
Клеточный цикл это кратко
Обновлено: 04.07.2024
Описание клеточного деления базируется на данных световой микроскопии в сочетании с микрокиносъемкой и на результатах световой и электронной микроскопии фиксированных и окрашенных клеток.
1.1. Интерфаза G0
Клетки вступают в G0-фазу с контрольной точки в G1-фазе, например, точки рестрикции у животных и стартовой точки у дрожжей. Это обычно происходит в ответ на нехватку факторов роста или питательных веществ. В течение G0-фазы аппарат клеточного цикла разобран, исчезают циклины и циклин-зависимые киназы. Клетка пребывает в G0-фазе до тех пор, пока не появится повод начать деление. Клетки некоторых типов в зрелом организме, как, например, паренхимные клетки печени и почек, вступают в G0-фазу почти навсегда, и побудить их вновь начать делиться могут лишь особые обстоятельства. Другие типы клеток, как, например, эпителиальные клетки, продолжают делиться в течение всей жизни организма и редко входят в G0-фазу.
Хотя многие клетки в G0-фазе могут умереть вместе со всем организмом, не все клетки, вступающие в G0-фазу, обречены в скором времени умереть. Это часто является результатом отсутствия стимула для клетки вновь вступить в клеточный цикл.
Старение клеток отличается от состояния покоя тем, что старение клеток — это состояние, вызванное повреждением ДНК или деградацией, делающей размножение клетки невозможным. Кроме того, старение, в отличие от покоя, часто служит биохимической альтернативой саморазрушению — апоптозу для сильно повреждённых клеток. Наконец, покой — обратимое состояние, а старение — нет
1.2. Митоз
Продолжительность митоза в среднем составляет 1—2 часа[1][4]. Митоз клеток животных, как правило, длится 30—60 минут, а растений — 2—3 часа.[5] За 70 лет в теле человека суммарно осуществляется порядка 1014 клеточных делений[6].
Аппарат клеточного деления
Деление всех эукариотических клеток сопряжено с формированием специального аппарата клеточного деления. Активная роль в митотическом делении клеток зачастую отведена цитоскелетным структурам. Универсальным как для животных, так и для растительных клеток является двухполюсное митотическое веретено, состоящее из микротрубочек и связанных с ними белков. Веретено деления обеспечивает строго одинаковое распределение хромосом между полюсами деления, в области которых в телофазе образуются ядра дочерних клеток.
Ещё одна не менее важная структура цитоскелета отвечает за разделение цитоплазмы (цитокинез) и, как следствие, за распределение клеточных органелл. В животных клетках за цитокинез отвечает сократимое кольцо из актиновых и миозиновых филаментов. В большинстве клеток высших растений из-за наличия жёсткой клеточной стенки цитокинез протекает с образованием клеточной пластинки в плоскости между двумя дочерними клетками. При этом область образования новой клеточной перегородки определяется заранее предпрофазным пояском из актиновых микрофиламентов, а поскольку актин участвует также в формировании клеточных септ у грибов, возможно, что он направляет цитокинез у всех эукариот [19] .
Веретено деления
Поздняя метафаза митоза в клетке лёгкого тритона (использованы иммунофлуоресцентные красители). Четко просматривается веретено деления, образованное микротрубочками (зелёные), и хромосомы (синие)
По морфологии различают два типа митотического веретена: астральный (или конвергентный) и анастральный (дивергентный) [~ 1][24] .
Астральный тип митотической фигуры, характерный для животных клеток, отличают благодаря небольшим зонам, на полюсах веретена, в которых сходятся (конвергируют) микротрубочки. Зачастую центросомы, располагающиеся в области полюсов астрального веретена, содержат центриоли. От полюсов деления также расходятся во всех направлениях радиальные микротрубочки, не входящие в состав веретена, а образующие звездчатые зоны — цитастеры.
Анастральный тип митотической фигуры отличается широкими полярными областями веретена, так называемыми полярными шапочками, в их состав не входят центриоли. Микротрубочки при этом расходятся широким фронтом (дивергируют) от всей зоны полярных шапочек. Этот тип митотической фигуры также отличает отсутствие цитастеров. Анастральный тип митотического веретена наиболее характерен для делящихся клеток высших растений, хотя иногда наблюдается и в некоторых клетках животных.
Микротрубочки
Микротрубочки — динамичные структуры, принимающие активное участие в построении веретена деления во время митоза. Химически они представляют собой биополимеры, состоящие из субъединиц белка тубулина. Количество микротрубочек в клетках различных организмов может значительно отличаться. В метафазе веретено деления в клетках высших животных и растений может содержать до нескольких тысяч микротрубочек, тогда как у некоторых грибов их всего около 40 [22] .
Центромеры и кинетохоры
Основные статьи: Центромера, Кинетохор
Центромеры — специализированные последовательности ДНК, необходимые для связывания с микротрубочками веретена деления и для последующего расхождения хромосом. В зависимости от локализации различают несколько типов центромер. Для голоцентрических центромер характерно образование связей с микротрубочками веретена по всей длине хромосомы (некоторые насекомые, нематоды, некоторые растения). В противоположность голоцентрическим моноцентрические центромеры служат для связи с микротрубочками в единственной области хромосомы [24] .
В центромерной области обычно располагаются кинетохоры хромосом — сложные белковые комплексы, морфологически очень сходные по своей структуре для различных групп эукариот, как, например, для диатомовых водорослей, так и для человека [25] . Обычно на каждую хроматиду (хромосому) приходится по одному кинетохору. На электронных микрофотографиях кинетохор обычно выглядит как пластинчатая трёхслойная структура [26] . Порядок слоев следующий: внутренний плотный слой, примыкающий к телу хромосомы; средний рыхлый слой; внешний плотный слой, от которого отходит множество фибрилл, образуя т.н. фиброзную корону кинетохора.
К основным функциям кинетохора относят: закрепление микротрубочек веретена деления, обеспечение движения хромосом во время митоза при участии микротрубочек, связывание между собой сестринских хроматид и регуляцию их последующего разделения в анафазе митоза [27] . Минимально достаточно одной микротрубочки (например, для дрожжей) ассоциированной с кинетохором, чтобы обеспечить движение хромосомы. Однако с одним кинетохором могут быть связаны целые пучки, состоящие из 20—40 микротрубочек (например, у высших растений или человека), чтобы обеспечить расхождение хромосом к полюсам клетки [26][27] .
1.3. Циклины и циклин-зависимые киназы
Циклины — семейство белков-активаторов циклин-зависимых протеинкиназ (CDK) (англ. CDK, cyclin-dependent kinases) — ключевых ферментов, участвующих в регуляции клеточного цикла эукариот. Циклины получили своё название в связи с тем, что их внутриклеточная концентрация периодически изменяется по мере прохождения клеток через клеточный цикл, достигая максимума на его определенных стадиях.
Каталитическая субъединица циклин-зависимой протеинкиназы частично активируется в результате взаимодействия с молекулой циклина, которая образует регуляторную субъединицу фермента. Образование этого гетеродимера становится возможным после достижения циклином критической концентрации. В ответ на уменьшение концентрации циклина происходит инактивация фермента. Для полной активации циклин-зависимой протеинкиназы должно произойти специфическое фосфорилирование и дефосфорилирование определенных аминокислотных остатков в полипептидных цепях этого комплекса. Одним из ферментов, осуществляющих подобные реакции, является киназа CAK (CAK — CDK activating kinase).
Рис. Концентрация циклинов в клеточном цикле
| циклин | Cdk | фаза клеточного цикла |
|---|---|---|
| циклин Е | Cdk2 | вход в S-фазу |
| циклин A | Cdk2 | вход в G2-фазу |
| циклин B | Cdk1 | вход в M-фазу |
2. Регуляция клеточного цикла
Закономерная последовательность смены периодов клеточного цикла осуществляется при взаимодействии таких белков, как циклин-зависимые киназы и циклины. Клетки, находящиеся в G0 фазе, могут вступать в клеточный цикл при действии на них факторов роста. Разные факторы роста, такие как тромбоцитарный, эпидермальный, фактор роста нервов, связываясь со своими рецепторами, запускают внутриклеточный сигнальный каскад, приводящий в итоге к транскрипции генов циклинов и циклин-зависимых киназ. Циклин-зависимые киназы становятся активными лишь при взаимодействии с соответствующими циклинами. Содержание различных циклинов в клетке меняется на протяжении всего клеточного цикла. Циклин является регуляторной компонентой комплекса циклин-циклин-зависимая киназа. Киназа же является каталитическим компонентом этого комплекса. Киназы не активны без циклинов. На разных стадиях клеточного цикла синтезируются разные циклины. Так, содержание циклина B в ооцитах лягушки достигает максимума к моменту митоза, когда запускается весь каскад реакций фосфорилирования, катализируемых комплексом циклин-В/циклин-зависимая киназа. К окончанию митоза циклин быстро разрушается протеиназами.

2.1. Контрольные точки клеточного цикла
3. Ингибиторы циклин-зависимой киназы
Ингиби́тор цикли́н-зави́симой кина́зы (англ. Cdk inhibitor protein, CKI, CDI, CDKI) — белок, блокирующий активность циклин-зависимой киназы отдельно или циклин-зависимой киназы в комплексе с циклином. Обычно сдерживающая активность CKI приурочена к фазе G1 клеточного цикла. К тому же, активация CKI может происходить в ответ на провреждения ДНК или может быть вызвана внеклеточными ингибирующими сигналами[2].
Большинство эукариотических организмов обладают ингибиторами циклин-зависимых киназ. В животных клетках выделяют два семейства CKI: Cip/Kip и INK4.
Ингибиторы семейства Cip/Kip блокируют циклин-зависимую киназу в комплексе с циклином, а ингибиторы семейства INK4 блокируют отдельные циклин-зависимые киназы Cdk4 и Cdk6. В животных клетках ингибиторы циклин-зависимых киназ разделяются на два основных семейства: Cip/Kip и INK4. Семейство Cip/Kip включает ингибиторы CDK белки p21, p27, p57. К основным субстратам Cip/Kip-ингибиторов относятся циклин-киназные комплексы G1/S-Cdk и S-Cdk, отвечающие, соответственно, за G1/S-переход и вступление в S-фазу. Ингибиторы семейства INK4 блокируют циклин-зависимые киназы Cdk4 и Cdk6 регулирующие G1-фазу клеточного цикла.
| Рис. Схема ингибирования Cdk6 с участием INK4. Белок INK4 соединяется с циклин-зависимой киназой Cdk6 и смещает аминоконцевую долю киназы примерно на 15° относительно оси вращения. В итоге, деформируется каталитическая область Cdk6, а также снижается способность циклин-зависимой киназы к связыванию циклина. |
На протяжении фазы G1 в растущей клетке блокируется активность циклин-зависимых киназ (англ. Cdk) до момента вступления клетки в очередной клеточный цикл. Сдерживание активности Cdk обеспечивается тремя контрольными механизмами. Во-первых, снижением экспрессии генов циклинов. Во-вторых, увеличением степени деградации циклинов. Наконец, к третьему типу сдерживания активности Cdk относятся ингибиторы CKI. Помимо обеспечения стабильного роста клетки в фазе G1 ингибиторы циклин-зависимых киназ участвуют в аресте клеточного цикла на стадии G1 в ответ на неблагоприятные внешние условия. К тому же события клеточного цикла могут блокироваться с участием CKI при повреждениях ДНК[2].
Ингибиторы циклин-зависимых киназ: Sic1 у почкующихся дрожжей, Rum1 у делящихся дрожжей и Rux у Drosophila — несмотря на структурные различия обладают как минимум тремя сходными функциональными особенностями. Во-первых, основными мишенями данных CKI являются митотические циклин-киназы (англ. M-Cdk) и циклин-киназы синтетической фазы клеточного цикла (англ. S-Cdk). В то же время указанные ингибиторы CKI не могут блокировать циклин-зависимые киназы, обеспечивающие переход клетки из фазы G1 в S-фазу (англ. G1/S-Cdk). Наконец, третьей характерной особенностью всех перечисленных ингибиторов CKI является способ их деактивации. Все они разрушаются после фосфорилирования со стороны активных циклин-зависимых киназ[2].
Семейство Cip / Kip (p21, p27, p57) регулирует динамику актина посредством ингибирования пути Rho-ROCK-LIMK [2]

3.1. CDKN1A или P21
CDKN1A (англ. cyclin-dependent kinase inhibitor 1A, p21, Cip1) — внутриклеточный белок-ингибитор циклин-зависимой киназы 1A, играет критическую роль в клеточном ответе на повреждение ДНК. Уровень белка повышен в клетках, находящихся в стадии покоя, таких как дифференцированные клетки организма. Один из 9 известных белковых ингибиторов циклин-зависимой киназы.
p21 обеспечивает устойчивость гематопоэтических клеток к инфицированию ВИЧ за счёт связывания с вирусной интегразой, предотвращая таким образом встраивание провируса в хромосомный аппарат клетки.

Ген CDKN1A как правило не инактивируется полностью в злокачественных опухолях. Точная роль p21 в канцерогенезе до конца пока не установлена. Исследования показывают, что при некоторых типах опухолей потеря p21 является признаком плохих шансов на выживание. Однако известны ситуации, когда повышенная концентрация этого белка в клетках положительно коррелирует с агрессивностью опухоли и её способностью к метастазированию. Это особенно относится к тем случаям, когда p21 накапливается в цитоплазме, а не в ядре клетки
Ген р21 и регенерация.
Исследовательская группа профессора Эллен Хэбер-Катц (Ellen Heber-Katz) выяснила, что выключение гена р21 активирует у мышей способность к восстановлению тканей.
В природе регенеративные способности (восстановление тканей, органов и конечностей) наиболее выражены у амфибий, полипов, иглокожих, губок, гидр, дождевых червей, саламандр и нек. др., в то время как у млекопитающих они существенно ограничены.
В лаборатории молекулярного биолога Хэбер-Катц регенерацией у млекопитающих заинтересовались в 1996 году, когда во время изучения одного из аутоиммунных заболеваний зафиксировали у лабораторных мышей линииMurphy Roths Large (MRL) восстановление хрящевой ткани ушей после пожизненной маркировки (перфорации). У животных других линий такой регенерации не происходило. Ученые исследовали восстановленные участки и зафиксировали в них синтез ДНК, пролиферацию клеток и появление новых волосяных фолликулов.
Последующие эксперименты на мышах MRL показали, что после нанесенных им повреждений в области сердечной мышцы и спинного мозга, эти ткани так же успешно регенеририруют без образования рубцов.
Чтобы с наибольшей вероятностью вычислить гены, ответственные за процессы регенерации у мышей MRL, было решено картировать их геном. Так же под прицел ученых попал феномен влияния изменений в ДНК на цикл деления клеток.
Исследования показали, что у животных MRL линии неактивен ген р21, играющий важную роль в регуляции клеточного деления и в торможении процесса деления клеток при повреждении ДНК.
В новой работе ученые показали, что мыши без гена p21 демонстрируют регенеративные характеристики, наблюдаемые у мышей MRL линии. У генноинженерных мышей фиксировалось быстрое деление клеток, восстанавливающее ткани и быстрый апоптоз - механизм клеточного самоуничтожения, включающийся, если ДНК повреждена. Сочетание этих процессов свойственно видам, обладающим регенеративными способностями. Ученые считают, что такое совокупное воздействие позволяет клеткам данных видов делиться быстро, не выходя из-под антиракового контроля.
В научной публикации, опубликованной в Трудах Национальной академии наук, коллектив Эллен Хэбер-Катц предоставляет веские доказательства, что тканевая регенерация связана с контролем клеточного деления.
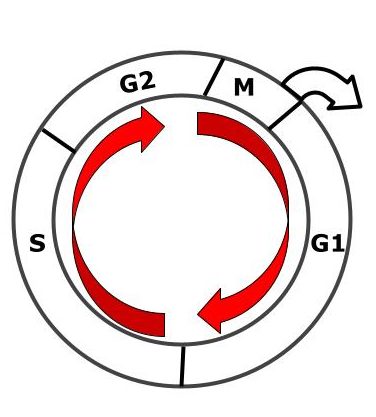
Сначала я хотел написать статью про митоз: какие клетки в него вступают, через какие фазы он проходит и его биологическое значение. Но передумал. Почему? Митоз или непрямое деление клетки — это только часть, а иногда и небольшая, более общего понятия. Я говорю про клеточный цикл. Поэтому сначала давайте разберемся с ним, а дальше уже перейдем к митозу. В самом конце статьи проведем сравнение между митозом и мейозом.
Клеточный цикл
Клеточный цикл — это время от образования клетки до ее деления или гибели. Состоит из двух периодов — интерфазы и митоза. Интерфаза в свою очередь состоит еще из трех периодов: G1 — пресинтетический, S — синтетический и G2 — постсинтетический. На самом деле, такие периоды клеточного цикла только у одного типа клеток, но давайте сначала разберем классику. Последовательность периодов интерфазы:
Рисунок. Клеточный цикл.
Пресинтетический или постмитотический период — G1
Клетка только образовалась путем митоза, а значит ей нужно немного подрасти — восстановить свои органеллы (рибосомы и митохондрии), синтезировать белок и РНК. Митоз все-таки был затратным, часть белков была потрачена на образование новой клетки. Хромосомный набор 2n2c, подробнее о хромосомном наборе здесь. Длительность G1 периода интерфазы — 9 часов.
Синтетический период или S-период
В этом периоде клетка удваивает количество молекул ДНК и синтезирует гистоны — белки, которые упаковывают новообразованную молекулу ДНК. Но зачем ее упаковывать? Потому что они огромные. 46 молекул ДНК в одной клетке — это примерно два метра, а тут молекул 92, то есть в одной клетке целых четыре метра ДНК. Шок контент. Но я отвлекся, так вот — молекула ДНК упаковалась и стала хроматидой. По сути, это почти готовая хромосома, но она связана со второй хроматидой с помощью центромеры. Поэтому хромосомный набор — 2n4c. Длительность S-периода интерфазы — 10 часов.

Рисунок. Синтетический период интерфазы.
Постсинтетический период — G2
Клетка синтезирует тубулин. Запасает энергетические субстраты — АТФ. Созревают центриоли(они тоже удваиваются), вместе с тубулином они образуют веретено деления. Подготовка к делению идет полным ходом.
Дальше клетка вступает в митоз и мы его рассмотрим ниже, но перед этим… Нам нужно серьезно поговорить. Такая схема клеточного цикла подходит не всем клеткам, а только одному виду. Есть идеи почему? Да, правильно! У нас в организме множество клеток, очень разных по структуре и функциям. Какие-то делятся постоянно (стволовые клетки), а другие не делятся вообще (эритроцит — у него ведь даже ядра нет, какое там деление?). Значит у разных клеток — разные клеточные циклы. Дальше мы увидим, что такой рисунок подходит только одному типу клеток. Набор без изменений — 2n4c. Длительность G2— периода интерфазы — 4,5 часа.
Типы клеточных циклов
Всего есть три варианта клеточных циклов, но у каждого есть свои особенности.
Митотический цикл
Здесь все просто, у клетки только одна задача — делиться. Зачем ей постоянно делиться? Затем, что ее последователи мрут как мухи.
- Гемопоэтические — нужно постоянно поддерживать количество эритроцитов, тромбоцитов и лейкоцитов в организме.
- Клетки базального слоя эпидермиса.
- Эпителиальные клетки кишечника.
- Ещё так делятся сперматогонии и оогонии — они образуют множество диплоидных клеток, которые потом вступят в мейоз.
Для таких клеток подходит круг и все его периоды.
Клеточный цикл у условно постмитотических клеток
Клетка образовалась, восстановила свои органоиды, синтезировала белки и РНК — G1 период. Дальше она дифференцируется и начинает выполнять свои функции. Клетка сохранила способность к делению, но есть один нюанс. Клетка может вступить в митотический цикл при определенном условии — чаще всего при повреждении. Эти клетки обеспечивают регенерацию тканей и органов в случае их травмы. Такие клетки:
- Фибробласты
- Гепатоциты
- Лимфоциты
- Стволовые клетки мышечной ткани и костной — эти просто “покоятся” и активируются при повреждении.
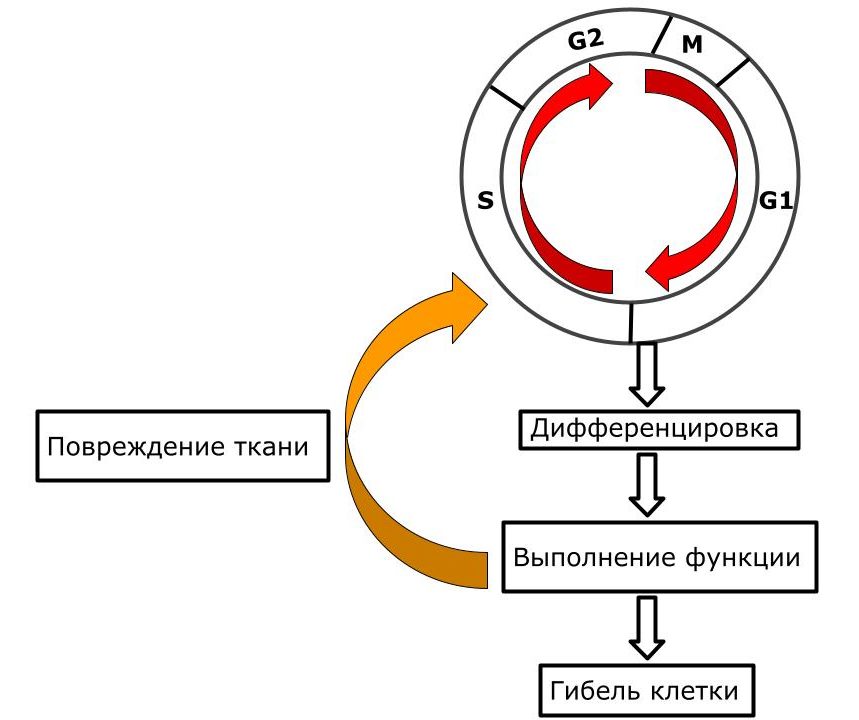
Схема. Клеточный цикл условно постмитотических клеток.
Клеточный цикл у постмитотических клеток
Клетка проходит через G1 период и дифференцировку. Но такие клетки не могут делиться. Но почему? Тут несколько вариантов, но все они связанны с функцией клетки.
Первый вариант — клетка настолько преисполнилась в своем познании, что ей уже это не интересно. Шуточка, на самом деле эти клетки выполняют такие сложные функции, что им не до деления — нервные клетки, клетки сердечной мышцы и обычных мышц (правда тут не совсем клетки, а симпласты — клетки объединились и если мы на них посмотрим, то увидим одну цитоплазму и кучу ядер).
Второй вариант — чтобы клетка могла нормально выполнять свои функции ей пришлось чем-то пожертвовать. Эритроциту нужно переносить как можно больше кислорода, поэтому он избавился от ядра. Клетки всех слоев кожи, кроме базального — постепенно наполняются кератином и погибают, а все для защиты внутренней среды организма.
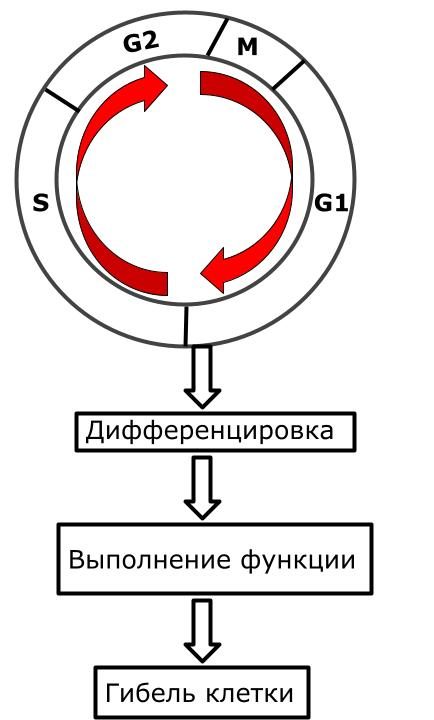
Схема. Клеточный цикл постмитотических клеток.
Вроде все, с клеточными циклами разобрались. Теперь можно переходить к митозу.
Митоз или непрямое деление клетки
В ходе митоза из одной материнской клетки образуется две генетически идентичные дочерние клетки. Можно даже сказать, что клетка образует своего клона. Хромосомный набор клетки остается без изменений: как был 2n2c, так и остается. Перед митозом произошла интерфаза и количество молекул ДНК удвоилось, мы видели это выше. Получается, что клетка готова к делению и входит в свою первую фазу.
Профаза митоза
Начинается с упаковки хромосом (конденсации). Зачем это нужно клетке? Так она пытается сохранить свой генетический материал и передать его в целостности и сохранности. Такая упаковка снижает риск его потери. Помните, что в микроскоп нельзя увидеть хромосомы? Так вот, здесь клетка постаралась и упаковала из настолько, что они видны. Каждая хромосома состоит из двух хроматид (одна хроматида — одна молекула ДНК). Из-за упаковки ДНК исчезают ядрышки и прекращается синтез РНК. Ядерная оболочка, эндоплазматическая система и аппарат Гольджи распадаются. Происходит формирование веретена деления — за счет удвоенных центриолей и тубулина.
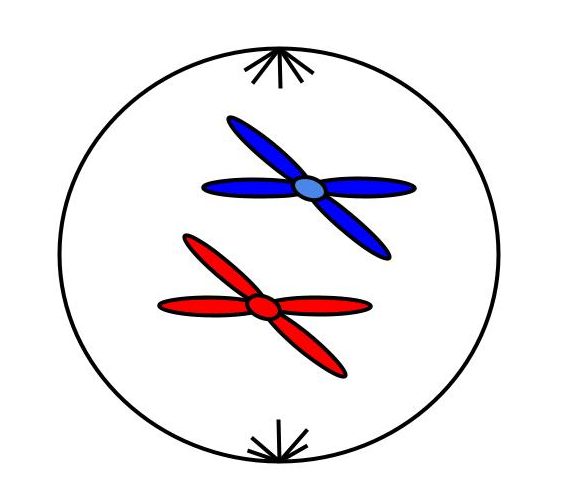
Схема. Профаза митоза
2n4c
Хромосомный набор в профазу митоза
Метафаза митоза
Заканчивается образование веретена деления, конденсация хромосом максимальна. Нити веретена деления прикрепляются к центромерам — хромосомы располагаются на экваторе клетки.
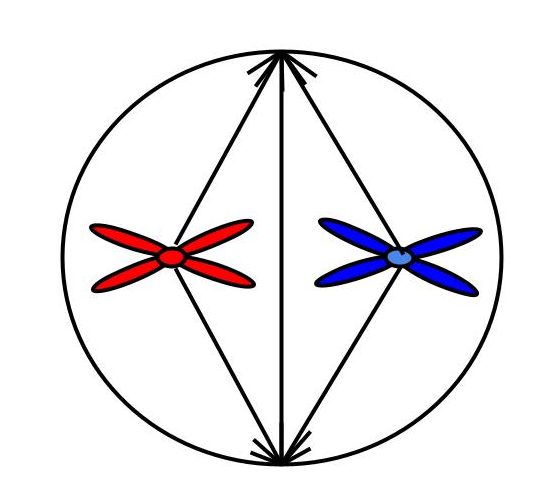
Схема. Метафаза митоза
2n4c
Хромосомный набор в метафазу митоза
Анафаза митоза
Нити веретена деления тянут хроматиды к полюсам клетки, таким образом из одной двухроматидной хромосомы образуется две хромосомы. И всего в клетке — четыре хромосомы.
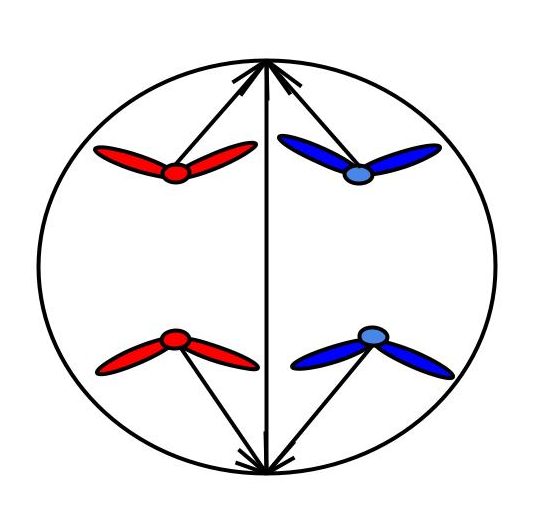
Схема. Анафаза митоза
У полюсов 2n2c, в клетке 4n4c
Хромосомный набор в анафазу
Телофаза митоза
Хромосомы постепенно раскручиваются, клетка прибыла в место назначения — теперь можно распаковать чемоданы. Вокруг хромосом образуется ядерная оболочка, а затем и клеточная перетяжка. Органеллы распределяются между двумя дочерними клетками. Вот и всё, митоз завершен.
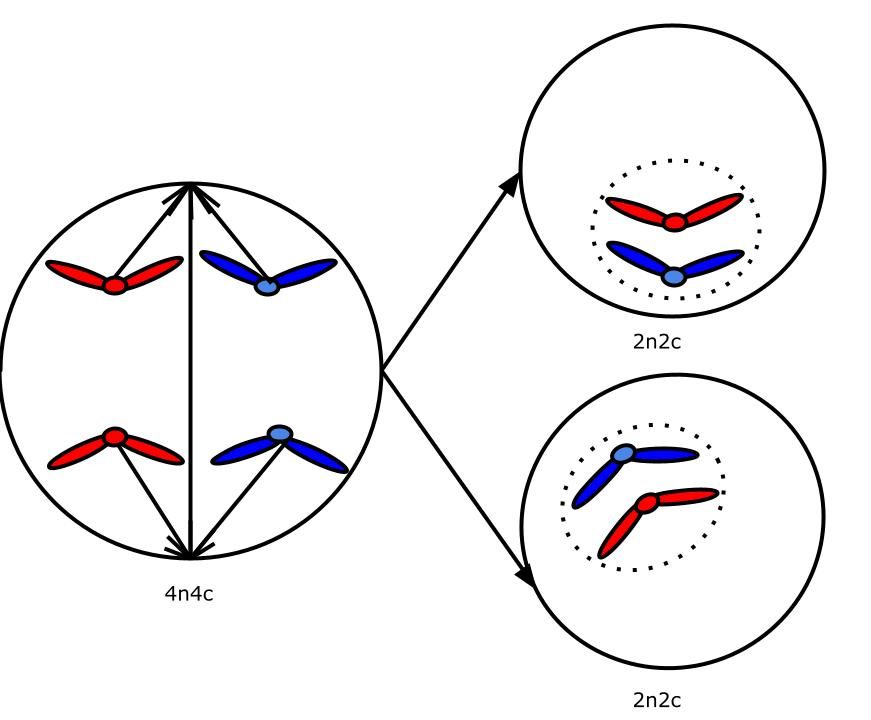
Схема. Телофаза митоза
Две диплоидных клетки — 2n2c
Результат митоза
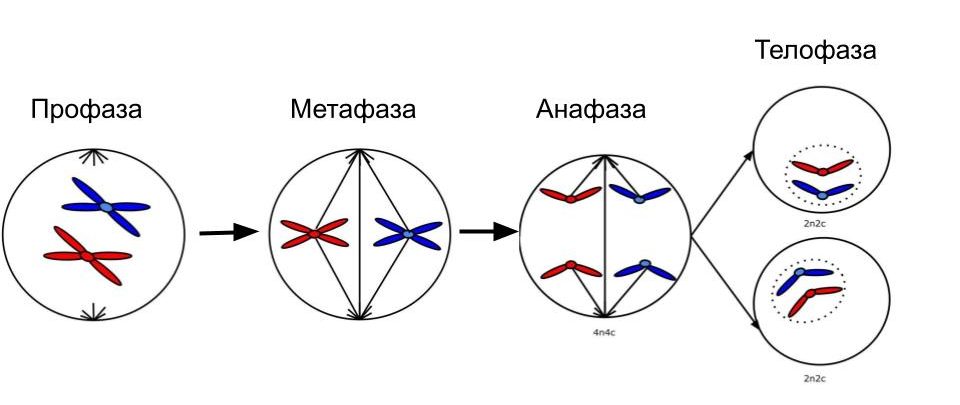
Общая схема митоза
Длительность митоза — полчаса. Следовательно, длительность митотического цикла = G1 + S + G2 + митоз = 9 + 10 + 4,5 + 0,5 = 24 часа.
Биологическое значение митоза
- Поддержание постоянного набора хромосом в клетке.
- Получение идентичных клеток в ходе деления.
- Митоз обеспечивает рост и регенерацию тканей.
Сходства и различия между мейозом и митозом
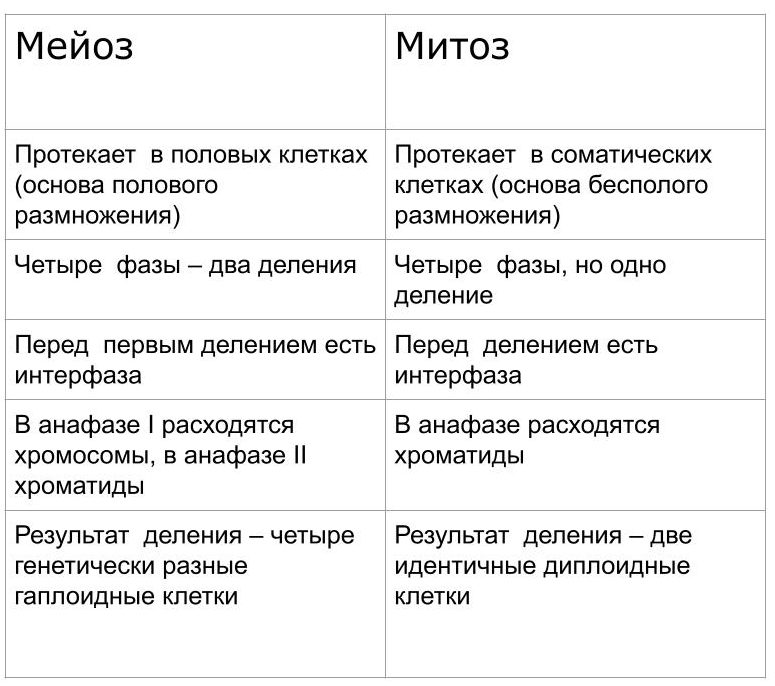
Таблица. Сравнительная характеристика мейоза и митоза
Введение
Для того чтобы клетка смогла полноценно разделиться, она должна увеличиться в размерах и создать достаточное количество органоидов. А для того чтобы не растерять наследственную информацию при делении пополам, она должна изготовить копии своих хромосом. И, наконец, для того чтобы распределить наследственную информацию строго поровну между двумя дочерними клеткам, она должна в правильном порядке расположить хромосомы перед их распределением по дочерним клеткам. Все эти важные задачи решаются в процессе клеточного цикла.
Клеточный цикл имеет важное значение, т.к. он демонстрирует важнейшие свойства клетки: способность к размножению, росту и дифференцировке. Обмен тоже идёт, но его не рассматривают при изучении клеточного цикла.
Определение понятия
Клеточный цикл (синоним: жизненный цикл клетки) - это период жизни клетки от её рождения до образования дочерних клеток или смерти.
У животных клеток клеточный цикл, как промежуток времени между двумя делениями (митозами), длится в среднем от 10 до 24 часов.
Клеточный цикл состоит из нескольких периодов (синоним: фазы), которые закономерно сменяют друг друга. В совокупности первые фазы клеточного цикла (G1, G0, S и G2) носят название интерфазы, а последняя фаза называется митозом.
Рис. 1. Клеточный цикл.
Периоды (фазы) клеточного цикла
1. Период первого роста G1 (от английского Growth - рост), составляет 30-40% цикла, и период покоя G0
Синонимы: постмитотический (наступает после митоза) период, пресинтетический (проходит перед синтезом ДНК) период.
Клеточный цикл начинается от рождения клетки в результате митоза. Дочерние клетки, появившиеся в результате деления материнской клетки, уменьшены в размерах и в них меньше органоидов, чем в норме. Поэтому "новорожденная" маленькая клетка в первом периоде (фазе) клеточного цкла (G1) растёт и увеличивается в размерах, а также формирует недостающие органоиды. Идёт активный синтез белков, необходимых для ввсего этого. В результате клетка становится полноценной, можно сказать, "взрослой".
Чем обычно заканчивается для клетки период роста G1?
- Вступллением клетки в процесс дифференцировки. За счёт дифференцировки клетка приобретает специальные особенности для выполнения функций, необходимых всему органу и организму. Запускается дифференцировка управляющими веществами (гормонами), воздействующими на соответствующие молекулярные рецепторы клетки. Клетка, завершившая свою дифференцировку, выпадает из круговорота делений и находится в периоде покоя G0. Требуется воздействие активирующих веществ (митогенов) для того, чтобы она претерпела дедифференцировку и вновь вернулась в клеточный цикл.
- Гибелью (смертью) клетки.
- Вступлением в следующий период клеточного цикла -синтетический.
2. Синтетический период S (от английского Synthesis - синтез), составляет 30-50% цикла
Понятие синтеза в названии этого периода относится к синтезу (репликации) ДНК, а не к каким-либо другим процессам синтеза. Достигнув определенного размера в результате прохождения периода первого роста, клетка вступает в синтетический период, или фазу, S, в котором происходит синтез ДНК. За счёт репликации ДНК клетка удваивает свой генетический материал (хромосомы), т.к. в ядре образуется точная копия каждой хромосомы. Каждая хроммосома становится двойной и весь хромосомный набор становится двойным, или диплоидным. В результате клетка теперь готова поделить наследственный материал поровну между двумя дочерними клетками, не потеряв при этом ни одного гена.
3. Период второго роста G2 (от английского Growth - рост), составляет 10-20% цикла
Синонимы: премитотический (проходит перед митозом) период, постсинтетический (наступает после синтетического) период.
Период G2 является подготовительным к очередному делению клетки. Во время второго периода роста G2 клетка производит белки, требующиеся для митоза, в частности, тубулин для веретена деления; создаёт запас энергии в виде АТФ; проверяет, закончена ли репликация ДНК, и готовится к делению.
4. Период митотического деления M (от английского Mitosis - митоз), составляет 5-10% цикла
Митоз M (синоним: митотический цикл), заключается в том, что клетка правильно делится на две дочерние клетки. Благодаря механизмам комплементарного синтеза при репликации ДНК в синтетическом периоде и механизму распределения хроматид в митозе каждая дочерняя клетка получает идентичный набор хромосом, являющийся точной копией хромосомного набора материнской клетки. Короче говоря, за счёт танцевв хромосом, они расределяются пополам и поровну между двумя дочерними клетками, образовавшимися в результате деления. Подробнее о митозе.
После деления клетка оказывается в новой фазе G1, и клеточный цикл завершается.


Регуляция клеточного цикла
На молекулярном уровне переход от одной фазы цикла к другой регулируют два белка - циклин и циклинзависимая киназа (CDK).
Для регуляции клеточного цикла используется процесс обратимого фосфорилирования/дефосфорилирования регуляторных белков, т.е. присоединение к ним фосфатов с последующим отщеплением. Ключевым веществом, регулирующим вступление клетки в митоз (т.е. её переход от фазы G2 к фазе M), является специфическая серин/треонин-протеинкиназа, которая носит название фактор созревания - ФС, или MPF, от английского maturation promoting factor. В активной форме этот белковый фермент катализирует фосфорилирование многих белков, принимающих участие в митозе. Это, например, входящий в состав хроматина гистон H1, ламин (компонент цитоскелета, находящийся в ядерной мембране), факторы транскрипции, белки митотического веретена, а также ряд ферментов. Фосфорилирование этих белков фактором созревания MPF активирует их и запускает процесс митоза. После завершения митоза регуляторная субъединица ФС, циклин, маркируется убиквитином и подвергается распаду (протеолизу). Теперь наступает очередь протеинфосфатаз, которые дефосфорилируют белки, принимавшие участие в митозе, чем переводят их в неактивное состояние. В итоге клетка возвращается в состояние интерфазы.
ФС (MPF) — это гетеродимерный фермент, включающий в себя регуляторную субъединицу, а именно циклин, и каталитическую субъединицу, а именно циклинзависимую киназу ЦЗК (CDK от англ. cyclin dependent kinase), она же p34cdc2; 34 кДа. Активной формой этого фермента является лишь димер ЦЗК+циклин. Кроме того, активность ЦЗК регулируется путем обратимого фосфорилирования самого фермента. Циклины получили такое название потому, что их концентрация циклически изменяется в соответствии с периодами клеточного цикла, в частности, она снижается перед началом деления клетки.
В клетках позвоночных присутствует ряд различных циклинов и циклинзависимых киназ. Разнообразные сочетания двух субъединиц фермента регулируют запуск митоза, начало процесса транскрипции в G1-фазе, переход критической точки после завершения транскрипции, начало процесса репликации ДНК в S-периоде интерфазы (стартовый переход) и другие ключевые переходы клеточного цикла (на схеме не приведены).
В ооцитах лягушки вступление в митоз (G2/M-переход) регулируется путем изменения концентрации циклина. Циклин непрерывно синтезируется в интерфазе до достижения максимальной концентрации в фазе М, когда запускается весь каскад фосфорилирования белков, катализируемый ФС. К окончанию митоза циклин быстро разрушается протеиназами, также активируемыми ФС. В других клеточных системах активность ФС регулируется за счет различной степени фосфорилирования самого фермента.

Клеточный цикл представляет собой сложную последовательность событий предшествующих делению или гибели клетки. В эукариотических клетках этот процесс включает в себя четыре отдельные фазы: фаза Gap 1 (G1); фаза синтеза (S); фаза Gap 2 (G2); митоз (M).
Фазы G1, S и G2 клеточного цикла вместе называются интерфазой. Делящаяся клетка проводит большую часть своего времени именно в интерфазе, поскольку при подготовке к делению она растет. Фаза митоза связана с разделением ядерных хромосом с последующим цитокинезом (разделение цитоплазмы на две отдельные клетки). В конце митотического цикла образуются две разные дочерние клетки. Каждая клетка содержит идентичный генетический материал.
Время, необходимое для завершения деления клетки зависит от ее типа. К примеру, клетки крови в костном мозге, клетки кожи, клетки желудка и кишечника, делятся быстро и постоянно. Другие клетки делятся при необходимости, заменяя поврежденные или мертвые клетки. К таким типам клеток относятся клетки почек, печени и легких. Другие типы клеток, в том числе нервные клетки, прекращают деление после созревания.
Периоды и фазы клеточного цикла
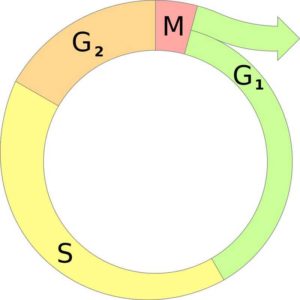
Два основные периода клеточного цикла эукариот включат интерфазу и митоз:
Интерфаза
Во время этого периода, клетка удваивает свою цитоплазму и синтезирует ДНК. По оценкам, делящаяся клетка тратит около 90-95% своего времени на интерфазу, которая состоит из следующих 3-х фаз:
- Фаза G1: промежуток времени до синтеза ДНК. В этой фазе клетка увеличивает свои размеры и количество органелл, подготавливаясь к делению. Клетки животных в этой фазе диплоидны, что означает наличие двух наборов хромосом.
- S-фаза: этап цикла, в течение которого синтезируется ДНК. В большинстве клеток имеется узкое временное окно, в течение которого происходит синтез ДНК. Содержание хромосом в этой фазе удваивается.
- Фаза G2: период после синтеза ДНК, но до начала митоза. Клетка синтезирует дополнительные белки и продолжает увеличиваться в размерах.
Фазы митоза
Во время митоза и цитокинеза содержимое материнское клетки равномерно распределяется между двумя дочерними клетками. Митоз имеет пять фаз: профаза, прометафаза, метафаза, анафаза и телофаза.
Цитокинез – процесс разделение цитоплазмы в эукариотических клетках, которые продуцируют различные дочерние клетки. Цитокинез возникает в конце клеточного цикла после митоза или мейоза.
При делении клеток животных цитокинез возникает, когда сократительное кольцо образует расщепленную борозду, которая зажимает клеточную мембрану пополам. В растительных клетках строится клеточная пластинка, которая делит клетку на две части.
Как только клетка завершит все фазы клеточного цикла, она возвращается в фазу G1 и весь цикл повторяется снова. Клетки организма также способны находится в состояние покоя, которое называется фазой Gap 0 (G0) в любой момент своего жизненного цикла. Они могут оставаться на этой стадии в течение очень длительного периода времени, пока не поступят сигналы к прохождению через клеточный цикл.
Клетки, которые содержат генетические мутации, постоянно помещаются в фазу G0, чтобы препятствовать их реплицированию. Когда клеточный цикл идет не так, как надо, нарушается нормальный рост клеток. Могут развиться раковые клетки, которые получают контроль над своими собственными сигналами роста и продолжают размножаться беспрепятственно.
Клеточный цикл и мейоз
Не все клетки делятся через процесс митоза. Организмы, которые размножаются половым путем, также подвергаются типу клеточного деления, называемого мейозом. Мейоз возникает в половых клетках и аналогичен процессу митоза. Однако после полного клеточного цикла в мейозе образуются четыре дочерние клетки. Каждая клетка содержит половину числа хромосом исходной (родительской) клетки. Это означает, что половые клетки являются гаплоидными клетками. Когда гаплоидные мужские и женские половые клетки объединяются в процессе, называемом оплодотворением, они образуют одну диплоидную клетку, называемую зиготой.
Читайте также: