
Мембранная теория бернштейна кратко
Обновлено: 07.07.2024
Потенциа́л поко́я (ПП) - потенциал возбудимой клетки в невозбужденном состоянии. Он представляет собой разность электрических потенциалов, имеющихся на внутренней и наружной сторонах мембраны и составляет у теплокровных от -55 до -100 мВ [1] . У нейронов и нервных волокон обычно составляет -70 мВ. Измеряется изнутри клетки.
Содержание
История открытия
В 1902 г. Ю. Бернштейн выдвинул гипотезу, согласно которой клеточная мембрана пропускает внутрь клетки ионы К + , и они накапливаются в цитоплазме. Расчет величины потенциала покоя по уравнению Нернста для калиевого электрода удовлетворительно совпал с измеренным потенциалом между саркоплазмой мышцы и окружающей средой, который составлял около — 70 мВ.
Согласно теории Ю. Бернштейна, при возбуждении клетки её мембрана повреждается, и ионы К + вытекают из клетки по концентрационному градиенту до тех пор, пока потенциал мембраны не становится равным нулю. Затем мембрана восстанавливает свою целостность, и потенциал возвращается к уровню потенциала покоя. Это утверждение, относящееся скорее к потенциалу действия, было опровергнуто Ходжкином и Хаксли в 1939 году.
Общие положения
Для того, чтобы на мембране поддерживалась разность потенциалов, необходимо, чтобы была определенная разность концентрации различных ионов внутри и снаружи клетки.
Концентрации ионов в клетке скелетной мышцы и во внеклеточной среде
| Ионы | Концентрация в саркоплазме (ммоль) | Концентрация вне клетки (ммоль) | |
|---|---|---|---|
| K + | 140 | 2,5 | |
| Na + | 10 | 120 | |
| Cl - | 3-4 | 120 | |
| Ca 2+ | - (полипептиды) | 140 | 0 |

Различия в концентрации ионов на противоположных сторонах клеточной мембраны приводят к напряжению, названному мембранным потенциалом. Много ионов имеют градиент концентрации поперек мембраны, включая калий (K+), который является в высокой внутренней части и низкой концентрации вне мембраны. Натрий (Na+) и хлорид (Замкнутый–) ионы - при высоких концентрациях во внеклеточной области, и низких концентрациях во внутриклеточных областях. Эти градиенты концентрации обеспечивают потенциальную энергию вести формирование мембранного потенциала. Это напряжение установлено, когда мембрана имеет проходимость к одному или более ионам. В самом простом случае, иллюстрированном здесь, если мембрана выборочно водопроницаема к калию, они положительно обвиняли, что ионы могут распространить вниз градиент концентрации к внешней стороне ячейки, оставляя позади неданные компенсацию отрицательные обвинения. Это разделение обвинений-, каков вызывает мембранный потенциал. Отметьте, что оптовые решения любой стороны мембраны electo-нейтральны. Аналогично, система в целом нейтральна гальваностереотипом. "Неданные компенсацию" положительные обвинения вне ячейки, и неданных компенсацию отрицательных обвинений в ячейке, физически выстраиваются в линию на мембранной поверхности и привлекают друг друга поперек мембраны. Таким образом, мембранный потенциал физически расположен только в непосредственной близости мембраны. Это - разделение этих обвинений поперек них мембрана, которая является основанием мембранного напряжения. Отметьте также, что эта диаграмма - только приближение ионных вкладов в мембранный потенциал. Другие ионы, включая натрий, хлорид, кальций и другие играют более незначительную роль, даже при том, что они имеют сильные градиенты концентрации, потому что они более ограничили проходимость чем калий. Ключ: Синие пятиугольники - ионы натрия; Фиолетовые площади - ионы калия; Желтые круги - ионы Choloride; Оранжевые прямоугольники - Анионы (они являются результатом разнообразия источников, включая белки). Большая фиолетовая структура со стрелкой представляет трансмембранный канал калия и руководство чистого движения калия.
С помощью хлора, вернее, их равновесные потенциалы.
Впоследствии было доказано, что основной вклад в создание потенциала покоя вносит выходящий калиевый ток, который осуществляется через специфические белки-каналы — калиевые каналы постоянного тока. В покое калиевые каналы открыты, а натриевые каналы закрыты. Ионы калия выходят из клетки по градиенту концентрации, что создает на наружной стороне мембраны избыток положительных зарядов; при этом на внутренней стороне мембраны остаются отрицательные заряды. Некоторый (небольшой) вклад в создание потенциала покоя вносит также работа так называемого "натрий-калиевого насоса", который образован особым мембранным ферментом - натрий-калиевой АТФазой .
Потенциал покоя для большинства нейронов составляет величину порядка −60 мВ — −70 мВ. У клеток невозбудимых тканей на мембране также имеется разность потенциалов, разная для клеток разных тканей и организмов.
Формирование потенциала покоя
ПП формируется в два этапа.
Первый этап: создание незначительной (-10 мВ) отрицательности внутри клетки за счёт неравного асимметричного обмена Na + на K + в соотношении 3 : 2. В результате этого клетку покидает больше положительных зарядов с натрием, чем возвращается в неё с калием. Такая особенность работы натрий-калиевого насоса, осуществляющего взаимообмен этих ионов через мембрану с затратами энергии См. также
Примечания
Ссылки
Дудель Й., Рюэгг Й., Шмидт Р. и др. Физиология человека: в 3-х томах. Пер. с англ / под ред Р. Шмидта и Г. Тевса. — 3. — М .: Мир, 2007. — Т. 1. — 323 с илл. с. — 1500 экз. — ISBN 5-03-000575-3. (см. ISBN )
Современная теория электрогенеза вытекает исторически из положений Бернштейна (1940), связавшему впервые ионную проницаемость мембран с природой биопотенциалов.
Ее основные положения:
1. В состоянии покоя мембрана проницаема только для ионов калия.
2. Ионы калия, выходя из клетки, создают потенциал, величина которого рассчитывается по уравнению Нернста:
где [К + ]i и [К + ]e – концентрации внутриклеточных – i и внеклеточных ионов – e, соответственно.
3. Этот потенциал равновесный, так как различие концентраций [К+]i и [К+]e присутствует в клетках постоянно.
В качестве доказательства своей теории Бернштейн указывал на зависимость мембранного потенциала от температуры, изменений внеклеточной концентрации ионов калия [К + ]e, т.е. от расчетных показателей уравнения Нернста.
Теория постоянного поля и потенциал покоя (ПП)
Современные представления об электрогенезе основаны на положениях теории постоянного поля. Ее основные создатели: Ходжкин, Хаксли и Катц (1949) разработали ряд упрощений для возможного анализа протекания электрических процессов на мембране:
1. Ионы движутся в мембране по градиенту концентрации и электрического поля, как в растворе.
2. Напряженность электрического поля в мембране постоянна и потенциал падает линейно ее толщины по мере удаления от источника электрического поля.
3. Мембрана гомогенна, имея постоянную диэлектрическую проницаемость по всей толщине.
4. Концентрация ионов в мембране на границе с раствором пропорциональна концентрации ионов в самом растворе.
Из первого выражения вытекает, что плотность ионного тока через мембрану:
zi –валентность i-го иона, V- потенциал в точке, x – расстояние по толщине мембраны от наружней ее границы. Т.е.плотность ионного тока определяется разностью между диффузионным потоком i-го иона по концентрационному градиенту (1-ое слагаемое) и противоположно направленным потоком этого иона, обусловленным электрическим полем, возникшем при его диффузии (2-ое слагаемое).
При постоянстве электрического поля dV/dx =const=V/a, где а – тощина мебраны константа проницаемости мембраны (Pi) определяется относительной подвижностью и растворимостью в мембране:
βi – коэффициент распределения i-го иона между водным раствором и мембраной, ui –подвижность этого иона в электрическом поле.
На основании представленных положений теории постоянного поля Ходжкин, Гольдман и Катц вывели уравнение зависимости плотности ионных токов от мембранного потенциала и ионной проницаемости:
Предполагая, что при потенциале покоя (ПП) суммарный ионный ток равен нулю Ходжкин, Гольдман и Катц вывели уравнение:
[К + ]i, [Na + ]i, [Сl - ]i и [К + ]e [Na + ]e и [Cl - ]e – концентрации внутриклеточных – intra и внеклеточных ионов – extra соответственно,
PK, PNa и PCl – проницаемость соответствующих ионов.
Для ПП гигантского аксона кальмара соотношение:
Таким образом, ПП имеет в основном калиевую природу, что и определяет его вклад в изменения мембранного потенциала при модуляции концентрации ионов калия. Распределение ионов между клеткой и средой при ПП устанавливается на постоянном уровне, и суммарный ионный ток равняется нулю:
Постоянно существующий градиент, поддерживающий пассивный поток ионов через клеточную мембрану, требует процессов, его восстанавливающих. Их выполняет система активного транспорта – Mg 2+ -зависимая Na + /K + -АТФ-аза, которая осуществляет противоградиентный транспорт ионов Na + и K + . Энергозависимый процесс происходит в режиме одновременного выброса 3-х ионов Na + из клетки и закачивания 2-х ионов K + внутрь. Создание дополнительного дефицита внутриклеточного содержимого в положительном заряде позволило режим работы данной системы, кроме энергозависимого, считать еще и электрогенным.
Потенциал действия (ПД)
Раздражение, как уже упоминалось, лишь при достижении порога возбуждения вызывает самоподдерживающийся процесс изменения значений биопотенциалов в клетке – потенциал действия. Пороговые условия начала развития ПД определяются многочисленными условиями (законами), подробно рассмотренными в соответствующих электрофизиологических литературных разделах. Например, они включают в себя силу (амплитуду) и длительность раздражающего стимула. Взаимосвязь этих параметров представляет собой известную гиперболическую зависимость "сила – длительность" (Рис.17).

 | Рис.17. Зависимость сила – длительность. По оси абсцисс- длительность стимула; по оси ординат -амплитуда стимула С - полезное время; А -хроноксия; О - реобаза |
Такие характеристики, как полезное время, реобаза, хронаксия являются общими для электровозбудимых тканей (там же).
Электротонический потенциал и локальный ответ. Изменения, проходящие на мембране при действии раздражения до порогового уровня, проявляются в виде электротонического потенциала и локального ответа.
Электротонический потенциал представляет собой изменения пассивных (омических и емкостных) электрических характеристик мембран в ответ на раздражение, амплитуда которого меньше 50 – 70% пороговых (100%) значений.
Локальный ответ представляет собой изменения подпороговых активных (ионная проницаемость) электрических характеристик мембран в ответ на раздражение, амплитуда которого ниже пороговых значений.
Таким образом, основной причиной инициации процесса развития ПД является достижение критического (порогового) уровня смещения мембранного потенциала.
Современная теория электрогенеза вытекает исторически из положений Бернштейна (1940), связавшему впервые ионную проницаемость мембран с природой биопотенциалов.
Ее основные положения:
1. В состоянии покоя мембрана проницаема только для ионов калия.
2. Ионы калия, выходя из клетки, создают потенциал, величина которого рассчитывается по уравнению Нернста:
где [К + ]i и [К + ]e – концентрации внутриклеточных – i и внеклеточных ионов – e, соответственно.
3. Этот потенциал равновесный, так как различие концентраций [К+]i и [К+]e присутствует в клетках постоянно.
В качестве доказательства своей теории Бернштейн указывал на зависимость мембранного потенциала от температуры, изменений внеклеточной концентрации ионов калия [К + ]e, т.е. от расчетных показателей уравнения Нернста.
Теория постоянного поля и потенциал покоя (ПП)
Современные представления об электрогенезе основаны на положениях теории постоянного поля. Ее основные создатели: Ходжкин, Хаксли и Катц (1949) разработали ряд упрощений для возможного анализа протекания электрических процессов на мембране:
1. Ионы движутся в мембране по градиенту концентрации и электрического поля, как в растворе.
2. Напряженность электрического поля в мембране постоянна и потенциал падает линейно ее толщины по мере удаления от источника электрического поля.
3. Мембрана гомогенна, имея постоянную диэлектрическую проницаемость по всей толщине.
4. Концентрация ионов в мембране на границе с раствором пропорциональна концентрации ионов в самом растворе.
Из первого выражения вытекает, что плотность ионного тока через мембрану:
zi –валентность i-го иона, V- потенциал в точке, x – расстояние по толщине мембраны от наружней ее границы. Т.е.плотность ионного тока определяется разностью между диффузионным потоком i-го иона по концентрационному градиенту (1-ое слагаемое) и противоположно направленным потоком этого иона, обусловленным электрическим полем, возникшем при его диффузии (2-ое слагаемое).
При постоянстве электрического поля dV/dx =const=V/a, где а – тощина мебраны константа проницаемости мембраны (Pi) определяется относительной подвижностью и растворимостью в мембране:
βi – коэффициент распределения i-го иона между водным раствором и мембраной, ui –подвижность этого иона в электрическом поле.
На основании представленных положений теории постоянного поля Ходжкин, Гольдман и Катц вывели уравнение зависимости плотности ионных токов от мембранного потенциала и ионной проницаемости:
Предполагая, что при потенциале покоя (ПП) суммарный ионный ток равен нулю Ходжкин, Гольдман и Катц вывели уравнение:
[К + ]i, [Na + ]i, [Сl - ]i и [К + ]e [Na + ]e и [Cl - ]e – концентрации внутриклеточных – intra и внеклеточных ионов – extra соответственно,
PK, PNa и PCl – проницаемость соответствующих ионов.
Для ПП гигантского аксона кальмара соотношение:
Таким образом, ПП имеет в основном калиевую природу, что и определяет его вклад в изменения мембранного потенциала при модуляции концентрации ионов калия. Распределение ионов между клеткой и средой при ПП устанавливается на постоянном уровне, и суммарный ионный ток равняется нулю:
Постоянно существующий градиент, поддерживающий пассивный поток ионов через клеточную мембрану, требует процессов, его восстанавливающих. Их выполняет система активного транспорта – Mg 2+ -зависимая Na + /K + -АТФ-аза, которая осуществляет противоградиентный транспорт ионов Na + и K + . Энергозависимый процесс происходит в режиме одновременного выброса 3-х ионов Na + из клетки и закачивания 2-х ионов K + внутрь. Создание дополнительного дефицита внутриклеточного содержимого в положительном заряде позволило режим работы данной системы, кроме энергозависимого, считать еще и электрогенным.
Потенциал действия (ПД)
Раздражение, как уже упоминалось, лишь при достижении порога возбуждения вызывает самоподдерживающийся процесс изменения значений биопотенциалов в клетке – потенциал действия. Пороговые условия начала развития ПД определяются многочисленными условиями (законами), подробно рассмотренными в соответствующих электрофизиологических литературных разделах. Например, они включают в себя силу (амплитуду) и длительность раздражающего стимула. Взаимосвязь этих параметров представляет собой известную гиперболическую зависимость "сила – длительность" (Рис.17).
| | Рис.17. Зависимость сила – длительность. По оси абсцисс- длительность стимула; по оси ординат -амплитуда стимула С - полезное время; А -хроноксия; О - реобаза |
Такие характеристики, как полезное время, реобаза, хронаксия являются общими для электровозбудимых тканей (там же).
Электротонический потенциал и локальный ответ. Изменения, проходящие на мембране при действии раздражения до порогового уровня, проявляются в виде электротонического потенциала и локального ответа.
Электротонический потенциал представляет собой изменения пассивных (омических и емкостных) электрических характеристик мембран в ответ на раздражение, амплитуда которого меньше 50 – 70% пороговых (100%) значений.
Локальный ответ представляет собой изменения подпороговых активных (ионная проницаемость) электрических характеристик мембран в ответ на раздражение, амплитуда которого ниже пороговых значений.
Таким образом, основной причиной инициации процесса развития ПД является достижение критического (порогового) уровня смещения мембранного потенциала.
Введение
Зачем нам нужно знать, что такое потенциал покоя?
Что такое "животное электричество"? Откуда в организме берутся "биотоки"? Как живая клетка, находящаяся в водной среде, может превратиться в "электрическую батарейку"?
- На эти вопросы мы сможем ответить, если узнаем, как клетка за счёт перераспределения электрических зарядов (заряженных частиц - ионов) создаёт себе электрический потенциал на мембране.
Как работает нервная система? С чего в ней всё начинается? Откуда в ней берётся электричество для нервных импульсов?
- На эти вопросы мы также сможем ответить, если узнаем, как нервная клетка создаёт себе электрический потенциал на мембране.
Итак, понимание того, как работает нервная система, начинается с того, что надо разобраться, как работает отдельная нервная клетка - нейрон.
В основе работы нейрона с нервными импульсами лежит перераспределение электрических зарядов на его мембране и изменение величины электрических потенциалов. Но чтобы потенциал изменять, его нужно для начала иметь. Поэтому можно сказать, что нейрон, готовясь к cвоей нервной работе, создаёт на своей мембране электрический потенциал, как возможность для такой работы.
Таким образом, наш самый первый шаг к изучению работы нервной системы - это понять, каким образом перемещаются электрические заряды в нервных клетках и как за счёт этого на мембране появляется электрический потенцил. Этим мы и займёмся, и назовём этот процесс появления электрического потенциала у нейронов - формирование потенциала покоя.
Определение
В норме, когда клетка готова к работе, у неё уже есть электрический потенциал на поверхности мембраны. Он называется мембранный потенциал покоя.
Потенциал покоя - это разность электрических потенциалов между внутренней и наружной сторонами мембраны, когда клетка находится в состоянии физиологического покоя. Его средняя величина составляет -70 мВ (милливольт).
"Потенциал" - это возможность, он сродни понятию "потенция". Электрический потенциал мембраны - это её возможности по перемещению электрических зарядов, положительных или отрицательных. В роли зарядов выступают заряженные химические частицы - ионы натрия и калия, а также кальция и хлора. Из них только ионы хлора заряжены отрицательно (-), а остальные - положительно (+).
Таблица 1. Физико-химические свойства ионов натрия и калия
Координационное число
Равновесный потенциал, мВ
Предельная температура гидратации, ºС
Однако, и Na + , и К + потенциально способны перемещаться под действием электрических сил туда, где преобладают не положительные, а отрицательные электрические заряды (отрицательно заряженные частицы - анионы).
Итак, имея электрический потенциал, мембрана может перемещать в клетку или из клетки указанные выше заряженные ионы с помощью электрических сил.
Важно понимать, что в нервной системе электрические заряды создаются не электронами, как в металлических проводах, а ионами - химическими частицами, имеющими электрический заряд. Электрический ток в организме и его клетках - это поток ионов, а не электронов, как в проводах. Обратите также внимание на то, что заряд мембраны измеряется изнутри клетки, а не снаружи.
Если говорить уж совсем примитивно просто, то получается, что снаружи вокруг клетки будут преобладать "плюсики", т.е. положительно заряженные ионы, а внутри - "минусики", т.е. отрицательно заряженные ионы. Можно сказать, что внутри клетка электроотрицательна. И теперь нам всего лишь надо объяснить, как это так получилось. Хотя, конечно, неприятно сознавать, что все наши клетки - отрицательные "персонажи". ((
Сущность
Сущность потенциала покоя - это преобладание на внутренней стороне мембраны отрицательных электрических зарядов в виде анионов и недостаток положительных электрических зарядов в виде катионов, которые сосредотачиваются на её наружной стороне, а не на внутренней.
Внутри клетки - "отрицательность", а снаружи - "положительность".
Такое положение вещей достигается с помощью трёх явлений: (1) поведения мембраны, (2) поведения положительных ионов калия и натрия и (3) соотношения химической и электрической силы.
1. Поведение мембраны
В поведении мембраны для потенциала покоя важны три процесса:
1) Обмен внутренних ионов натрия на наружные ионы калия. Обменом занимаются специальные транспортные структуры мембраны: ионные насосы-обменники. Таким способом мембрана перенасыщает клетку калием, но обедняет натрием.
2) Открытые калиевые ионные каналы. Через них калий может как заходить в клетку, так и выходить из неё. Он выходит в основном.
3) Закрытые натриевые ионные каналы. Из-за этого натрий, выведенный из клетки насосми-обменниками, не может вернуться в неё обратно. Натриевые каналы открываются только при особых условиях - и тогда потенциал покоя нарушается и смещается в сторону нуля (это называется деполяризацией мембраны, т.е. уменьшением полярности).
2. Поведение ионов калия и натрия
Ионы калия и натрия по-разному перемещаются через мембрану:
2) Через постоянно открытые калиевые каналы калий выходит из клетки, но может и возвращаться в неё обратно через них же.
3) Натрий "хочет" войти в клетку, но "не может", т.к. каналы для него закрыты.
3. Соотношение химической и электрической силы
По отношению к ионам калия между химической и электрической силой устанавливается равновесие на уровне - 70 мВ.
1) Химическая сила выталкивает калий из клетки, но стремится затянуть в неё натрий.
2) Электрическая сила стремится затянуть в клетку любые положительно заряженные ионы (как натрий, так и калий).
Формирование потенциала покоя
Попробую рассказать коротко, откуда берётся мембранный потенциал покоя в нервных клетках - нейронах. Ведь, как всем теперь известно, наши клетки только снаружи положительные, а внутри они весьма отрицательные, и в них существует избыток отрицательных частиц - анионов и недостаток положительных частиц - катионов.
И вот тут исследователя и студента поджидает одна из логических ловушек: внутренняя электроотрицательность клетки возникает не из-за появления лишних отрицательных частиц (анионов), а наоборот - из-за потери некоторого количества положительных частиц (катионов).
И поэтому сущность нашего рассказа будет заключаться не в том, что мы объясним, откуда берутся отрицательные частицы в клетке, а в том, что мы объясним, каким образом в нейронах получается дефицит положительно заряженных ионов - катионов.
Куда же деваются из клетки положительно заряженные частицы? Напомню, что это ионы натрия - Na + и калия - K + . Удивительно, но оба этих иона покидают клетку, хотя и разными путями.
С чего начать, чтобы не сбиться и не запутаться? Начнём с калия.
Чтобы легче было запомнить, образно можно сказать так:
"Клетка любит калий!"
или: "клетка копит калий!" (К + )
и "ненавидит натрий" (Na + ).
И что же она, клетка, делает с этими ионами? Она их обменивает друг на друга! С помощью специального обменного насоса.
Натрий-калиевый насос
Дело заключается в том, что в мембране нервной клетки постоянно работают насосы-обменники, образованные специальными белками, встроенными в мембрану. Что они делают? Они меняют "собственный" натрий клетки на наружный "чужой" калий. Из-за этого в клетке оказывается в конце концов недостаток натрия, который ушёл на обмен. И в то же время клетка переполняется ионами калия, который в неё натащили эти молекулярные насосы.
Итак: "Клетка любит калий!" (Хотя об истинной любви здесь, разумеется, не может идти и речи!) Поэтому она и затаскивает калий в себя, несмотря на то, что его итак уже полно. Поэтому она невыгодно обменивает его на натрий, отдавая 3 иона натрия за 2 иона калия (любимого). Поэтому она тратит на этот обмен энергию АТФ. И как тратит! До 70% всех энергозатрат нейрона может уходить на работу натрий-калиевых насосов. Вот что делает любовь, пусть даже не настоящая!
Кстати, интересно, что клетка не рождается с потенциалом покоя в готовом виде. Например, при дифференцировке и слиянии миобластов при формировании мышечного волокна потенциал их мембраны изменяется от -10 до -70 mV, т.е. их мембрана становится более электроотрицательной, она поляризуется в процессе дифференцировки. А в экспериментах на мультипотентных мезенхимальных стромальных клетках (ММСК) костного мозга человека искусственная деполяризация ингибировала дифференцировку клеток (Fischer-Lougheed J., Liu J.H., Espinos E. et al. Human myoblast fusion requires expression of functional inward rectifier Kir2.1 channels. Journal of Cell Biology 2001; 153: 677-85; Liu J.H., Bijlenga P., Fischer-Lougheed J. et al. Role of an inward rectifier K+ current and of hyperpolarization in human myoblast fusion. Journal of Physiology 1998; 510: 467-76; Sundelacruz S., Levin M., Kaplan D.L. Membrane potential controls adipogenic and osteogenic differentiation of mesenchymal stem cells. Plos One 2008; 3 ).
Образно говоря, можно выразиться так:
Создавая потенциал покоя, клетка "заряжается любовью".
Это любовь к двум вещам:
1) любовь клетки к калию,
2) любовь калия к свободе.
Как ни странно, но результат этих двух видов любви - пустота!
Именно она, пустота, создаёт в клетке отрицательный электрический потенциал - потенциал покоя. Точнее, отрицательный потенциал создают пустые места, оставшиеся от убежавшего из клетки калия.
Итак, результат деятельности мембранных ионных насосов-обменников таков:
Натрий-калиевый ионный насос-обменник создаёт три потенциала (возможности):
1. Электрический потенциал - возможность притягивать внутрь клетки любые положительно заряженные частицы (ионы).
2. Ионный натриевый химический потенциал - возможность затягивать внутрь клетки ионы натрия (и именно натрия, а не какого-нибудь другого вещества).
3. Ионный калиевый химический потенциал - возможновть выталкивать из клетки ионы калия (и именно калия, а не какого-нибудь другого вещества).
4. Дефицит натрия (Na + ) в клетке.
5. Избыток калия (K + ) в клетке.
Можно сказать так: ионные насосы мембраны создают разность концентраций ионов, или градиент (перепад) концентрации, между внутриклеточной и внеклеточной средой.
Именно из-за получившегося дефицита натрия в клетку теперь "полезет" этот самый натрий снаружи. Так всегда ведут себя вещества: они стремятся выровнять свою концентрацию во всём объёме раствора.
И в то же время в клетке получился избыток ионов калия по сравнению с наружной средой. Потому что насосы мембраны накачали его в клетку. И он стремится уравнять свою концентрацию внутри и снаружи, и поэтому стремится выйти из клетки.
Тут ещё важно понять, что ионы натрия и калия как бы "не замечают" друг друга, они реагируют только "на самих себя". Т.е. натрий реагирует на концентрацию натрия же, но "не обращает внимания" на то, сколько вокруг калия. И наоборот, калий реагирует только на концентрацию калия и "не замечает" натрий. Получается, что для понимания поведения ионов в клетке надо по-отдельности сравнивать концентрации ионов натрия и калия. Т.е. надо отдельно сравнить концентрацию по натрию внутри и снаружи клетки и отдельно - концентрацию калия внутри и снаружи клетки, но не имеет смысла сравнивать натрий с калием, как это часто делается в учебниках.
По закону выравнивания концентраций, который действует в растворах, натрий "хочет" снаружи войти в клетку. Но не может, так как мембрана в обычном состоянии плохо его пропускает. Его заходит немножко, и клетка его опять тут же обменивает на наружный калий. Поэтому натрий в нейронах всегда в дефиците.
А вот калий как раз может легко выходить из клетки наружу! В клетке его полно, и она его удержать не может. Так вот он и выходит наружу через особые белковые дырочки в мембране (ионные каналы).
Анализ
От химического - к электрическому
А теперь - самое главное, следите за излагаемой мыслью! Мы должны перейти от движения химических частиц к движению электрических зарядов.
Калий заряжен положительным зарядом, и поэтому он, когда выходит из клетки, выносит из неё не только себя, но и "плюсики" (положительные заряды). На их месте в клетке остаются "минусы" (отрицательные заряды). Это и есть мембранный потенциал покоя!
Мембранный потенциал покоя - это дефицит положительных зарядов (положительно заряженных ионов) внутри клетки, образовавшийся за счёт утечки из клетки положительных ионов калия.
Заключение

Рис. Схема формирования потенциала покоя (ПП). Автор благодарит Попову Екатерину Юрьевну за помощь в создании рисунка.
Составные части потенциала покоя
Потенциал покоя - отрицательный со стороны клетки и состоит как бы из двух частей.
1. Первая часть - это примерно -10 милливольт, которые получаются от неравносторонней работы мембранного насоса-обменника (ведь он больше выкачивает "плюсиков" с натрием, чем закачивает обратно с калием).
2. Вторая часть - это утекающий всё время из клетки калий, утаскивающий положительные заряды из клетки. Он дает большую часть мембранного потенциала, доводя его до -70 милливольт.
Калий перестанет выходить из клетки (точнее, его вход и выход сравняются) только при уровне электроотрицательности клетки в -90 милливольт. Но этому мешает постоянно подтекающий в клетку натрий, который тащит с собой свои положительные заряды. И в клетке поддерживается равновесное состояние на уровне -70 милливольт.
Обратите внимание на то, что для создания потенциала покоя нужны затраты энергии. Эти затраты производятся ионными насосами, которые обменивают "свой" внутренний натрий (ионы Na + ) на "чужой" внешний калий (K + ). Вспомним, что ионные насосы являются ферментами АТФазами и расщепляют АТФ, получая из неё энергию на указанный обмен ионов разного типа друг на друга.Тут очень важно понять, что с мембраной "работают" сразу 2 потенциала: химический (концентрационный градиент ионов) и электрический (разность электрических потенциалов по разные стороны мембраны). Ионы перемещаются в ту или иную сторону под действием обеих этих сил, на которые и тратится энергия. При этом один из двух потенциалов (химический или электрический) уменьшается, а другой увеличивается. Разумеется, если рассматривать электрический потенциал (разность потенциалов) отдельно, то не будут учитываться "химические" силы, перемещающие ионы. И тогда может сложиться неверное впечатление о том, что энергия на движение ионо берётся как бы ниоткуда. Но это не так. Необходимо рассматривать обе силы: химическую и электрическую. При этом крупные молекулы с отрицательными зарядами, находящиеся внутри клетки играют роль "статистов", т.к. их не перемещают через мембрану ни химические, ни электрические силы. Поэтому эти отрицательные частицы обычно и не рассматривают, хотя они существуют и именно они обеспечивают отрицательную сторону разности потенциалов между внутренней и наружной сторонами мембраны. А вот шустрые ионы калия, как раз способны к перемещению, и именно их утечка из клетки под действием химических сил создаёт львиную долю электрического потенциала (разности потенциалов). Ведь именно ионы калия перемещают на наружную сторону мембраны положительные электрические заряды, будучи положительно заряженными частицами.
Так что всё дело в натрий-калиевом мембранном насосе-обменнике и последующем вытекании из клетки "лишнего" калия. За счёт потери положительных зарядов при этом вытекании внутри клетки нарастает электроотрицательность. Она-то и есть "мембранный потенциал покоя". Он измеряется внутри клетки и составляет обычно -70 мВ.
Выводы
Говоря образно, "мембрана превращает клетку в "электрическую батарейку" с помощью управления ионными потоками".
Мембранный потенциал покоя образуется за счёт двух процессов:
1. Работа калий-натриевого насоса мембраны.
Новая гипотеза механизма работы Na,K-АТФазы рассматривается здесь: Механизм натрий-калиевого насоса
Работа калий-натриевого насоса, в свою очередь, имеет 2 следствия:
1.1. Непосредственное электрогенное (порождающее электрические явления) действие ионного насоса-обменника. Это создание небольшой электроотрицательности внутри клетки (-10 мВ).
Виноват в этом неравный обмен натрия на калий. Натрия выбрасывается из клетки больше, чем поступает в обмен калия. А вместе с натрием удаляется и больше "плюсиков" (положительных зарядов), чем возвращается вместе с калием. Возникает небольшой дефицит положительных зарядов. Мембрана изнутри заряжается отрицательно (примерно -10 мВ).
1.2. Создание предпосылок для возникновения большой электроотрицательности.
Эти предпосылки - неравная концентрация ионов калия внутри и снаружи клетки. Лишний калий готов выходить из клетки и выносить из неё положительные заряды. Об этом мы скажем сейчас ниже.
2. Утечка ионов калия из клетки.
Из зоны повышенной концентрации внутри клетки ионы калия выходят в зону пониженной концентрации наружу, вынося заодно положительные электрические заряды. Возникает сильный дефицит положительных зарядов внутри клетки. В итоге мембрана дополнительно заряжается изнутри отрицательно (до -70 мВ).
Финал
Калий-натриевый насос создает предпосылки для возникновения потенциала покоя. Это - разность в концентрации ионов между внутренней и наружной средой клетки. Отдельно проявляет себя разность концентрации по натрию и разность концентрации по калию. Попытка клетки выравнять концентрацию ионов по калию приводит к потере калия, потере положительных зарядов и порождает электроотрицательность внутри клетки. Эта электроотрицательность составляет большую часть потенциала покоя. Меньшую его часть составляет непосредственная электрогенность ионного насоса, т.е. преобладающие потери натрия при его обмене на калий.
Видео: Мембранный потенциал покоя (Resting membrane potential)
Все клетки живого организма поляризованы. Между внутренней средой клетки и внешней средой существует разность потенциалов, которая носит название мембранного потенциала. В случае возбудимых тканей он называется потенциалом покоя. Его величина в разных тканях неодинакова.
Существование животного электричества открыл Л.Гальвани в 1791 г. Гипотеза, объясняющая происхождение мембранного потенциала была выдвинута Чаговцем в 1896 г, получила дальнейшее развитие в трудах Бернштейна ( 1902 г) и превратилась в теорию после экспериментальной проверки группой экспериментаторов ( Ходжкин, Катц, Хаксли 1949-1952 гг).
Согласно этой теории, потенциал покоя возникает вследствие неравномерного распределения ионов внутри клетки по от- ношению к внешней среде, а также селективных свойств мембраны.
Селективность мембраны обусловливает ее важнейшее свойство – полупроницаемость. В состоянии покоя проницаемость для калия, натрия и хлора неодинаковы:
Р калия : Р натрия : Р хлора = 1 : 0,04 : 0, 45
В связи с наилучшей проницаемостью мембраны и выраженным концентрационным градиентом, катионы калия выходят через калиевые каналы из клетки. При этом клетка теряет положительно заряженные ионы и приобретает отрицательный заряд. Силами электростатического взаимодействия катионы калия удерживаются на наружной стороне мембраны, являясь материальным носителем ее положительного заряда. Носителями отрицательного заряда внутренней стороны мембраны являются крупные органические анионы.

По мере выхода калия клетка все больше поляризуется. При этом все больше нарастает сила, обусловленная электрическим градиентом, препятствующая выходу положительно заряженных ионов калия. В со- стоянии покоя выходящий ток калия (по концентрационному градиенту) и входящий ток ( по электрохимическому градиенту ) уравновешиваются и мембранный потенциал становится стабильным от -60 до -80 мв.
В зависимости от его величины мембрана может быть поляризована (величина мембранного потенциала равна потенциалу покоя), деполяризована (мембранный потенциал меньше потенциала покоя), гиперполяризована (мембранный потенциал больше потенциала покоя).
Возможные изменения мембранного потенциала будут возникать или при нарушении градиентов, или при изменениях проницаемости мембраны ( наиболее распространенная ситуация ). Для катионов калия можно рассчитать по формуле Нернста равновесный калиевый потенциал:

Свой вклад в потенциал покоя вносят другие потенциалобразующие ионы ( натрий, хлор, кальций ). Для каждого из них можно рассчитать равновесный потенциал по формуле Нернста. Суммарная величина мембранного потенциала приближается к сумме равновесных протенциалов основных потенциалобразующих ионов.

В процессе поляризации мембраны клетка теряет катионы калия и получает ионы натрия и хлора, однако нарушения ионного градиента не происходит. Обеспечение постоянства последнего связано с деятельностью механизмов активного транспорта ионов ( ионных насосов). Перенос калия внутрь клетки и натрия наружу обеспечивается котранспортом этих ионов.
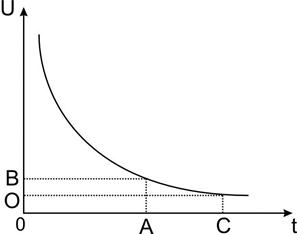
Основной перенос осуществляется калий- натриевым насосом (АТФазой) . Этот механизм является электрогенным, поскольку на 2 катиона калия, переносимого внутрь клетки, наружу переносится 3 катиона натрия. Тем самым происходит увеличение разности потенциалов клеточной мембраны (до 25% от общей величины потенциала покоя), рис. 10.
Читайте также: