форма клетки.
Одними из таких фибрилл являются микротрубочки (обычно длиной от нескольких микрометров до нескольких миллиметров), представляющие собой длинные тонкие цилиндры (диаметром около 25 нм) с полостью внутри. Их относят к органоидам клетки.
Стенки микротрубочек состоят из спирально упакованных субъединиц белка тубулина, состоящего из двух частей, то есть представляющего собой димер.

Соседние трубочки могут быть связаны между собой выступами своих стенок.
Данный клеточный органоид относится к динамическим структурам, так может нарастать и распадаться (полимеризуется и деполимеризуется). Рост происходит за счет добавления новых тубулиновых субъединиц с одного конца (плюс), а разрушение – с другого (минус-конец). То есть микротрубочки полярны.
В животных клетках (а также у многих простейших) центрами организации микротрубочек являются центриоли. Они сами состоят из девяти триплетов укороченных микротрубочек и располагаются около ядра. От центриолей трубочки радиально расходятся, то есть растут к периферии клетки. У растений центрами организации выступают другие структуры.
Из микротрубочек состоит веретено деления, которое осуществляет расхождение хроматид или хромосом при митозе или мейозе. Из них состоят базальные тельца, лежащие в основании ресничек и жгутиков. Движение веретена, ресничек и жгутиков происходит за счет скольжения трубочек.

Похожей функцией является перемещение ряда клеточных органоидов и частиц (например, секреторных пузырьков, образующихся в аппарате Гольджи, лизосом, даже митохондрий). При этом микротрубочки играют роль своеобразных рельсов. Специальные моторные белки одним своим концом крепятся к трубочкам, а другим — к органеллам. За счет их движения вдоль трубочек происходит транспорт органелл. При этом одни моторные белки двигаются только от центра к периферии (кинезины), другие (динеины) — от периферии к центру.
Микротрубочки за счет своей жесткости участвуют в формировании опорной системы клетки — цитоскелета. Определяют форму клетки.
Сборка и разборка микротрубочек, а также транспорт по ним идет с затратой энергии.
Расположение микротрубочек
В функциональном отношении периферическая опорно-сократимая фибриллярная система клетки находится в теснейшем взаимодействии с системой периферических микротрубочек. Это дает нам основание рассматривать последние как часть субмембранной системы клетки.
Белки микротрубочек
Система микротрубочек является вторым компонентом опорно-сократимого аппарата, находящаяся, как правило, в тесном контакте с микрофибриллярным компонентом.
Стенки микротрубочек образованы в поперечнике чаще всего 13 димерными глобулами белка, каждая глобула состоит из α- и β-тубулинов (рис. 6). Последние в большинстве микротрубочек расположены в шахматном порядке. Тубулин составляет 80% белков содержащихся в микротрубочках.
Остальные 20% приходятся на долю высокомолекулярных белков МАР1, МАР2 и низкомолекулярного тау-фактора. МАР-белки (microtubule-associated proteins- белки, связанные с микротрубочками) и тау-фактор представляют собой компоненты, необходимые для полимеризации тубулина. В их отсутствие самосборка микротрубочек путем полимеризации тубулина крайне затруднена и образующиеся микротрубочки сильно отличаются от нативных.
Микротрубочки — очень лабильная структура, так, микротрубочки теплокровных животных, как правило, разрушаются на холоде.
Существуют и холодоустойчивые микротрубочки, например в нейронахцентральной нервной системы позвоночных их количество варьирует от 40 до 60%. Термостабильные и термолабильные микротрубочки не различаются по свойствам входящего в их состав тубулина; по-видимому, эти отличия определяются добавочными белками.
Функции микротрубочек
Так же как и микрофибриллы, микротрубочки подвержены функциональной изменчивости.
Какие функции выполняют микротрубочки?
Для них характерны самосборка и саморазборка, причем разборка происходит до тубулиновых димеров. Соответственно микротрубочки могут быть представлены большим или меньшим количеством в связи с преобладанием процессов либо саморазборки, либо самосборки микротрубочек из фонда глобулярного тубулина гиалоплазмы.
Интенсивные процессы самосборки микротрубочек обычно приурочены к местам крепления клеток к субстрату, т. е. к местам усиленной полимеризации фибриллярного актина из глобулярного актина гиалоплазмы.
Такая корреляция степени развития этих двух механохимических систем не случайна и отражает их глубокую функциональную взаимосвязь в целостной опорно-сократимой и транспортной системе клетки.
В эту группу органоидов входят рибосомы, микротрубочки и микрофиламенты, клеточный центр.
Рибосома
Рибосомы (рис. 1) присутствуют в клетках как эукариот, так и прокариот, поскольку выполняют важную функцию в биосинтезе белков.
В каждой клетке имеются десятки, сотни тысяч (до нескольких миллионов) этих мелких округлых органоидов. Это округлая рибонуклеопротеиновая частица. Диаметр ее составляет 20—30 нм. Состоит рибосома из большой и малой субъединиц, которые объединяются в присутствии нити м-РНК (матричной, или информационной, РНК). Комплекс из группы рибосом, объединенных одной молекулой м-РНК наподобие нитки бус, называется полисомой. Эти структуры либо свободно расположены в цитоплазме, либо прикреплены к мембранам гранулярной ЭПС (в обоих случаях на них активно протекает синтез белка).

Рис.1.Схема строения рибосомы, сидяшей на мембране эндоплазматической сети: 1 — малая субъединииа; 2 иРНК; 3 — аминоацил-тРНК; 4 — аминокислота; 5 — большая субъединица; 6 — — мембрана эндоплазматической сети; 7 — синтезируемая полипептидная цепь
Полисомы гранулярной ЭПС образуют белки, выводимые из клетки и используемые для нужд всего организма (например, пищеварительные ферменты, белки женского грудного молока).
Кроме этого, рибосомы присутствуют на внутренней поверхности мембран митохондрий, где также принимают активное участие в синтезе белковых молекул.
Микротрубочки
Это трубчатые полые образования, лишенные мембраны. Внешний диаметр составляет 24 нм, ширина просвета — 15 нм, толщина стенки — около 5 нм. В свободном состоянии представлены в цитоплазме, также являются структурными элементами жгутиков, центриолей, веретена деления, ресничек.
Микротрубочки построены из стереотипных белковых субъединиц путем их полимеризации. В любой клетке процессы полимеризации идут параллельно процессам деполимеризации.
Причем соотношение их определяется количеством микротрубочек. Микротрубочки имеют различную устойчивость к разрушающим их факторам, например, к колхицину (это химическое вещество, вызывающее деполимеризацию). Функции микротрубочек:
1) являются опорным аппаратом клетки;
2) определяют формы и размеры клетки;
3) являются факторами направленного перемещения внутриклеточных структур.
Микрофиламенты
Это тонкие и длинные образования, которые обнаруживаются по всей цитоплазме.
Иногда образуют пучки. Виды микро-филаментов:
1) актиновые. Содержат сократительные белки (актин), обеспечивают клеточные формы движения (например, амебоидные), играют роль клеточного каркаса, участвуют в организации перемещений органелл и участков цитоплазмы внутри клетки;
2) промежуточные (толщиной 10 нм). Их пучки обнаруживаются по периферии клетки под плазмалеммой и по окружности ядра.
Выполняют опорную (каркасную) роль.
Микротрубочки
В разных клетках (эпителиальных, мышечных, нервных, фибробластах) построены из разных белков.
Микрофиламенты, как и микротрубочки, построены из субъединиц, поэтому их количество определяется соотношением процессов полимеризации и деполимеризации.
Клетки всех животных, некоторых грибов, водорослей, высших растений характеризуются наличием клеточного центра.
Клеточный центр обычно располагается рядом с ядром.
Он состоит из двух центриолей, каждая из которых представляет собой полый цилиндр диаметром около 150 нм, длиной 300—500 нм.
Центриоли расположены взаимоперпендикулярно.
Стенка каждой центриоли образована 27 микротрубочками, состоящими из белка тубулина. Микротрубочки сгруппированы в 9 триплетов.
Из центриолей клеточного центра во время деления клетки образуются нити веретена деления.
Центриоли поляризуют процесс деления клетки, чем достигается равномерное расхождение сестринских хромосом (хроматид) в анафазе митоза.
Так называются непостоянные компоненты в клетке, присутствующие в основном веществе цитоплазмы в виде зерен, гранул или капелек. Включения могут быть окружены мембраной или же не окружаются ею.
В функциональном отношении выделяют три вида включений: запасные питательные вещества (крахмал, гликоген, жиры, белки), секреторные включения (вещества, характерные для железистых клеток, продуцируемые ими, — гормоны желез внутренней секреции и т. п.) и включения специального назначения (в узкоспециализированных клетках, например гемоглобин в эритроцитах).
Реснички и жгутики
Характерная для ресничек инфузорий организация постоянных тубулин-динеиновых механохимических комплексов с двумя центральными и девятью периферическими парами микротрубочек имеет широкое распространение и в специализированных клетках метазойных животных (реснички и жгутики клеток ресничных эпителиев, жгутики сперматозоидов и др.). Однако такой принцип построения не является единственной конструктивной формой организации постоянных тубулин-динеиновых систем.
Микротрубочки, их строение и функции
Проведенный в последнее время детальный сравнительно-цитологический анализ организации жгутиков сперматозоидов у разных многоклеточных животных показал возможность существенных изменений стандартной формулы 9 + 2 даже у близкородственных животных.
В жгутиках сперматозоидов некоторых групп животных две центральные микротрубочки могут отсутствовать, а их роль выполняют цилиндры из электронно-плотного вещества. Среди низших многоклеточных (турбеллярии и близкие к ним группы) подобного рода модификации распределены у отдельных видов животных мозаично и, вероятно, полифилетичны по своему происхождению, хотя у всех этих видов образуются сходные морфологические структуры.
Принцип работы тубулин-динеинового комплекса
В настоящее время имеется несколько гипотез, объясняющих принцип работы тубулин-динеиновой механохимической системы.
Необходимо отметить и сходные признаки в свойствах основных сократимых белков динеина и миозина, с одной стороны, и тубулина и актина — с другой. Для динеина и миозина это близкие молекулярные веса и наличие АТФазной активности. Для тубулина и актина помимо сходства молекулярных весов характерны близкие аминокислотный состав и первичная структура белковых молекул.
Совокупность перечисленных признаков структурно-биохимической организации актин-миозиновой и тубулин-динеиновой систем позволяет предполагать, что они развились из одной механохимической системы первичных эукариотных клеток и сложились в результате прогрессивного усложнения их организации.
Взаимодействие актин-миозиновой и тубулин-динеиновой комплекса
Актин-миозиновая и тубулин-динеиновая комплексы, как правило, в большинстве эукариотных клеток объединяются при функционировании в одну систему.
Так, например, в динамичном субмембранном аппарате культивируемых in vitro клеток присутствуют обе механохимические системы: и актин-миозиновая, и тубулин-динеиновая. Возможно, что это связано с особой ролью микротрубочек как организующих и направляющих скелетных образований клетки. С другой стороны, наличие двух аналогичных систем может повышать пластичность сократимых внутриклеточных структур, тем более что регуляция работы актин-миозиновой системы принципиально отличается от регуляции работы динеин-тубулиновой системы.
Постоянная смешанная микротрубочковая и актин-миозиновая система обнаружена в субмембранной области таких крайне специализированных образований, как кровяные пластинки млекопитающих, представляющие собой свободно циркулирующие в крови участки цитоплазмы полиплоидных клеток мегакариоцитов.
Помимо хорошо развитой в периферической гиалоплазме актин-миозиновой фибриллярной системы здесь имеется мощное кольцо микротрубочек, по-видимому, обеспечивающих поддержание формы этих структур.
Актин-миозиновая система кровяных пластинок играет важную роль в процессе свертывания крови.
Смешанные постоянные актин-миозиновая и тубулин-динеиновая системы, очевидно, широко распространены у высших простейших и, в частности, у инфузорий.
Однако в настоящее время они изучены преимущественно на уровне чисто морфо логического, ультраструктурного анализа. Функциональное взаимодействие названных двух основных механохимически: систем интенсивно исследуется у метазойных клеток в процессах митотического деления. Этот вопрос мы подробнее рассмотрим ниже, при описании процессов репродукции клеток.
Микротрубочки представляют собой микроскопические полые трубки, изготовленные из белков альфа и бета тубулина, которые являются частью клетка «s цитоскелет сеть белковых нитей, которая распространяется по всей клетке, придает клетке форму и сохраняет ее органеллы на месте. Микротрубочки являются самыми крупными структурами в цитоскелете толщиной около 24 нанометров. У них есть роли в движении клеток, деление клеток и транспортировка материалов внутри камер.
Структура микротрубочек
Микротрубочки представляют собой полые цилиндры, состоящие из повторяющихся белковых структур, в частности димеров альфа- и бета-тубулинов (также обозначаемых в письменном виде как ɑ-тубулин и β-тубулин). Димеры представляют собой комплексы двух белков. tub-тубулин и β-тубулин связываются друг с другом, образуя димер, и затем несколько единиц этих димеров связываются вместе, всегда чередуя альфа и бета, образуя цепь, называемую протофиламентом. Затем тринадцать протофиламентов выстраиваются в цилиндрический рисунок, образуя микротрубочки. Микротрубочки постоянно собираются и разбираются путем добавления и удаления димеров. Говорят, что они находятся в состоянии динамическое равновесие потому что их структура сохраняется, хотя сами молекулы постоянно меняются.
Микротрубочки представляют собой полярные молекулы с положительно заряженным концом, который растет относительно быстро, и отрицательно заряженным концом, который растет относительно медленно. Протофиламенты располагаются параллельно друг другу в микротрубочке, поэтому на положительном конце микротрубочки всегда присутствуют бета-субъединицы, а на отрицательном конце – альфа-субъединицы. Наличие полярности позволяет микротрубочке собираться определенным образом и функционировать правильно.
В клетках животных микротрубочки выходят из органеллы в центре клетки называется центросома, который является центром организации микротрубочек (MTOC). Клетки растений и грибы не имеют центросом, и вместо этого ядерная оболочка – мембрана, окружающая ядро клетки – представляет собой MTOC.

Функция микротрубочек
Движение клетки
Клеточный отдел
Микротрубочки играют ключевую роль в формировании митотического веретена, также называемого веретенообразным аппаратом. Это структура, которая формируется во время митоз (деление клеток) в эукариотических клетках. Митотический веретено организует и разделяет хромосомы во время клеточного деления, так что хромосомы могут быть разделены на две отдельные дочерние клетки, Его компоненты включают микротрубочки, MTOC и микротрубочковые белки (MAP).
Три подгруппы микротрубочек помогают в процессе митоза: астральные, полярные и кинетохорные микротрубочки. Астральные микротрубочки излучают от MTOCs клетки к клеточная мембрана, удерживая митотический веретено на месте. Полярные микротрубочки переплетаются между двумя MTOCs и помогают разделить хромосомы. (Все микротрубочки являются полярными; их просто специально называют полярными микротрубочками.) Микротрубочки Kinetochore прикрепляются к хромосомам, чтобы помочь их разделить; хромосомы прикреплены к микротрубочкам с помощью комплекса белков, называемых кинетохорами.
Клеточный транспорт
Как часть цитоскелета, микротрубочки помогают перемещать органеллы внутри клетки. цитоплазма, который является всем содержимым клетки, за исключением ее ядра. Они также помогают различным областям клетки общаться друг с другом. Однако, хотя микротрубочки помогают компонентам клетки двигаться, они также придают клетке форму и структуру.
Другие цитоскелетные компоненты
Другими двумя основными компонентами эукариотического цитоскелета являются микрофиламенты и промежуточные филаменты. Микрофиламенты меньше микротрубочек диаметром около 7 нм. Они помогают в делении цитоплазмы во время деления клеток, а также играют роль в цитоплазматическом потоке, который является потоком цитозоль (клеточная жидкость) по всей клетке. Промежуточные нити больше, чем микрофиламенты, но меньше, чем микротрубочки. Они помогают придать клетке форму и обеспечивают структурную поддержку.
- Tubulin – Белок, из которого состоят микротрубочки.
- цитоплазма – все содержимое клетки, кроме ядра.
- Шпиндельный аппарат – Структура, образованная в основном микротрубочками, которая разделяет хромосомы во время митоза.
- кинетохорной – Белковый комплекс, который помогает прикрепить хромосомы к микротрубочкам в митотическом веретене.
викторина
1. Центросомы находятся в каком типе клеток?A. грибковыйB. животноеC. Растение D. бактериальный
Ответ на вопрос № 1
В верно. Центросомы обнаружены только в клетках животных. У растений и грибов микротрубочки организованы ядерной оболочкой.
2. Микротрубочки являются ______ компонентами цитоскелета.A. Самый толстыйB. ТончайшийC. самый длинныйD. самый короткий
Ответ на вопрос № 2
верно. Микротрубочки – самые толстые компоненты цитоскелета на длине около 24 нм. Промежуточные нити меньше, чем микротрубочки, а микрофиламенты меньше, чем промежуточные нити.
3. Что НЕ является функцией микротрубочек?A. Чтобы помочь разделить хромосомы во время деления клеткиB. Для формирования ресничек и жгутиковC. Чтобы помочь клетке перемещаться из одного места в другоеD. Помочь в делении цитоплазмы во время митоза
Ответ на вопрос № 3
D верно. Микротрубочки не разделяют цитоплазму во время митоза, но они помогают разделить хромосомы, чтобы в каждую дочернюю клетку могло попасть одинаковое количество хромосом.
Различают по меньшей мере три типа таких структур: микротрубочки, микрофиламенты и промежуточные филаменты. Их функции связаны с внутриклеточным движением, со способностью клеток поддерживать свою форму, а также с некоторыми другими видами активности клеток, такими, например, как эндоцитоз и экзоцитоз. Мы рассмотрим здесь только микротрубочки.
Микротрубочки содержатся почти во всех эукариотических клетках. Это полые, очень тонкие неразветвленные трубочки диаметром приблизительно 24 нм; их стенки толщиной около 5 нм построены из спирально упакованных субъединиц белка тубулина.
Рисунок дает представление о том, как выглядят микротрубочки на электронных микрофотографиях. Растут микротрубочки с одного конца путем добавления тубулиновых субъединиц. Рост видимо, может начаться лишь при наличии матрицы; есть основания полагать, что роль таких матриц играют какие-то очень мелкие кольцевые структуры, которые были выделены из клеток и которые, как выяснилось, состоят из тубулиновых субъединиц. В интактных клетках ту же функцию выполняют центриоли, поэтому их иногда называют центрами организации микротрубочек (ЦОМ). Центриоли состоят из коротких микротрубочек.
Микротрубочки принимают участие в различных внутриклеточных процессах; некоторые мы здесь упомянем.
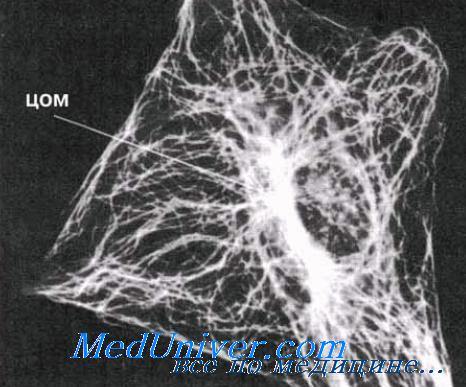
Распределение микротрубочек в клетке. Микротрубочки расходятся от центра организации микротрубочек (ЦОМ), находящегося рядом с ядром. В ЦОМ содержится центриоль. Микротрубочки видны на этой микрофотографии благодаря использованию флуоресцирующих антител, способных специфически соединяться с их белком. Представленная здесь клетка — фибробласт; фибробласты обычно содержатся в соединительной ткани; в них синтезируется коллаген.
Центриоли и деление ядра
Микротрубочки регулируют расхождение хроматид или хромосом. Осуществляется это за счет скольжения микротрубочек. В клетках высших растений центриоли отсутствуют, хотя веретено в них при делении ядра образуется. Возможно, что в этих клетках имеются какие-то очень мелкие центры организации микротрубочек, не выявляемые даже при помощи электронного микроскопа.
Базальные тельца, реснички и жгутики
В ресничках и жгутиках движение осуществляется за счет скольжения микротрубочек. Более подробно эти процессы описаны в нашей статье. Отметим, что жгутики бактерий устроены проще, чем жгутики эукариот, и базальные тельца у них отсутствуют.
Внутриклеточный транспорт
Микротрубочки участвуют также в перемещении различных клеточных органелл, например в перемещении пузырьков Гольджи к формирующейся клеточной пластинке (рис. 5.30). В клетке идет непрерывный транспорт: перемещаются пузырьки Гольджи, направляются к аппарату Гольджи пузырьки, отпочковывающиеся от ЭР, движутся лизосомы, митохондрии и другие органеллы. Все это движение приостанавливается, если повреждена система микротрубочек.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Читайте также: