
Концепция саморегуляции численности популяции кратко
Обновлено: 04.05.2024
Если экосистема – основная функциональная единица экологии, предмет ее исследования, то популяция – основной элемент каждой экосистемы.
Популяция – это совокупность особей одного вида, способная к самовоспроизведению, более или менее изолированная в пространстве и во времени от других аналогичных совокупностей того же вида [27].
Популяция обладает биологическими свойствами, присущими составляющим ее организмам, и групповыми свойствами, присущими только популяции в целом. Как и отдельный организм, популяция растет, дифференцируется и поддерживает сама себя. Но такие свойства, как рождаемость, смертность, возрастная структура, характерны только для популяции в целом.
При описании популяций используют две группы количественных показателей: статические, характеризующие состояние популяции в какой-то определенный момент времени, и динамические, характеризующие процессы, протекающие в популяции за некоторый промежуток времени. Общая численность популяции выражается определенным количеством особей. Для ее оценки применяются различные методы. Если речь идет о крупных и хорошо заметных организмах, применяется аэрофотосъемка. В других случаях применяется метод мечения. Животных ловят, метят и отпускают обратно в природу. Через некоторое время производят новый отлов и по доле меченных животных определяют численность популяции.
Количественным показателем оценки популяции является плотность – численность популяции, отнесенная к единице занимаемого пространства. Для характеристики пространственного распределения особей применяют методы математической статистики, которые позволяют оценить дисперсию наблюдаемого распределения плотности и сопоставить ее со средним значением плотности. При случайном распределении дисперсия равна среднему значению: σ 2 = , при регулярном – дисперсия меньше среднего: σ 2 2 > . В соответствии с этим отношение дисперсии к среднему значению есть показатель степени пространственной агрегированности. Если величина этого показателя примерно равна единице – распределение случайное; меньше единицы – регулярное; больше единицы – пятнистое.
Поскольку длительность существования популяции значительно превышает продолжительность жизни отдельных особей, в ней всегда присутствует смена поколений. И даже если численность популяции постоянна, то постоянство является результатом динамического равновесия прибыли и убыли особей.
Рождаемость – это число особей N, родившихся за некоторый промежуток времени dt – dNn/dt. Для сравнения популяций разной численности величину dNn/dt относят к общему числу особей N в начале промежутка времени dt. Полученную величину dNn/Ndt называют удельной рождаемостью. Единица времени, выбранная для оценки рождаемости, изменяется в зависимости от интенсивности размножения организмов, образующих популяцию. Для популяции бактерий единицей времени может быть час, для планктонных водорослей - сутки, для насекомых - недели или месяцы, для крупных млекопитающих – годы. Смертность – величина противоположная рождаемости. Она оценивается числом особей dNm, погибших за время dt. Удельная смертность выражается как dNm/Ndt.
Продолжительность жизни у разных видов различна, и, чтобы их сравнивать, строятся кривые выживания. На оси абсцисс откладывается время жизни, на оси ординат – число выживших (рис. 1.4). Кривая типа I (сильно выпуклая) характерна для популяций организмов, у которых смертность почти до конца жизни остается низкой. Этот тип кривой выживания характерен для многих видов крупных животных, в том числе и для человека. Другой крайний вариант – кривая типа III (сильно вогнутая) характерна для популяций, у которых смертность высока на ранних стадиях. Так, у личинок устриц и прорастающих желудей очень высокая смертность, но как только особи хорошо приживутся, продолжительность жизни резко увеличивается.

Кривая типа II (диагональная) соответствует постоянной смертности в течение всей жизни. Такие кривые встречаются у рыб, пресмыкающихся и птиц. Кривая выживания человека не всегда имела выпуклую форму. Например, кривые, построенные по надписям на надгробиях людей, живших в Римской империи в I-IV вв. н. э., были диагональными.
В популяции выделяют три экологические возрастные группы: пререпродуктивную, репродуктивную и пострепродуктивную. В быстрорастущих популяциях значительную долю составляют молодые особи. В популяциях, находящихся в стационарном состоянии, возрастное распределение относительно равномерное. В популяциях, численность которых снижается, содержится большая доля старых особей. У современного человека упомянутые три возрастные группы приблизительно одинаковы.
У первобытных людей пререпродуктивный период очень длителен, репродуктивный – короткий, а пострепродуктивный отсутствует совсем. У поденок, например, личиночное развитие занимает несколько лет, а во взрослом состоянии (в репродуктивный период) они живут несколько дней.
В среде, не ограничивающей рост популяции (нет ограничения в пище, пространстве и т.д.), удельная скорость роста, т. е. скорость, рассчитанная на одну особь, становится постоянной и максимальной. Этот показатель, обозначаемый r, является экспонентой в дифференциальном уравнении роста популяции в нелимитирующей среде.
dN/dt = rN; r = dN/Ndt, (1.1)
интегрируя, получаем экспоненциальную зависимость:
Где N0 – численность в начальный момент времени,
Nt – численность в момент времени t,
е – основание натурального логарифма.
Логарифмируя обе части равенства, получаем уравнение в форме, удобной для расчета:
In (Nt) = In (No) + rt, (1.3)
Когда популяция переходит в стационарное состояние, r называют внутренней скоростью естественного роста – биотическим потенциалом и обозначают rmax. Разницу между биотическим потенциалом и скоростью роста в реальных условиях называют сопротивлением среды. Сопротивление среды – это сумма всех лимитирующих факторов, препятствующих реализации rmах.
По форме кривых роста можно выделить два основных типа роста: описываемый J-образной кривой и S-образной (или сигмоидной) кривой (рис. 1.5). При J-образной кривой (рис 1.5 а) плотность популяции быстро возрастает по экспоненте, но затем, когда начинает действовать сопротивление среды, рост быстро прекращается. Этот тип роста может быть описан простым экспоненциальным уравнением (1.1) при заданном ограничении на величину N.

При типе роста, описываемом сигмоидной кривой (рис. 1.5 б), популяция сначала увеличивается медленно, затем рост ее все ускоряется, затем под влиянием среды рост постепенно замедляется и в конце концов достигает равновесия. Этот тип роста описывается простым логистическим уравнением:
где dN/dt - скорость роста популяции,
r - удельная, или внутренняя, скорость роста,
N - величина популяции (численность),
К – максимально возможная величина популяции. Этот предел является верхней асимптотой сигмоидной кривой.
В природе экспоненциальный рост или не происходит вообще, или происходит в течение очень короткого времени. Эта модель используется, чтобы количественно охарактеризовать потенциальные возможности популяции к росту. Она позволяет выявить факторы, ограничивающие рост изучаемой популяции.
Константы r и К из логистического уравнения характеризуют два типа естественного отбора, которые позволяют обосновать разные типы экологических стратегий:
- r-стратегия характерна для популяций в начальный период увеличения ее численности. Она определяется отбором в условиях, когда плотность популяции мала и соответственно слабо выражено тормозящее воздействие конкуренции. Эта стратегия характерна, например, для временных водоемов, заполняющихся водой только в период дождей. r-отбор направлен на высокую плодовитость, быстрое достижение половой зрелости, достижение короткого жизненного цикла, способности выживания в неблагоприятный период в виде покоящихся стадий;
- K-стратегия связана с отбором, направленным на повышение выживаемости и величины предельной плотности К в условиях стабилизирующейся численности популяции при сильном воздействии конкуренции. K-отбор направлен на оценку конкурентоспособности и предусматривает возможные пути защищенности от хищников и паразитов, и выживаемости потомства, а также совершенствования механизмов регуляции численности.
В южных регионах распространено растение амброзия. Она растет по свалкам, залежам и другим недавно нарушенным местообитаниям. С другой стороны, в умеренном поясе в стабильном нижнем ярусе леса обитают травянистые растения. Если сравнить эти растения по продукции семян, окажется, что амброзия продуцирует семян в 50 раз больше, чем растения леса, и тратит в 5 раз больше чистой энергии на размножение. Амброзия – пример r-отбора, растения лесного сообщества – K-отбора.
Выделение r- и K-стратегий в чистом виде условно. На самом деле каждый вид организмов испытывает некую комбинацию r- и K-отбора, т. е. оставляемые отбором особи должны обладать и достаточно высокой плодовитостью, и развитой способностью выживания при наличии конкуренции.
Логистическая модель популяционного роста исходит из предположения о том, что для каждой популяции существует определенный равновесный уровень плотности (численности). Это уровень, при котором рождаемость равна смертности. Если равновесный уровень превышен, то в самой популяции или в окружающей среде что-то должно измениться так, чтобы смертность стала больше рождаемости, а популяция соответственно начала сокращать свою численность. Напротив, в случае понижения численности ниже равновесного уровня процессы, происходящие в популяции или в среде, должны привести к росту численности популяции. Отсюда возник подход к оценке механизмов, поддерживающих численность популяции, называемый регуляционизмом.
Регуляционизм – представление о том, что каждая популяция обладает равновесным уровнем плотности (численности) и существуют механизмы, направленные на поддержание этой плотности. Наблюдения и эксперименты позволили считать, что динамика численности любой популяции есть автоматически регулируемый процесс, а действие факторов, контролирующих популяцию, определяется плотностью самой популяции.
Принципиально иной подход к оценке механизмов, поддерживающих численность популяции, – это стохастизм, при котором придается случайно действующим факторам среды, например, погодным, основное значение. Стохастизм отрицает существование равновесного уровня, отклонение от которого автоматически возвращает популяцию к исходному уровню плотности (численности). С позиции стохастизма равновесный уровень численности есть результат ее усреднения за длительный срок.
Концепция саморегуляции. Сторонники и регуляционизма, и стохастизма, несмотря на различия, сходятся в том, что главная роль в ограничении роста численности популяции принадлежит факторам внешней среды. Однако регуляционисты считают, что факторы среды приводят в действие автоматически регулируемую плотность популяции, стохастисты отводят случайным факторам ведущую роль в определении численности популяции.
В начале 60-х годов нашего столетия была предложена концепция саморегуляции популяций, согласно которой в процессе роста популяции изменяется не только и не столько качество среды, в которой существует эта популяция, сколько качество самих составляющих ее особей. Следовательно, суть концепции саморегуляции состоит в том, что любая популяция способна регулировать свою численность так, чтобы не подрывать возобновляемые ресурсы местообитания, и так, чтобы не потребовалось вмешательства каких-либо внешних факторов, например хищников или неблагоприятной среды.
Основанием для выдвижения этой концепции послужили наблюдения за колониями мышей, содержащихся в лаборатории. В этих условиях при достаточном снабжении пищей возрастание плотности популяции приводило к увеличению у мышей надпочечников – органов эндокринной системы. Гормональные сдвиги, происходящие в организме под влиянием нервного перевозбуждения при перенаселении, ведут к повышению агрессивности животных, изменению репродуктивного потенциала (позднее половое созревание, снижение продуктивности, иногда полное прекращение размножения), снижению устойчивости к заболеваниям. Эти изменения обычно ведут к резкому снижению жизнеспособности особей и их массовой гибели.
Если в лабораторных условиях результатом стрессового состояния, вызванного перенаселенностью, является возрастание смертности, то в природных условиях – миграция в новые местообитания, где больше риск гибели от разнообразных причин.
Учебники для вузов
Экология и безопасность жизнедеятельности
Концепция саморегуляции
Концепция саморегуляции. Сторонники и регуляционизма, и стохастизма, несмотря на различия, сходятся в том, что главная роль в ограничении роста численности популяции принадлежит факторам внешней среды. Однако регуляционисты считают, что факторы среды приводят в действие автоматически регулируемую плотность популяции, стохастисты отводят случайным факторам ведущую роль в определении численности популяции.
В начале 60-х годов нашего столетия была предложена концепция саморегуляции популяций, согласно которой в процессе роста популяции изменяется не только и не столько качество среды, в которой существует эта популяция, сколько качество самих составляющих ее особей. Следовательно, суть концепции саморегуляции состоит в том, что любая популяция способна регулировать свою численность так, чтобы не подрывать возобновляемые ресурсы местообитания, и так, чтобы не потребовалось вмешательства каких-либо внешних факторов, например хищников или неблагоприятной среды.
Основанием для выдвижения этой концепции послужили наблюдения за колониями мышей, содержащихся в лаборатории. В этих условиях при достаточном снабжении пищей возрастание плотности популяции приводило к увеличению у мышей надпочечников – органов эндокринной системы. Гормональные сдвиги, происходящие в организме под влиянием нервного перевозбуждения при перенаселении, ведут к повышению агрессивности животных, изменению репродуктивного потенциала (позднее половое созревание, снижение продуктивности, иногда полное прекращение размножения), снижению устойчивости к заболеваниям. Эти изменения обычно ведут к резкому снижению жизнеспособности особей и их массовой гибели.
Если в лабораторных условиях результатом стрессового состояния, вызванного перенаселенностью, является возрастание смертности, то в природных условиях – миграция в новые местообитания, где больше риск гибели от разнообразных причин.
Факторы, влияющие на численность популяции делят на независимые от плотности и зависимые от плотности.
Факторы, независимые от плотности (его влияние не зависит от величины популяции) – в большинстве случаев погодные условия, бури, пожары, загрязнение среды и т.д.
Факторы, зависимые от плотности – влияние их на популяцию есть функция плотности (в основном биотические факторы – внутривидовая конкуренция, паразиты, патогенные организмы и т.д.). Бывают прямого действия (с увеличением плотности их влияние увеличивается) и обратного.
В экосистемах с низким уровнем разнообразия, находящихся под влиянием физических стрессов, размеры популяции определяются в основном факторами, независимыми от плотности, а внутрипопуляционные факторы играют второстепенную роль. В таких случаях наблюдается отсутствие стабильности в динамике (местности с резко неблагоприятными условиями – аридные зона; мелкие организмы с коротким жизненным циклом – насекомые, планктон и т.д.).
В экосистемах с высоким уровнем разнообразия, функционирующих в благоприятной среде регуляция осуществляется в основном за счет биологических факторов (наблюдается стабильность плотности или постоянство характера изменений ее во времени).
Большинство ученых, изучающих динамику численности популяции, сходились до недавнего времени на том, что ее ведущей силой является нехватка жизненного пространства и ресурсов. Однако в 60-х годах в экологии появилась концепция саморегуляции популяций, согласно которой в процессе роста популяции изменяется не только качество окружающей среды, но и качество особей, составляющих популяцию. Как подчеркнул автор теории Д.Читти (1960), любая популяция способна регулировать свою численность так, чтобы не подрывались возобновляемые ресурсы местообитания и не требовалось вмешательства внешних факторов (хищников, погоды).
Согласно концепции, изменения организмов могут быть фенотипическими и генотипическими.
Фенотипические изменения По наблюдениям за популяции лабораторных мышей, при увеличении плотности их популяции у них возникает состояние стресса, которое характеризуется рядом признаков, в том числе и резким повышением активности надпочечников. Гормональные сдвиги тормозят развитие половых желез, что приводит к более позднему половому созреванию и снижению плодовитости.
Так в одном опыте плодовитость домовых мышей снижалась, если каждый день в течение одной минуты им показывали разъяренного самца.
В природе неоднократно было отмечено, что при увеличении плотности некоторых млекопитающих они часто гибли от “шоковой” болезни.
Имеют место также поведенческие механизмы саморегуляции. Так, при увеличении плотности животных с территориальным поведением все большее их количество не могут удержать свою территорию, мигрируют в менее благоприятные места и чаще гибнут.
Генотипические изменения Специальный генетический механизм саморегуляции подразумевает наличие в популяции минимум двух разных генотипов, один из которых лучше себя чувствует в условиях большей плотности, другой - меньшей. Так в опыте с пенсильванской полевкой показано, что на пике ее численности преобладают особи, выживающие при большей скученности, но с меньшей плодовитостью. При низкой плотности получают распространение особи, с высокой плодовитостью, но не выносящие тесноты.
Крайняя выраженность разнокачественности особей – явление фазовой изменчивости у саранчи (две фазы – одиночная и стадная, различаются морфологически и физиологически. Стадная – яркая окраска, высокая двигательная активность, стремление собираться в собираться в стаи, молодые особи быстрее развиваются, лучше выживают). Массовой миграции могут способствовать климатические условия.
Структура популяции
Возрастная структура популяции
Это важная характеристика популяции, показывающая ее способность к размножению в данный момент и чего можно ожидать в будущем. Для каждой популяции характерны свои соотношения возрастных групп, которые зависят от особей (продолжительность жизни, время достижения половой зрелости, интенсивность размножения и т.д. – вырабатываются в процессе эволюции) и условий среды.
Обычно в молодых быстро растущих популяциях велика доля молодых особей, в популяции, находящейся в стационарном состоянии, возрастное распределение более равномерно, а в популяциях, численность которых снижается, велика доля старых особей.
Возрастные пирамиды. Существуют три типа возрастных пирамид:
Популяции с высокой, умеренной и малой долей молодых особей
Если достигнуто стабильное возрастное распределение, временные отклонения от него могут быть вызваны лишь необычными возрастаниями рождаемости и смертности, после чего происходит самопроизвольный возврат к стабильной ситуации.
По мере развития человеческого общества постепенно переходит переход от пирамиды типа а к типу б и в.
Правило стабильности возрастной структуры популяции: любая популяция стремиться к стабильной возрастной структуре, четкому количественному распределению особей по возрастам (справедливо для высших организмов).
В популяциях животных выделяется три экологические возрастные группы: пререпродуктивная, репродуктивная, пострепродуктивная (у растений – стадия первичного покоя (семена), предгенеративная, генеративная, посгенеративная). Длительность этих возрастов варьирует у разных организмов.
У современного человека эти периоды примерно равны, у первобытного пострепродуктивный период был короче.
Цикада имеет личиночную стадию около 17 лет, а взрослые насекомые живут менее одного сезона.
Соотношение численности разных возрастных групп у промысловых птиц и пушных зверей служит показателем тенденции изменения численности популяции.
Иногда отмечается явление “доминирования возрастного класса” часто отмечаемое у промысловых рыб. Когда в определенный период отмечается высокая выживаемость икры и личинок, значительную часть популяции составляют особи определенного возраста. В этом случае в последующие годы размножение популяции подавляется, причем механизм этого еще не вполне ясен. Пример с североморской сельдью: рыбы возрастного класса 1904 года преобладают в улове с 1910 года, когда стали достаточно крупны, до 1918 года. Такая ситуация временами порождает периодичность уловов.
Пространственная структура популяции
Распределение особей может быть: 1) случайным, 2) равномерным, 3) групповым (случайное групповое, равномерное групповое, с образованием скоплений групп).
Случайное распределение наблюдается тогда, когда среда относительно однородна, а особи не стремятся объединяться в группы, а факторы среды в пространстве изменяются случайным образом.
Равномерное – там, где между особями очень сильна конкуренция или существует антагонизм, способствующий равномерному распределению в пространстве.
Чаще всего особи стремятся образовывать группы. Пространственное распределение популяции важно учитывать при отборе проб.
Механизмы, поддерживающие определенное пространственное распределение организмов. Практически всегда в пределах ареала или других крупных площадей организмы будут распределяться пятнисто, что связано с неоднородностью ландшафтов (леса-поля, болота-сухие участки). Поэтому для эколога важно знать распределение в районе, занятом только одной популяцией.
Чаще всего встречается групповое распределение особей. Это может быть обусловлено следующими причинами:
1) векторными, определяемыми градиентами различных абиотических факторов (света, температуры, влажности, концентрации биогенов и т.п.),
2) связанными со способом размножения и расселения молодых особей (при вегетативном размножении, при отсутствии у семян средств передвижения),
3) поведенческими (образование стад, гнездовых колоний и т.д.),
4) связанными с взаимодействием разных видов (конкуренция, выедание хищниками).
Агрегация может вызвать нехватку ресурсов в определенном местообитании, но она уравновешивается повышением жизнеспособности группы, поскольку группа в отличие от отдельной особи обладает большими возможностями для своей защиты, обнаружения ресурсов или изменения микроклимата местообитания (пример – муравьи, пчелы, волки и т.п.). Степень агрегации, при которой наблюдаются оптимальный рост и выживание популяции, варьирует у разных видов и в разных условиях. Поэтому как “недонаселенность”, так и перенаселенность могут оказывать лимитирующее влияние – принцип Олли.
Особый тип агрегации назван “образованием безопасных поселений”. Здесь обширные социально организованные группы животных обосновываются в благоприятно расположенном центральном участке, откуда они регулярно расходятся для удовлетворения своих потребностей (пример – человек). Однако, чрезмерная агрегация может привести к ухудшению состояния местообитания, поэтому встает вопрос об оптимальной величине поселения.
Примеры регулярного распределения: деревья в саду или лук на грядке. В природе оно встречается реже. Так, при большой плотности растений одного вида наблюдается равномерное распределение: в густом ельнике стволы соседних деревьев удалены друг от друга на расстояние, равное сумме радиусов двух крон. В случае отдельно стоящих деревьев затенение кроной может предотвращать появление проростков особей другого вида. Обнаружено также, что некоторые растения выделяют корнями вещества, ингибирующие прорастание особей своего, а иногда и другого вида (растения пустынь).
Для некоторых животных характерно территориальное поведение: охрана отдельными особями или их небольшими группами некоторой территории от вторжения других особей своего вида. Наиболее оно выражено для хищников и растительноядных, которые специализируются на определенном виде корма.
Наблюдения за популяцией большой синицы в Оксфорде показали, что размещение ее гнезд близко к регулярному. В небольшом лесном массиве 15,5 га из 16 пар синиц изъяли 6. Уже через 3 дня на их место из зарослей придорожного кустарника проникли 4 новых пары, а кроме того увеличили размеры своей территории еще три пары. Это важно даже не только из-за необходимости сохранять кормовую базу, а для снижения вероятности стать жертвой хищников. Анализ показал, что чем меньше расстояние между гнездами, тем выше процент кладок, уничтожаемый хищниками, главным образом лаской.

Любой популяции присуща определенная организация. Распределение особей по территории, соотношение групп особей по полу, возрасту, морфологическим, физиологическим, поведенческим и генетическим особенностям отражают соответствующую структуру популяции: пространственную, половую, возрастную и т.д. Структура формируется, с одной стороны, на основе общих биологических свойств видов, а с другой – под влиянием абиотических факторов среды и популяций других видов (рис. 4).
Возрастная и половая структуры популяции. Возрастная структура популяции, т.е. соотношение в ней разных возрастных групп, определяется особенностями жизненного цикла вида и внешними условиями.
В любой популяции можно условно выделить три экологические группы: предрепродуктивную, репродуктивную, пострепродуктивную. К предрепродуктивной относится группа особей, возраст которых не достиг способности к воспроизведению; репродуктивная – группа, способная к воспроизведить новые особи; наконец, пострепродуктивная – особи, которые по ряду причин утратили способность участвовать в воспроизведении новых поколений.
Имеются виды с очень простой возрастной структурой популяций, которые состоят практически из представителей одного возраста. Так все однолетние растения весной находятся в проростках, затем почти одновременно зацветают, дают семена и к осени отмирают.
В популяции со сложной возрастной структурой представлены все возрастные группы, одновременно живут несколько поколений. Так, в стадах слонов, например, есть и новорожденные, и подростки, и молодые крепнущие животные, и способные к размножению самцы, и самки, и старые особи.
Половые группировки внутри популяций формируются на базе различной морфологии (формы и строения тела) и экологии различных полов. Отличие самцов от самок затрагивает не только строение и функцию половой системы, но и морфологию в целом (рога у самцов и отсутствие их у самок; крылатые самцы и бескрылые самки у некоторых насекомых; яркое оперение самцов и скромное у самок и т.д.).
Пространственная структура популяций. Пространство, которое занимает популяция, дает ей необходимое для жизни условия.
Пространственная структура популяции – это особенность размещения особей популяции в пространстве. Она зависит как от свойств мест обитания, так и от биологических особенностей вида. Она может изменяться во времени, зависит от сезона года, от численности популяции и т.д. Пространственное размещение особей в популяциях может быть равномерным, случайным и групповым (рис. 5).
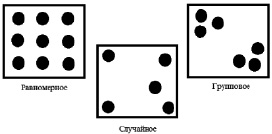
Рис. 5. Пространственное размещение особей в популяциях
При равномерном (регулярном) распределении особи размещены через более или менее равные промежутки, например, деревья в зрелом сосновом лесу. В действительности же равномерное распределение особей встречается в природе редко.
При случайном (диффузионном) распределении особи размещены неравномерно и их встречи друг с другом носят случайный характер. Такой тип распределения широко представлен среди растений и многих видов животных. Случайное расселение имеет место в однородной среде.
При групповом (мозаичном) расселении особи встречаются группами, как, например, осоковые кочки на низинном болоте, стада млекопитающих или колонии птиц. Групповое размещение обеспечивает популяции более высокую устойчивость по отношению к неблагоприятным условиям.
Распространение организмов за пределы популяции называется расселением. Способы расселения отражают, каким образом популяция занимает все большее пространство. Среди них выделяют следующие: анемохория (распространение ветром), гидрохория (водой), фитохория (растениями), зоохория (животными), антропохория (человеком). Расселение может быть пассивным (аллохория) и активным (автохория).
Передвижения животных, вызванные изменением условий существования ил связанные с циклами их развития, называют миграциями. Они могут быть регулярными (суточными и сезонными) и нерегулярными (при засухах, наводнениях, пожарах и т.д.).
Тенденция популяций поддерживать внутреннюю стабильность с помощью собственных регулирующих механизмов называется гомеостазом, а колебания численности популяций в пределах какой-то средней величины – их динамическим равновесием. Все биологические системы характеризуются способностью к гомеостазу, т.е. к саморегуляции. С помощью саморегуляции поддерживается в целом существование каждой системы – её состав и структура, внутренние связи и преобразования в пространстве и во времени.
Саморегулирование популяции осуществляется действующими в природе двумя взаимно уравновешивающимися буферными силами. С одной стороны, это биотический потенциал, составляющий совокупность всех факторов, способствующих увеличению численности популяции, а с другой – это сопротивление среды – совокупность факторов, снижающих численность популяции.
Изменения численности популяции какого-либо вида – это результат нарушения равновесия, между её биотическим потенциалом и сопротивлением окружающей среды.
Примером регулирующих факторов являются взаимоотношения организмов типа хищник – жертва, паразит – хозяин и конкуренция. Высокая численность жертвы создает условия (пищевые) для размножения хищника. Последний, в свою очередь, увеличив численность, снижает количество жертвы. Численность обоих видов в результате этого носит синхронно-колебательный характер.
Действие регулирующих факторов можно рассматривать на уровне межвидовых и внутривидовых (внутрипопуляционных) взаимоотношений организмов. К межвидовым механизмам гомеостаза относятся взаимоотношения хищник – жертва, паразит – хозяин и конкуренция. В растительном мире внутривидовая конкуренция проявляется в самоизреживании фитоценозов. В животном мире результат острой внутривидовой борьбы проявляется в форме каннибализма (поедания себе подобных). Например, явления каннибализма характерны для некоторых грызунов, личинок насекомых (майского жука), хищных рыб – щук, окуней, корюшки, трески, наваги и др.
Выделяют три типа динамики популяций: стабильный, изменчивый и взрывной (рис. 6).
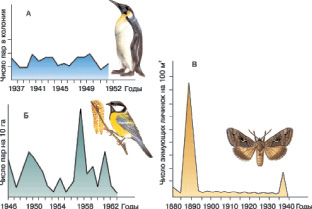
Рис. 6. Типы популяционной динамики: А – стабильный; Б – изменчивый; В – взрывной
Стабильным считается ход численности при изменениях всего в несколько раз, изменчивым – при колебаниях в десятки раз, а взрывная динамика характеризуется периодическим превышением обычной численности в сотни и тысячи раз.
Читайте также: