
Механизм транспорта лекарственных веществ через клеточные мембраны кратко
Обновлено: 05.07.2024
Транспортные структуры мембраны
Транспортные структуры мембраны - это специальные белковые структуры, встроенные в мембрану и обеспечивающие трансмембранный транспорт, т.е. перенос веществ через мембрану.
Мы уже сказали в предыдущем разделе 1_1 Строение клеточной мембраны, что многие вещества могут проходить сквозь клеточную мембрану самостоятельно, за счёт простой диффузии. Они "автоматически" перемещаются из зоны своей повышенной концентрации в зону пониженной концентрации, т. к. стремятся уравнять свою концентрацию по обе стороны мембраны. Так обычно ведут себя все вещества в растворах: они стремятся равномерно распределиться по всему объёму жидкости, пытаясь преодолеть разделяющую раствор на отсеки преграду. В принципе, любая молекула может пройти через липидный бислой клеточной мембраны, потому что составляющие его молекулы липидов сохраняют некоторую подвижность относительно друг друга и могут временами раздвигаться в стороны, пропуская различные вещества. Однако скорость такой пассивной диффузии, т.е. перехода вещества через мембрану из области с большей концентрацией в область с меньшей, может сильно различаться для разных веществ. Для многих веществ диффузия занимает столь длительное время, что можно говорить о практической непроницаемости для них мембраны. Скорость диффузии различных веществ через мембрану зависит главным образом от размера их молекул, электрического заряда (полярности) и их относительной растворимости в жирах.
Легче и лучше всего через мембрану пассивно проникают жирорастворимые неполярные мелкие молекулы.
Для транспорта в клетку веществ, слабо способных к диффузии через липидный слой мембраны, необходимы специальные транспортные структуры.
Виды транспортных структур мембраны:
1. Ионные каналы - это специальные молекулярные трубочки с порами (дырочками) в мембране, образованные канальными белками, позволяющие ионам проходить через мембрану в обоих направлениях: как внутрь, так и наружу. Ионные каналы могут открываться при определённых условиях, в этом случае они являются управляемыми этими условиями. Через открытую пору ионного канала ионы могут свободно диффундировать, не вызывая конформационных изменений в белке канала. При токе через отдельный канал в 1 пА по нему проходят 6×10 6 одновалентных ионов в секунду.
2. Транслоказы, - специальные мембранные белки, облегчающие переход вещества через мембрану за счёт своего временного связывания с диффундирующим веществом. Не требуют энергии, работают в обоих направлениях в зависимости от концентрации переносимого вещества. Раньше их представляли себе как некие "белки-переносчики" в виде "челноков", но когда стали известны аминокислотные последовательности транспортных белков, то стало понятно, что все они обычно содержат многочисленные трансмембранные домены и образуют сквозной путь через мембрану. Транспортёр, связываясь с субстратом по одну сторону мембраны, изменяет свою конформацию для переноса субстрата через мембрану. При ко-транспорте, в дополнение к сказанному, повышается аффинность транспортёра в новой конформации ко второму субстрату, и субстраты переносятся через мембрану сопряженно, совместно.
3. Транспортёры - белковые структуры, насильно протаскивающие определённые вещества сквозь клеточную мембрану в определённом направлении с затратами энергии. Ионные насосы - это транспортёры ионов. По способу использования энергии для своей работы транспортёры можно разделить на "симпортные" и "антипортные". Симпортные транспортёры используют совместный транспорт в одном направлении двух веществ: одно из них должно иметь большую потенциальную энергию для движения через мембрану. Например, симпорт в клетку с помощью ионов натрия глюкозы, или симпорт ионов кальция с помощью ионов натрия. Антипортные транспортёры (обменники) используют встречный транспорт двух веществ с разной потенциальной энергией диффузии. Так работает, например, натрий-калиевый ионный насос.
Итак, перенос веществ через клеточную мембрану происходит различными путями.
Механизмы транспорта веществ через мембрану
1. Простая диффузия жирорастворимых (гидрофобных) веществ через жировой слой мембраны. Это пассивный процесс под действием градиента (перепада) концентрации вещества по разные стороны мембраны. (Смотрите видео 1 : пассивный транспорт через мембрану).
2. Неуправляемая диффузия (неуправляемый пассивный перенос) водорастворимых веществ через постоянно открытые ионные каналы мембраны.
3. Управляемая диффузия (управляемый пассивный перенос) водорастворимых веществ через управляемые ионные каналы мембраны. (Смотри: Ионные каналы мембраны)
4. Активный транспорт водорастворимых веществ с помощью специальных белковых транспортных структур (транспортёров) за счёт использования энергии расщепления АТФ. (Смотрите видео 2 : активный транспорт через мембрану).
5. Эндоцитоз крупных частиц за счёт образования мембранных пузырьков.
Видео 3 : Эндоцитоз и экзоцитоз
Видео 4 : Фагоцитоз (эндоцитоз) бактерий макрофагом
Как это происходит? Вот этот вопрос мы сейчас и рассмотрим.
Видеоанимация 5 : Пассивный мембранный транспорт
Видео 6 : Мембрана и транспорт веществ
Видео 7 : Мембранный транспорт
1. Транспортёры мембраны - ферменты по имени АТФазы
Одна из самых главных транспортных структур мембраны — это фермент АТФаза (произносится как "а\тэ\эф\аза"). АТФазы разных видов транспортируют через мембрану ионы. Они переносят их как внутрь клетки, так и, наоборот, наружу. Поэтому их называют "ионными насосами" или "ионными помпами". Электрическая проводимость ионных насосов очень мала: Na/K-АТФаза переносит всего около 300 Na + и 200 K + в секунду.
Название АТФаза означает, что это фермент, нацеленный на расщепление АТФ, его полное название - аденозинтрифосфатаза.
Только не надо думать, что АТФаза существует в единственном варианте! В настоящее время уже обраружено множество различных видов транспортных АТФаз. Они схожи между собой по строению и механизму действия, но имеют разную специализацию, т.е. каждый их вид перетаскивает через мембрану что-то своё. В настоящее время достаточно хорошо изучены Na + /K + -АТФаза, Ca 2+ -АТФаза, H + -АТФаза, H + ,K + -АТФаза, Mg 2+ -АТФаза, которые обеспечивают перемещение соответственно ионов Na + , K + , Ca 2+ , H + , Mg 2+ изолированно или сопряжённо: например, Na + сопряжённо с К + ; Н + сопряжённо с К + .
В чём принцип работы АТФазы?
Самой главной мембранной АТФазой по праву можно считать Na,K-АТФазу (натрий-калиевую аденозинтрифосфатазу).

По своей структуре она является представителем гетеродимерных АТФаз Р-типа.
При этом следует помнить, что, как истинный фермент, Na,K-АТФаза параллельно расщепляет АТФ, получая от этого энергию на свою транспортную деятельность.
Далее цикл повторяется.
Гипотеза работы Na,K-АТФазы рассмотрена подробнее здесь: Механизм натрий-калиевого насоса
Какие есть ещё АТФазы?
Н + ,К + -АТФазы обеспечивают секрецию соляной кислоты париетальными клетками желудка. Они перемещают на наружную сторону мембраны ионы водорода, которые создают кислую среду в желудке. Этот транспорт тоже работает по принципу обменника, т.к. меняет внутриклеточные ионы водорода на внеклеточные ионы калия. Кстати, в мембрану боковой и базальной поверхности этих клеток встроен хлорно-бикарбонатный анионообменни, через который анионы: Cl - вводятся в клетку в обмен на HCO3 -
Н + -АТФаза растений обеспечивает поглощение из почвы солей корнями растений. Принцип действия тот же: обмен одних ионов на другие за счёт энергии, полученной из АТФ. Из клеток корня в почву выделяются ионы водорода Н+, а на их место в клетку переносятся ионы солей.
Протонная АТФаза грибка Neurospora crassa состоит из 920 аминокислот (источник: Hager et al. 1986).
Са 2+ -АТФаза саркоплазматического ретикулюма в мышечных клетках обеспечивает транспорт кальция из цитоплазмы мышечных клеток во внутриклеточные цистерны для депонирования (запасания) кальция.

Выводы
Специальные транспортные ферменты АТФазы, встроенные в клеточную мембрану, работают как транспортёры для различных веществ. Они насильно переносят вещества в клетку и из клетки. При этом АТФазы получают на свой активный транспорт энергию за счёт расщепления АТФ.
Активный транспорт веществ через мембрану клетки - это насильственный перенос вещества с затратой энергии. Он возможен даже против градиента концентрации вещества, т.е. из зоны пониженной концентрации в зону повышенной концентрации.
На рисунке справа - структура кальциевой АТФазы по Toyoshima et al. Nature 405 (2000) 647-655 PDB ID: 1EUL.pdb
2. Транспортёры глюкозы
Глюкозные транспортёры - это белки, переносящие глюкозу через мембрану. Их называют белками-переносчиками, а также рецепторами глюкозы. Эти белки образуют гидрофильные трансмембранные каналы.
Глюкозные трнанспортёры делятся на две группы.
1. Na + -глюкозные ко-транспортёры (симпортёры). Эти транспортёры занимаются активным транспортом глюкозы с помощью ионов Na + и их работа зависит от градиента концентрации натрия. Они работают только в почечных канальцах и кишечнике, обеспечивая всасывание глюкозы против градиента её концентрации.
2. Транспортные белки семейства ГЛЮТ. Они отличаются от сходных по функции белков, транспортирующих глюкозу через мембрану в кишечнике и почках, и обеспечивают облегчённую диффузию, а не активный транспорт. Белки ГЛЮТ обнаружены во всех тканях и их существует несколько разновидностей.
Все 5 типов ГЛЮТ имеют сходную первичную структуру и доменную организацию.
- ГЛЮТ-1 (эритроцитарный тип) обеспечивает стабильный поток глюкозы в глиальные клетки мозга.
- ГЛЮТ-2 (печёночный тип) обнаружен в клетках органов, выделяющих глюкозу в кровь. Именно при участии ГЛЮТ-2 глюкоза переходит в кровь из энтероцитов и печени. ГЛЮТ-2 участвует в транспорте глюкозы в β-клетки поджелудочной железы. В то же время ГЛЮТ-2 обеспечивает проникновение глюкозы из крови в клетки печени (гепатоциты) по механизму облегчённой диффузии. Там глюкоза превращается в активное вещество глюкозо-6-фосфат, участвующее в обмене углеводов, жиров и в энергообмене.
- ГЛЮТ-3 (мозговой тип) обладает большим, чем ГЛЮТ-1, сродством к глюкозе. Он также обеспечивает постоянный приток глюкозы к клеткам нервной и других тканей.
- ГЛЮТ-4 (мышечно-жировой тип) - главный переносчик глюкозы в клетки мышц и жировой ткани. Это единственный переносчик, регулируемый инсулином, поэтому мышцы и жировую ткань называют инсулинзависимыми тканями.
- ГЛЮТ-5 (кишечный тип) встречается, главным образом, в клетках тонкого кишечника.
Влияние инсулина на транспортёры ГЛЮТ
Все типы ГЛЮТ могут находиться как в наружной мембране клетки, так и в цитозольных везикулах. ГЛЮТ-4 (и в меньшей мере ГЛЮТ-1) почти полностью находятся в цитоплазме клеток. Влияние инсулина на такие клетки приводит к перемещению везикул, содержащих ГЛЮТ, к плазматической мембране, слиянию с ней и встраиванию этих белков-транспортёров в мембрану. После этого становится возможным облегчённый транспорт глюкозы в эти клетки. Скорость потребления глюкозы возрастает в 30-40 раз. После снижения концентрации инсулина в крови транспортёры глюкозы снова перемещаются в цитоплазму, и поступление глюкозы в клетку прекращается.
Видео 8 : Транспорт веществ в клетку
Видео 9 : Виды клеточного транспорта (англ.яз.)
На всех этапах своего пребывания в организме — всасывания, распределения, метаболизма, экскреции — лекарственные средства проникают через клеточные мембраны. Поэтому очень важно понимать механизм мембранного транспорта, знать физико-химические свойства препарата и самой мембраны, от которых этот транспорт зависит. Так, решающее значение имеют размеры и форма молекулы лекарственного средства, степень ионизации, растворимость в жирах и связывание с белками тканей.
Клеточная мембрана — наиболее универсальный биологический барьер, который приходится преодолевать лекарственным средствам. Он входит в состав других биологических барьеров, таких, как слизистая кишечника, состоящая из одного слоя клеток, или кожа, состоящая из нескольких таких слоев. В большинстве случаев лекарственные средства переносятся через клетки, а не между ними, поэтому механизм проникновения через столь разные по строению барьеры во многом сходен.
Клеточная мембрана представляет собой двойной слой амфифильных липидов, гидрофобные углеводородные хвосты которых обращены внутрь, а гидрофильные головки — наружу. Мембраны разных клеток различаются по липидному составу. Перемещаясь вдоль поверхности, липидные молекулы придают мембране текучесть и эластичность. Кроме того, мембраны обладают высоким электрическим сопротивлением и плохо проницаемы для полярных молекул. Встроенные в двойной липидный слой мембранные белки выполняют функции рецепторов, ионных каналов и переносчиков, участвуя в восприятии и передаче химических и электрических сигналов. Эти белки служат мишенями для лекарственных средств.
При преодолении биологических барьеров лекарственные средства могут проходить как сквозь клетки, так и между ними. Между эндотелиальными клетками большинства капилляров препараты транспортируются с током воды, происходящим путем фильтрации по градиенту гидростатического давления. Такой вид транспорта доступен только свободным лекарственным средствам, так как молекулы белков, с которыми они связаны, слишком велики. Транспорт веществ через межклеточные контакты осуществляется довольно легко. В капиллярах он зависит только от интенсивности кровотока (см. ниже). Этот вид транспорта играет важную роль в проникновении веществ через почечный фильтрационный барьер. В некоторых органах и тканях (капилляры ЦНС, многие эпителиальные ткани) клетки связаны плотными контактами, транспорт через которые ограничен (см. ниже). При прохождении сквозь клетки лекарственные средства должны переноситься через клеточную мембрану. Последняя хорошо проницаема для воды, которая проникает через мембрану путем диффузии по градиенту осмотического давления. Вместе с водой через мембрану могут проходить небольшие (молекулярная масса не более 100—200), растворимые в воде молекулы лекарственных средств. Большинство липофильных препаратов с высоким молекулярным весом транспортируются путем активного или пассивного транспорта.
Многие липофильные лекарственные средства проходят через клеточную мембрану посредством простой диффузии (разновидность пассивного транспорта) по градиенту концентрации. Скорость простой диффузии прямо пропорциональна величине трансмембранного концентрационного градиента, коэффициенту распределения препарата между липидной и водной фазами и площади диффузионной поверхности. Чем больше коэффициент распределения, тем выше концентрация препарата в мембране и тем выше скорость диффузии. В случае неионизированных веществ по достижении стационарного состояния концентрации свободного вещества по обе стороны мембраны выравниваются. Трансмембранное распределение ионизированных веществ зависит от электрохимических градиентов для ионов и от градиента pH между внутри- и внеклеточным пространством, так как величина pH влияет на степень ионизации.
Большинство лекарственных средств представляют собой слабые кислоты или слабые основания, которые могут находиться в водном растворе как в ионизированной, так и в неионизированной форме. Неионизированные молекулы обычно липофильны и легко проходят через мембрану (неионная диффузия), в отличие от плохо растворимых в жирах ионизированных молекул.
Трансмембранное распределение слабых электролитов зависит от показателя кислотной диссоциации pКа (рКа соответствует pH, при котором половина всех молекул слабой кислоты или слабого основания ионизирована) и градиента pH. На рис. 1.2 представлено распределение слабой кислоты (рКа 4,4) между плазмой (pH 7,4) и желудочным соком (pH 1,4). Слизистая желудка выполняет функцию липидного барьера, проницаемого только для неионизированных, липофильных молекул. Соотношение неионизированной и ионизированной форм препарата легко вычислить с помощью уравнения Гендерсона—Гассельбальха. В плазме это соотношение равно 1:1000, а в желудочном соке — 1:0,001 (на рисунке эти данные приведены в квадратных скобках, а смещение равновесия в сторону ионизированной или неионизированной Формы указано толстыми горизонтальными стпелками). Таким образом, в стационарном состоянии соотношение концентраций препарата в плазме и желудочном соке составит 1000:1. Для слабого основания с рКа 4,4 указанные соотношения обратные, и равновесие между ионизированной и неионизированной формами также смещено в обратную сторону. Итак, в стационарном состоянии слабая кислота накапливается преимущественно с той стороны мембраны, где среда более щелочная, а слабое основание — с той стороны, где среда более кислая (так называемая ионная ловушка). Эти закономерности во многом определяют всасывание и экскрецию лекарственных средств (см. ниже). Разность концентраций слабого электролита по обе стороны биологических барьеров устанавливается без затрат энергии, за счет избирательной проницаемости мембраны для неионизированных молекул и трансмембранного градиента pH. Последний, впрочем, создается активным транспортом ионов.
Хотя наиболее распространенный механизм мембранного транспорта лекарственных средств — простая диффузия, препараты могут проходить через клеточную мембрану с участием белков-переносчиков — путем облегченной диффузии или активного транспорта. Облегченная диффузия (разновидность пассивного транспорта) идет по концентрационному или электрохимическому градиенту без затрат энергии, но, в отличие от простой диффузии, осуществляется с помощью белка-переносчика. Активный транспорт идет против концентрационного либо электрохимического градиента и требует затрат энергии. Облегченная диффузия и активный транспорт характеризуются насыщением, специфичностью и конкурентным ингибированием. Белки-переносчики участвуют в переносе через мембрану эндогенных веществ, скорость простой диффузии которых слишком мала, и могут обладать высокой специфичностью к определенной конформации молекулы лекарственного вещества. Кроме того, с помощью белков-переносчиков из клетки выводятся токсичные вещества.
От локализации переносчика в той или иной части клетки часто зависит направление переноса (в клетку или из нее), что во многих случаях обеспечивает направленный транспорт лекарственных средств через клеточные слои. Так, переносчики в базолатеральной мембране гепатоцитов транспортируют желчные кислоты и амфифильные органические ионы в клетку, а системы активного транспорта в апикальной мембране — из клетки в желчь. Сходные механизмы действуют в кишечнике и почечных канальцах. Во всех этих органах, а также в эндотелии капилляров головного мозга присутствует Р-гликопротеид — мембранный белок-переносчик, отвечающий за выведение лекарственных средств из клетки. Этот белок кодируется геном ABCВ1 (старое название MDR- 1) и обусловливает устойчивость опухолевых клеток к химиотерапии (гл. 52). В кишечнике Р-гликопротеид транспортирует лекарственные средства из энтеропита обратно в просвет кишечника, уменьшая их всасывание.
Пассивный транспорт – транспорт веществ через мембрану, осущестляемый без затрат энергии.
1. Простая диффузия. Небольшие нейтральные молекулы (Н2О, СО2, О2, NH3, мочевина, этанол, гидрофобные низкомолекулярные органические вещества (стероидные гормоны, бензол)) диффундируют через мембрану без участия специальных механизмов. Перенос веществ осуществляется по градиенту концентрации и с низкой скоростью (рис. 27, 1).
2. Облегченная диффузия. Для более крупных полярных молекул (глюкоза, аминокислоты), а также для ионов липидный бислой практически непроницаем, так как его внутренняя часть гидрофобна. Такие вещества переносятся через мембрану также по градиенту концентрации, но с участием мембранных белков.
2а. Перенос с участием ионных каналов. Трансмембранный перенос ряда ионов (Са 2+ , Na + , K + , C1 − ) происходит через ионные каналы - белковые структуры, пронизывающие мембрану. Они образуют трансмембранный гидрофильный (заполненный водой) канал. Избирательность каналов к ионам определяется наличием в белках канала специфического центра связывания иона. Каналы могут быть или закрыты, или открыты. Сигналом для изменения состояния канала являются гормон или иная сигнальная молекула
(рис. 27, 2).
2б. Перенос с помощью трансмембранных белков-переносчиков (транслоказ). Для каждого вещества или группы сходных веществ имеется свой переносчик. Переносимое вещество присоединяется к транслоказе, в результате чего изменяется ее конформация, в мембране открывается канал, и вещество освобождается с другой стороны мембраны. Поскольку в канале нет гидрофобного препятствия, то этот механизм называют облегченной диффузией (рис. 27, 3). Пример - облегченная диффузия (унипорт) глюкозы в эритроциты с помощью ГЛЮТ-1.
 |
| Рис. 27. Перенос веществ через мембрану: 1 - пассивная диффузия, 2 - диффузия с помощью ионного канала, 3 - диффузия с помощью белка-переносчика, 4 - активный транспорт |
Активный транспорт веществ протекает против концентрационного градиента и требует затрат энергии (рис. 27, 4). Таким способом происходит перенос многих минеральных ионов из межклеточной жидкости в клетку или в обратном направлении, перенос аминокислот из просвета кишечника в клетки кишечника, перенос глюкозы из первичной мочи через клетки канальцев почки в кровь. Основным источником энергии для активного транспорта является АТФ. Поэтому, как правило, эти системы представляют собой АТФазы.
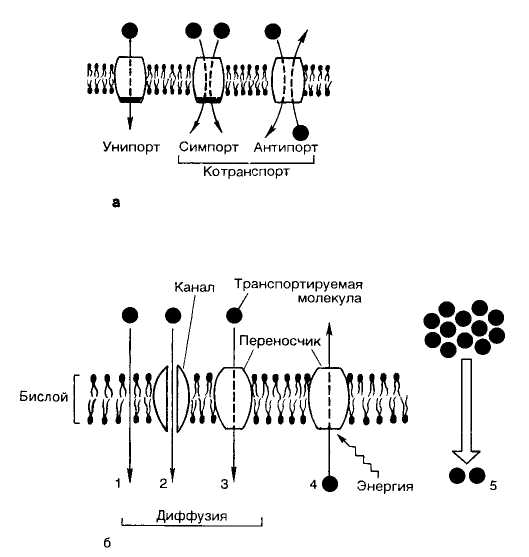
В зависимости от количества веществ, переносимых через один белок-переносчик, и направления транспорта, различают:
унипорт - транспорт одного вещества;
симпорт - транспорт двух веществ в одном направлении через один переносчик;
антипорт - перемещение двух веществ в разных направлениях через один переносчик (рис. 28).
Рис. 28. Виды переноса веществ через мембрану
Эта классификация справедлива как для пассивного, так и для активного транспорта. Примером пассивного антипорта является перенос ионов хлора и гидрокарбонат-ионов.
Примером активного антипорта яляется натрий–калиевая АТФаза. Она переносит в клетку ионы калия, а из клетки - ионы натрия.
АТФ-аза присоединяет с внутренней стороны мембраны три иона Na + . Эти ионы изменяют конформацию активного центра АТФазы, и она гидролизует одну молекулу АТФ, присоединяя к себе фосфат. Выделившаяся энергия расходуется на изменение конформации АТФазы, после чего три иона натрия оказываются на внешней стороне мембраны, а фосфат замещается на 2 иона K + из внешней среды. Затем конформация переносчика изменяется на первоначальную, и ионы K + оказываются на внутренней стороне мембраны. Здесь ионы K + отщепляются.
Работа Na + ,K + -ATФазы создает не только разность концентраций ионов, но и потенциал на мембране. На внешней стороне мембраны создается положительный заряд, на внутренней - отрицательный.
Вторично-активный транспорт. Градиент одного вещества используется для транспорта другого. Переносчик в этом случае имеет специфические центры связывания для обоих веществ. Вещество транспортируется противградиента своей концентрации путем симпорта или антипорта. Симпорт и антипорт, например, могут происходить за счет энергии градиента концентрации ионов Na + , создаваемого Na + ,K + -ATФазой. Таким способом происходит всасывание аминокислот из кишечника и глюкозы из первичной мочи икишечника.

Пример вторично-активного симпорта – транспорт глюкозы и ионов натрия; вторично-активного антипорта – транспорт ионов кальция и натрия.
Для переноса углеводов, аминокислот и других метаболитов вторично-активный транспорт имеет, по-видимому, наибольшее значение по сравнению с другими механизмами.
Структура и функции мембран нарушаются при ряде заболеваний.
Контрольные вопросы
1. Перечислите основные мембранные структуры клетки.
2. Каков качественный и количественный состав мембран?
3. Какие липиды входят в состав биологических мембран? Каковы их свойства и функции?
4. В чем различие между периферическими и интегральными белками биологических мембран?
5. Перечислите функции биологических мембран.
6. Какие факторы могут вызвать изменение структуры и проницаемости мембраны?
7. Перечислите виды транспорта веществ через мембраны. Какие из них требуют энергетических затрат?
8. Почему перенос ионов даже по градиенту концентрации происходит только с участием ионных каналов?
9. Какую роль играют АТФ-азы в функционировании биологических мембран?
10. Изобразите схему работы натрий-калиевой АТФазы.
Пассивный транспорт – транспорт веществ через мембрану, осущестляемый без затрат энергии.
1. Простая диффузия. Небольшие нейтральные молекулы (Н2О, СО2, О2, NH3, мочевина, этанол, гидрофобные низкомолекулярные органические вещества (стероидные гормоны, бензол)) диффундируют через мембрану без участия специальных механизмов. Перенос веществ осуществляется по градиенту концентрации и с низкой скоростью (рис. 27, 1).
2. Облегченная диффузия. Для более крупных полярных молекул (глюкоза, аминокислоты), а также для ионов липидный бислой практически непроницаем, так как его внутренняя часть гидрофобна. Такие вещества переносятся через мембрану также по градиенту концентрации, но с участием мембранных белков.
2а. Перенос с участием ионных каналов. Трансмембранный перенос ряда ионов (Са 2+ , Na + , K + , C1 − ) происходит через ионные каналы - белковые структуры, пронизывающие мембрану. Они образуют трансмембранный гидрофильный (заполненный водой) канал. Избирательность каналов к ионам определяется наличием в белках канала специфического центра связывания иона. Каналы могут быть или закрыты, или открыты. Сигналом для изменения состояния канала являются гормон или иная сигнальная молекула
(рис. 27, 2).
2б. Перенос с помощью трансмембранных белков-переносчиков (транслоказ). Для каждого вещества или группы сходных веществ имеется свой переносчик. Переносимое вещество присоединяется к транслоказе, в результате чего изменяется ее конформация, в мембране открывается канал, и вещество освобождается с другой стороны мембраны. Поскольку в канале нет гидрофобного препятствия, то этот механизм называют облегченной диффузией (рис. 27, 3). Пример - облегченная диффузия (унипорт) глюкозы в эритроциты с помощью ГЛЮТ-1.
|
| Рис. 27. Перенос веществ через мембрану: 1 - пассивная диффузия, 2 - диффузия с помощью ионного канала, 3 - диффузия с помощью белка-переносчика, 4 - активный транспорт |
Активный транспорт веществ протекает против концентрационного градиента и требует затрат энергии (рис. 27, 4). Таким способом происходит перенос многих минеральных ионов из межклеточной жидкости в клетку или в обратном направлении, перенос аминокислот из просвета кишечника в клетки кишечника, перенос глюкозы из первичной мочи через клетки канальцев почки в кровь. Основным источником энергии для активного транспорта является АТФ. Поэтому, как правило, эти системы представляют собой АТФазы.
В зависимости от количества веществ, переносимых через один белок-переносчик, и направления транспорта, различают:
унипорт - транспорт одного вещества;
симпорт - транспорт двух веществ в одном направлении через один переносчик;
антипорт - перемещение двух веществ в разных направлениях через один переносчик (рис. 28).
Рис. 28. Виды переноса веществ через мембрану
Эта классификация справедлива как для пассивного, так и для активного транспорта. Примером пассивного антипорта является перенос ионов хлора и гидрокарбонат-ионов.
Примером активного антипорта яляется натрий–калиевая АТФаза. Она переносит в клетку ионы калия, а из клетки - ионы натрия.
АТФ-аза присоединяет с внутренней стороны мембраны три иона Na + . Эти ионы изменяют конформацию активного центра АТФазы, и она гидролизует одну молекулу АТФ, присоединяя к себе фосфат. Выделившаяся энергия расходуется на изменение конформации АТФазы, после чего три иона натрия оказываются на внешней стороне мембраны, а фосфат замещается на 2 иона K + из внешней среды. Затем конформация переносчика изменяется на первоначальную, и ионы K + оказываются на внутренней стороне мембраны. Здесь ионы K + отщепляются.
Работа Na + ,K + -ATФазы создает не только разность концентраций ионов, но и потенциал на мембране. На внешней стороне мембраны создается положительный заряд, на внутренней - отрицательный.
Вторично-активный транспорт. Градиент одного вещества используется для транспорта другого. Переносчик в этом случае имеет специфические центры связывания для обоих веществ. Вещество транспортируется противградиента своей концентрации путем симпорта или антипорта. Симпорт и антипорт, например, могут происходить за счет энергии градиента концентрации ионов Na + , создаваемого Na + ,K + -ATФазой. Таким способом происходит всасывание аминокислот из кишечника и глюкозы из первичной мочи икишечника.
Пример вторично-активного симпорта – транспорт глюкозы и ионов натрия; вторично-активного антипорта – транспорт ионов кальция и натрия.
Для переноса углеводов, аминокислот и других метаболитов вторично-активный транспорт имеет, по-видимому, наибольшее значение по сравнению с другими механизмами.
Структура и функции мембран нарушаются при ряде заболеваний.
Контрольные вопросы
1. Перечислите основные мембранные структуры клетки.
2. Каков качественный и количественный состав мембран?
3. Какие липиды входят в состав биологических мембран? Каковы их свойства и функции?
4. В чем различие между периферическими и интегральными белками биологических мембран?
5. Перечислите функции биологических мембран.
6. Какие факторы могут вызвать изменение структуры и проницаемости мембраны?
7. Перечислите виды транспорта веществ через мембраны. Какие из них требуют энергетических затрат?
8. Почему перенос ионов даже по градиенту концентрации происходит только с участием ионных каналов?
• Существует два основных типа мембранных транспортных белков: каналы и переносчики
• Ионные каналы катализируют быстрый и избирательный транспорт ионов по электрохимическому градиенту
• Транспортеры и насосы представляют собой белки-переносчики, которым необходима энергия для транспорта веществ против электрохимического градиента
• В каждой клетке несколько различных мембранных транспортных белков действуют как единое целое
Некоторые из транспортных белков находятся в плазматической мембране, в то время как другие входят в состав мембраны клеточных органелл. Для поддержания определенного состава внутриклеточной среды и клеточных компартментов необходимо, чтобы транспортные белки обладали избирательностью по отношению к определенным метаболитам. В зависимости от характера процесса транспорта мембранные транспортные белки подразделяются на две группы: каналы и переносчики.
Канальные белки содержат поры, через которые при открытом канале с высокой скоростью проходят метаболиты. Белок-переносчик связывает метаболит, находящийся по одну сторону мембраны, после чего конформация его меняется, и метаболит высвобождается с другой стороны мембраны.
Существует несколько типов белков, образующих каналы в мембранах. Порины, присутствующие в клетках некоторых прокариот, в митохондриях и в нексусах, где соединяется цитоплазма соседних клеток, обеспечивают прохождение метаболитов в соответствии с размером их молекул.
К более селективным каналам относятся комплексы ядерных пор и каналы, обеспечивающие перемещение белков через мембрану эндоплазматического ретикулума. В статьях на сайте будут рассмотрены ионные каналы и аквапорины, через которые осуществляется селективный трансмембранный перенос ионов и молекул воды соответственно. К настоящему времени описано более 100 различных типов канальных белков. Все они обладают следующими характеристиками:
• селективностью по отношению к переносимому метаболиту,
• высокой скоростью переноса метаболита и
• воротным механизмом, регулирующим транспорт метаболитов.
Участок канального белка, через который осуществляется трансмембранный транспорт, называется порой канала. Канал может находиться в нескольких различных конфигурациях. Некоторые каналы состоят из одного белка, трансмембранные сегменты которого образуют пору. Другие существуют в виде олигомеров, состоящих из идентичных или разных субъединиц, которые вместе образуют пору.
Наконец существуют каналы, состоящие из двух и более субъединиц, каждая из которых сама образует пору. Часто комплексы олигомеров каналов регулируются или позиционируются на определенных мембранах с участием дополнительных субъединиц.
Большинство канальных белков обладают высокой селективностью в отношении определенных метаболитов, например ионов натрия (Na+), калия (К+), кальция (Са2+), хлорида (Сl-) или воды. Другие канальные белки являются неселективными катионными или анионными каналами. Как будет показано далее, при рассмотрении свойств специфических канальных белков, их поры характеризуются определенными структурными особенностями, т. е. являются селективным фильтром, который позволяет им проявлять избирательность по отношению к различным метаболитам.
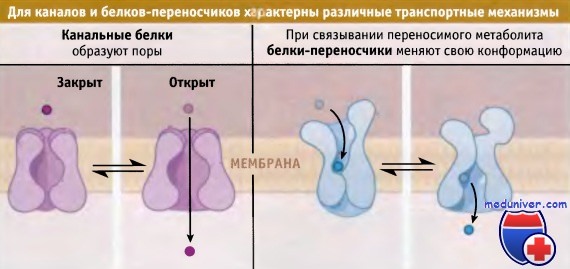
Каналы и переносчики представляют собой два основных типа белков мембранного транспорта.
Растворенные метаболиты с максимальной скоростью диффундируют через пору каналов,
а переносчики связывают метаболит с одной стороны мембраны, затем претерпевают конформационные изменения и высвобождают его с другой стороны.
Этот процесс протекает значительно медленнее.
Направление потока ионов растворенных веществ через канал определяется электрохимическим градиентом. Иными словами, метаболиты движутся через канал в энергетически-выгодном направлении, т. е. по электрохимическому градиенту. Например, для покоящейся клетки, схематически изображенной на рисунке ниже, существует направленный в клетку ионный поток через Na+-, Са2+-, Cl--каналы, и противоположный, проходящий через К+-каналы.
Так как других источников энергии, кроме электрохимического градиента, в данном случае не используется, то такой тип транспорта называется пассивным. Канальные белки обеспечивают высокую скорость транспорта. Так, для ионных каналов скорость составляет 108 в 1 с, что близко к максимальной скорости диффузии ионов в водной среде.
Транспортные мембранные белки контролируют различные клеточные функции при участии воротного механизма. В основе этого механизма лежит индукция конформационных изменений в белке в ответ на воздействие определенного стимула. Эти изменения приводят к открытию или закрытию канала. Например, открытие и закрытие ионных каналов могут регулироваться лигандами, величиной электрического потенциала, механическими или температурными воздействиями. Активация каналов может происходить очень быстро, что делает их идеальным инструментом обработки поступающих в клетку сигналов.
Например, передача нервного импульса зависит от электрического заряда, который создается ионным каналом в мембране. Каналы также играют важную роль в регуляции объема клетки и внутриклеточного pH, в транспорте солей и воды через эпителиальные клетки, в закислении внутриклеточных органелл и во внеклеточной передаче сигналов.
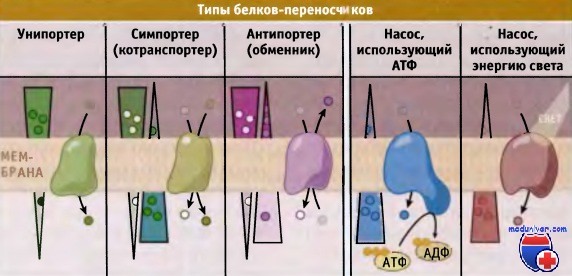
Транспортеры и насосы представляют собой два основных типа белков-переносчиков.
Транспортеры делятся на три типа, каждый из которых осуществляет один из типов переноса: унипорт, симпорт, или антипорт.
Показано направление электрохимического градиента через мембрану.
В зависимости от типа белка-переносчика транспорт может происходить или в направлении градиента (от высокой концентрации к низкой), или против него.
При транспорте субстратов против градиента концентрации, белки-переносчики используют свободную энергию электрохимического градиента, АТФ или других источников. Поскольку при этом происходит потребление энергии, процесс носит название активный транспорт. Белки-переносчики подразделяются на две группы, транспортеры и насосы.
Для транспортировки субстратов через мембрану транспортеры используют энергию электрохимического градиента. Они подразделяются на унипортеры, симпортеры (или котранс-портеры) и антипортеры (обменники). Для энергозависимого транспорта метаболитов мембранные насосы используют энергию, например высвобождающуюся при гидролизе АТФ. По сравнению с канальными белками, переносчики характеризуются более медленной скоростью транспорта, порядка 1000 молекул в 1 с.
Различают два типа активного транспорта, первичный и вторичный. Белки-переносчики, осуществляющие первичный активный транспорт, в качестве источника энергии используют АТФ и транспортируют метаболиты против электрохимического градиента. Таким образом, они помогают поддерживать градиент концентрации метаболитов по обеим сторонам мембран клетки. Са2+-АТФаза и Na+/К+-АТФаза являются характерными примерами транспортных насосов, которые управляют процессом первичного активного транспорта.
Белки-переносчики, которые участвуют во вторичном активном транспорте, непосредственно не используют энергию АТФ. Вместо этого они используют свободную энергию электрохимического градиента, который возникает за счет первичного активного транспорта. Вторичный активный транспорт происходит с участием симпортеров и антипортеров. Механизмы транспорта метаболитов представлены на рисунке ниже.
В клетке все типы мембранных транспортных белков-каналы, транспортеры и насосы-функционируют совместно. Таким образом, работа какого-либо одного типа транспортного белка зависит от работы других. В данной главе мы рассмотрим несколько примеров такого взаимодействия транспортных белков. Например, ионный градиент по обеим сторонам мембраны поддерживается за счет сложных взаимодействий между различными типами транспортных белков. Для правильного функционирования эпителиальных клеток, например почек, слизистой кишечника и легкого, необходимо протекание транспортных процессов с участием различных ионов и метаболитов. Мы также рассмотрим роль нарушений процессов транспорта в развитии различных заболеваний.

Транспорт растворенных метаболитов через мембрану может быть пассивным или активным.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Читайте также:
- Защита граждан подвергшихся воздействию радиации вследствие катастрофы на чернобыльской аэс кратко
- История моды кратко на английском
- Присоединение сибирского ханства кратко 7 класс история россии
- Концепция информационного общества японского социолога и футуролога енэдзи масуда кратко
- История промышленного дизайна кратко