
История открытия рнк кратко
Обновлено: 23.06.2024
Нуклеиновые кислоты (НК) входят в состав клеток всех живых организмов. Они играют главную роль в хранении, передаче наследственной информации и ее реализации в процесса синтеза белков.
НК были обнаружены были обнаружены в XIX веке щвейцарским врачом Иоганом Фридрихом Мишером.
В 1909 г. Фибус Левин предположил, что НК состоят из мономеров – нуклеотидов. А в 1930 г. установил различия нуклеотидов по типу углеводной компоненты. НК, содержащие рибозу – РНК, дезоксирибозу – ДНК.
В 1934 г. Торбьёрн Оскар Касперссон вместе с Эйнаром Хаммерстеном показали, что НК — это гетерополимеры, мономерами которых являются нуклеотидные звенья.
Одновременно с изучением структуры нуклеиновых кислот проводились исследования, направленные на установление их биологической роли.
В 1881 г. Эдуард Захариас доказал, что нуклеин содержится в хромосомах, которые уже тогда считались ответственными за наследственные признаки. Однако идея о том, что нуклеиновые кислоты являются материальной основой наследственности, в науке не утвердилась. Так, советский биолог Н.К. Кольцов считал, что за передачу наследственности отвечает белковая компонента хромосом. Считалось также, что РНК встречается только у растений, а ДНК – у животных.
И только в 1936 г. благодаря работам Ж. Браше и Дж. Дэвидсона, а также А.Н. Белозерского стало ясно, что оба вида нуклеиновых кислот присуще как для животных, так и для растений.
Ф. Гриффит (1928 г.), О. Эвери (1944 г.), А. Херши (1952 г.) опытным путем доказали, что нуклеиновые кислоты обеспечивают хранение, передачу и реализацию наследственной информации в процессе синтеза белка в клетке. Они пометили ДНК и капсид радиоактивным фосфором. Заразили вирусом кишечную палочку и пришли к выводу, что внутрь клетки попадает ДНК, а капсид остается в питательной среде. Таким образом, было доказано, что ДНК является носителем наследственной информации.
В 1950-е г. Э. Чаргафф установил строгие количественные соотношения азотистых оснований в молекулах ДНК, при помощи бумажной хроматографии которые получили название правил Чаргаффа.
Важнейшие сведения были получены благодаря рентгеноструктурным исследованиям биомолекул. Л. Полинг продемонстрировал возможность спирализации белковых молекул. Первые рентгенограммы молекул ДНК, полученные английскими биофизиками М.Уилкинсом и Р. Франклин свидетельствовали о наличии в их пространственной структуре элементов периодичности.
Основываясь на этих открытиях, в 1953 г. американский генетик Джеймс Уотсон и английский физик Френсис Крик сформулировали основные представления о структуре двойной спирали ДНК. Модель ДНК Уотсона-Крика позволила объяснить многие фундаментальные биологические феномены, такие как структурная организация молекул ДНК, способ хранения и точного копирования генетической информации, возможность изменения структуры генов в процессе эволюции.
В 1957 г. Ф.Крик сформулировал центральную догму молекулярной биологии (рис.1), согласно которой наследственная информация ( генетическая) информация реализуется однонаправленно. Заключенная в последовательностях нуклеотидов ДНК информация о первичной структуре белка сначала переписывается на молекулу РНК (процесс транскрипции), а затем в процессе биосинтеза белка реализуется в последовательности аминокислотных остатков полипептидной цепи (процесс трансляции). Согласно этой концепции, ДНК является хранилищем информации о структуре белка. Посредником между ними является РНК, реализующая генетическую информацию, заключенную. в ДНК, в структуру белка.
Рибонуклеиновая кислота (РНК) — это химическое вещество, очень похожее на ДНК. Оно необходимо для передачи генетической информации от ДНК в ядре клетки ко всей клетке в целом.

Американский биохимик Роберт У. Холли был одним из лауреатов Нобелевской премии по физиологии и медицине 1968 г. за описание структуры транспортной РНК, которая влияет на механизм синтеза белка
Прошло еще несколько лет, прежде чем биологи установили, что имеется два типа нуклеиновых кислот — ДНК и РНК. В 1933 г. бельгийский биохимик Жан Браше показал, что ДНК локализована в хромосомах, тогда как РНК присутствует в цитоплазме всех клеток. Браше отметил, что клетки, богатые РНК, как правило, активно участвуют в синтезе белка, и пришел к выводу, что РНК имеет отношение к этому процессу. Механизм, связывающий ДНК, РНК и синтез белка, сегодня известен как центральная догма молекулярной биологии, но он открылся ученым, только когда стало ясно строения ДНК, то есть в 1953 г.
Вирус гриппа не содержит ДНК, генетическую информацию о нем несет РНК. Вирус не может размножаться самостоятельно: для этого он захватывает клетки организма-хозяина, вызывая знакомые всем симптомы
Типы РНК
ДНК имеет в составе азотистые основания — аденин, цитозин, гуанин и тимин, и их последовательность определяет генетический код. РНК имеет сходную структуру, но вместо тимина содержит урацил. Кроме того, РНК — это, как правило, одинарная нить, тогда как ДНК — спираль, состоящая из двух нитей. РНК из одной нити — известная как матричная РНК (мРНК) — передает генетическую информацию в клетку. Эту информацию конвертируют в белки структуры под названием рибосомы; они состоят частично из переплетенных молекул РНК (рибосомной РНК). мРНК проходит через рибосому, при помощи транспортной РНК (тРНК) считывает последовательность оснований, и выстраивается белок.
Честно говоря, насчет РНК никто не задумывался долгие годы. Существовала догма, что вот есть клетка, есть хромосомы, в которых есть ДНК — хранитель генетической информации.
В конце концов, на рибосомах синтезируются белки. А РНК — она где-то в промежутке, переносчик информации от ДНК — и только. А потом посыпались открытия, которые заставили совершенно по-другому взглянуть на РНК Главное отличие нуклеиновых кислот заключается в их углеводной компоненте. В РНК сахар — рибоза, а в ДНК — дезоксирибоза: там, где у ДНК имеется атом водорода (Н), у РНК стоит оксигруппа (ОН). Результаты таких незначительных, на неискушенный взгляд, различий поражают. Так, ДНК существуют в основном в форме всем известных жестких спиралей, в которых две цепи ДНК удерживаются вместе за счет образования водородных связей между комплементарными нуклеотидами.
РНК также могут формировать спирали из двух цепочек, похожие на спирали ДНК, однако в большинстве случаев РНК существуют в виде сложных структур-клубков. Структуры эти формируются не только за счет образования упомянутых водородных связей между разными участками РНК, но и благодаря оксигруппе рибозы, которая может образовывать дополнительные водородные связи и взаимодействовать с фосфорной кислотой и ионами металлов. Глобулярные структуры РНК не только внешне напоминают белковые структуры, но и приближаются к ним по свойствам: они могут взаимодействовать с самыми разными молекулами, как маленькими, так и полимерными.
Почему же именно РНК мы называем праматерью ныне существующей жизни? Чтобы ответить на этот вопрос, давайте разберемся, где проходит граница между живым и неживым.
РНК умеет все?
Чем больше углублялись в изучение различных биосинтетических процессов, тем чаще обнаруживали ранее неизвестные функции РНК. Оказалось, что кроме процесса транскрипции (синтеза РНК путем копирования участка ДНК) в ряде случаев, наоборот, может происходить синтез ДНК на РНК-матрицах. Этот процесс, названный обратной транскрипцией, используют в ходе своего развития многие вирусы, в том числе печально известные онкогенные вирусы и ВИЧ-1, вызывающий СПИД.
Таким образом, выяснилось, что поток генетической информации не является, как первоначально считалось, однонаправленным — от ДНК к РНК. Роль ДНК как изначально главного носителя генетической информации стала подвергаться сомнению. Тем более что многие вирусы (гриппа, клещевого энцефалита и другие) вообще не используют ДНК в качестве генетического материала, их геном построен исключительно из РНК. А далее посыпались одно за другим открытия, которые заставили совершенно по-другому взглянуть на РНК.
Наиболее удивительным было открытие каталитической способности РНК. Прежде считалось, что катализировать реакции умеют только белки, ферменты. Ученые, например, никак не могли выделить ферменты, осуществляющие разрезание и сшивание некоторых РНК. После длительных исследований выяснилось, что РНК прекрасно справляются с этим сами. Структуры РНК, действующие подобно ферментам, назвали рибозимами (по аналогии с энзимами, белками-катализаторами). Вскоре было обнаружено множество разнообразных рибозимов. Особенно широко их используют для манипулирования своими РНК вирусы и другие простые инфекционные агенты. Таким образом, РНК оказались мастерами на все руки: они могут выступать в роли носителей наследственной информации, могут служить катализаторами, транспортными средствами для аминокислот, образовывать высокоспецифичные комплексы с белками.
Конечно, полную картину еще предстоит реконструировать — осталось много нерешенных вопросов. Например, в современной клетке активацию аминокислот и их присоединение к соответствующим тРНК осуществляют специфичные белки-ферменты. Возникают вопросы: могла ли эта реакция осуществляться без участия белков, только с помощью РНК? Могли ли сами РНК катализировать синтез РНК из нуклеотидов или присоединение азотистых оснований к сахару? В общем-то, после открытия рибозимов такие потенциальные способности РНК уже не вызывали особых сомнений. Но наука требует, чтобы гипотезы экспериментально подтверждались.
Дарвиновская Эволюция в Пробирке
Хороший метод зачастую позволяет осуществить революцию в науке. Именно так можно сказать о методе полимеразной цепной реакции (ПЦР), который позволяет размножать нуклеиновые кислоты в неограниченных количествах. Кратко опишем суть метода. Для размножения ДНК в методе ПЦР используются ферменты ДНК- полимеразы, т. е. те самые ферменты, которые при размножении клеток синтезируют из активированных мономеров-нуклеотидов комплементарные цепочки ДНК.
При методе ПЦР в пробирку с ДНК вносят смесь активированных нуклеотидов, фермент ДНК-полимеразу и так называемые праймеры — олигонуклеотиды, комплементарные концам размножаемой ДНК. При нагревании раствора цепи ДНК расходятся. Затем, при охлаждении, с ними связываются праймеры, образуя короткие фрагменты спиральных структур. Фермент присоединяет к праймерам нуклеотиды и собирает цепочку, комплементарную цепочке исходной ДНК. В результате реакции из одной двуцепочечной ДНК получается две. Если повторить процесс, получится четыре цепочки, а после n повторений — 2 n молекул ДНК. Все очень просто.
Изобретение ПЦР и разработка методов химического синтеза ДНК позволили создать потрясающую технологию молекулярной селекции. Принцип молекулярной селекции тоже прост: сначала синтезируется множество молекул, обладающих разными свойствами (так называемая молекулярная библиотека), а затем из этой смеси отбираются молекулы с желаемым свойством.
Библиотеки нуклеиновых кислот — это смеси молекул, имеющих одинаковую длину, но отличающихся последовательностью нуклеотидов. Получить их можно в том случае, если при химическом синтезе на авто-матическом синтезаторе добавлять на каждой стадии удлинения нуклеотидной последовательности одно-временно все четыре нуклеотида. Каждый из них будет включаться в растущую нуклеиновую кислоту с равной вероятностью, в результате чего на каждом этапе присоединения будет получаться 4 варианта последовательностей. Если таким образом синтезировать нуклеиновую кислоту длиной в n звеньев, то разнообразие полученных молекул составит 4 в степени n. Поскольку обычно используются участки длиной 30—60 мономеров, то в результате синтеза получается от 4 30 до 4 60 разных молекул! Цифры, привычные разве что для астрономов.
С помощью такого метода были получены тысячи разных РНК-аптамеров, которые образуют специфические комплексы с различными органическими соединениями и молекулами.
Рассмотренная схема молекулярной селекции может быть применена для получения молекул с любыми свойствами. Например, были получены РНК, способные катализировать реакции синтеза РНК и белков: присоединение азотистых оснований к рибозе, полимеризацию активированных нуклеотидов на цепочках РНК, присоединение аминокислот к РНК. Эти исследования еще раз подтвердили, что в условиях предбиологической эволюции из случайных полимеров могли возникать молекулы РНК
со специфическими структурами и функциями.
Делайте Ваш Заказ!
Метод молекулярной селекции обладает очень большими возможностями. С его помощью можно решать задачи поиска нужных молекул даже в том случае, если исходно нет идеи, как такие молекулы должны быть устроены. Однако, если придумать процедуру отбора, их можно выделить по принципу требуемых свойств, а затем уже заняться и вопросом, как эти свойства достигаются. Продемонстрируем это на примере выделения РНК, способных связываться с клеточными мембранами и модулировать их проницаемость.
Мембраны современных клеток и липосом, построенные из жирных кислот, несут отрицательный заряд. Поскольку РНК также заряжены отрицательно, то по закону Кулона они должны отталкиваться от липидной поверхности и тем более не могут проникать в глубь липидного слоя. Единственный известный способ взаимодействия нуклеиновых кислот с поверхностью мембран — через двухзарядные ионы металлов. Эти положительно заряженные ионы могут играть роль мостиков, располагаясь между отрицательно заряженными группами на поверхности мембраны и фосфатными группами нуклеиновой кислоты. Поскольку такие мостиковые взаимодействия достаточно слабые, с мембраной может связаться только очень большая нуклеиновая кислота благодаря множеству слабых связей с поверхностью мембраны. Так маленькие враги привязали Гулливера к земле множеством тоненьких веревок.
Этот селекционный эксперимент подсказал, что у РНК есть дополнительный способ приобретения новых свойств путем образования сложных надмолекулярных комплексов. Этот механизм мог использоваться и для удерживания эволюционирующих систем РНК в виде колоний на поверхностях еще до того, как эти системы обзавелись изолирующей мембраной.
Редакция благодарит сотрудников Института химической биологии и фундаментальной медицины
СО РАН к. х. н. В. В. Коваля, к. х. н. С. Д. Мызину и к. х. н. А. А. Бондаря за помощь в подготовке статьи

Пре-мРНК со стеблем-петлёй. Атомы азота в основаниях выделены голубым, кислорода в фосфатном остове молекулы — красным
Рибонуклеи́новые кисло́ты (РНК) — нуклеиновые кислоты, полимеры нуклеотидов, в состав которых входят остаток ортофосфорной кислоты, рибоза (в отличие от ДНК, содержащей дезоксирибозу) и азотистые основания — аденин, цитозин, гуанин и урацил (в отличие от ДНК, содержащей вместо урацила тимин). Эти молекулы содержатся в клетках всех живых организмов, а также в некоторых вирусах.
Клеточные РНК образуются в ходе процесса, называемого транскрипцией, то есть синтеза РНК на матрице ДНК, осуществляемого специальными ферментами — РНК-полимеразами. Затем матричные РНК (мРНК), принимают участие в процессе, называемом трансляцией. Трансляция — это синтез белка на матрице мРНК при участии рибосом. Другие РНК после транскрипции подвергаются химическим модификациям, и после образования вторичной и третичной структур выполняют функции, зависящие от типа РНК.
Для одноцепочечных РНК характерны разнообразные пространственные структуры, в которых часть нуклеотидов одной и той же цепи спарены между собой. Некоторые высокоструктурированные РНК принимают участие в синтезе белка клетки, например, транспортные РНК служат для узнавания кодонов и доставки соответствующих аминокислот к месту синтеза белка, а рибосомные РНК служат структурной и каталитической основой рибосом.
Однако функции РНК в современных клетках не ограничиваются их ролью в трансляции. Так малые ядерные РНК принимают участие в сплайсинге эукариотических матричных РНК и т. д.
Геномы некоторых вирусов состоят из РНК, то есть у них она выполняет роль, которую у высших организмов выполняет ДНК. На основании разнообразия функций РНК в клетке была выдвинута гипотеза, согласно которой РНК — первая молекула, которая была способна к самовоспроизведению в добиологических системах.
Содержание
История изучения
Химический состав и модификации мономеров

Структура
Азотистые основания в составе РНК могут образовывать водородные связи между цитозином и гуанином, аденином и урацилом, а также между гуанином и урацилом [15] . Однако возможны и другие взаимодействия, например, несколько аденинов могут образовывать петлю, или петля, состоящая из четырёх нуклеотидов, в которой есть пара оснований аденин — гуанин [16] .

Разные формы нуклеиновых кислот. На рисунке (слева направо) представлены A (типична для РНК), B (ДНК) и Z (редкая форма ДНК)
Важная структурная особенность РНК, отличающая её от ДНК — наличие гидроксильной группы в 2' положении рибозы, которая позволяет молекуле РНК существовать в А, а не В-конформации, наиболее часто наблюдаемой у ДНК [17] . У А-формы глубокая и узкая большая бороздка и неглубокая и широкая малая бороздка [18] . Второе последствие наличия 2' гидроксильной группы состоит в том, что конформационно пластичные, то есть не принимающие участие в образовании двойной спирали, участки молекулы РНК могут химически атаковать другие фосфатные связи и их расщеплять [19] .
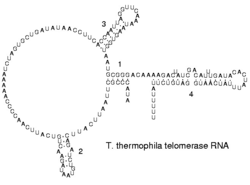
Примером зависимости функции молекул РНК от их вторичной структуры являются участки внутренней посадки рибосомы (стресса [22] .
Многие типы РНК, например, рРНК и мяРНК в клетке функционируют в виде комплексов с белками, которые ассоцииируют с молекулами РНК после их синтеза или (у эукариот) экспорта из ядра в цитоплазму. Такие РНК-белковые комплексы называются рибонуклеопротеиновыми комплексами или рибонуклеопротеидами.
Сравнение с ДНК
Между ДНК и РНК есть три основных отличия:
- ДНК содержит сахар дезоксирибозу, РНК — рибозу, которая содержит одну дополнительную, по сравнению с дезоксирибозой, гидроксильную группу. Эта группа увеличивает вероятность гидролиза молекулы, то есть уменьшает стабильность молекулы РНК. , комплементарный аденину, в РНК не тимин, как в ДНК, а урацил — неметилированная форма тимина.
- ДНК существует в форме двойной спирали, состоящей из двух отдельных молекул. Молекулы РНК, в среднем, гораздо короче и преимущественно одноцепочечные.
Структурный анализ биологически активных молекул РНК, включая тРНК, рРНК, мяРНК и другие молекулы, которые не кодируют белков показал, что они состоят не из одной длинной спирали, а из многочисленных коротких спиралей, расположенных близко друг к другу и образующих нечто, похожее на третичную структуру белка. В результате этого РНК может катализировать химические реакции, например, пептидил-трансферазный центр рибосомы, участвующий в образовании пептидной связи белков, полностью состоит из РНК [23] [24] .
Синтез
Синтез РНК в живой клетке проводится ферментом — РНК-полимеразой. У эукариот разные типы РНК синтезируются разными, специализированными РНК-полимеразами. В целом матрицей синтеза РНК может выступать как ДНК, так и другая молекула РНК. Например, полиовирусы используют РНК-зависимую РНК-полимеразу для репликации генетического материала, состоящего из РНК [25] . Но РНК-зависимый синтез РНК, который раньше считался характерным только для вирусов, происходит и в клеточных организмах, в процессе так называемой РНК-интерференции [26] .
Например, у кишечной палочки гены рРНК расположены в составе одного оперона (в rrnB порядок расположения такой: 16S — tRNA Glu 2 — 23S —5S) считываются в виде одной длинной молекулы, которая затем подвергается расщеплению в нескольких участках с образованием сначала пре-рРНК, а затем зрелых молекул рРНК [28] . Процесс изменения нуклеотидной последовательности РНК после синтеза носит название процессинга или редактирования РНК.

Типы РНК
Матричная (информационная) РНК — РНК, которая служит посредником при передаче информации, закодированной в ДНК к рибосомам, молекулярным машинам, синтезирующим белки живого организма. Кодирующая последовательность мРНК определяет последовательность аминокислот полипептидной цепи белка [29] . Однако подавляющее большинство РНК не кодируют белок. Эти некодирующие РНК могут транскрибироваться с отдельных генов (например, рибосомальные РНК) или быть производными интронов [30] . Классические, хорошо изученные типы некодирующих РНК — это транспортные РНК (тРНК) и рРНК, которые участвуют в процессе трансляции [31] . Существуют также классы РНК, ответственные за регуляцию генов, процессинг мРНК и другие роли. Существуют также молекулы некодирующих РНК, способные катализировать химические реакции, такие как разрезание и лигирование молекул РНК [32] . По аналогии с белками, способными катализировать химические реакции — энзимами (ферментами), каталитические молекулы РНК называются рибозимами.
Участвующие в трансляции

Информация о последовательности аминокислот белка содержится в мРНК. Три последовательных нуклеотида (кодон) соответствуют одной аминокислоте. В эукариотических клетках транскирибированный предшественник мРНК или пре-мРНК процессируется с образованием зрелой мРНК. Процессинг включает удаление некодирующих белок последовательностей (интронов). После этого мРНК экспортируется из ядра в цитоплазму, где к ней присоединяются рибосомы, транслирующие мРНК с помощью соединённых с аминокислотами тРНК.
В безъядерных клетках (бактерии и археи) рибосомы могут присоединяться к мРНК сразу после транскрипции участка РНК. И у эукариот, и у прокариот цикл жизни мРНК завершается её контролируемым разрушением ферментами рибонуклеазами [29] .
Транспортные (тРНК) — малые, состоящие из приблизительно 80 нуклеотидов, молекулы с консервативной третичной структурой. Они переносят специфические аминокислоты в место синтеза пептидной связи в рибосоме. Каждая тРНК содержит участок для присоединения аминокислоты и антикодон для узнавания и присоединения к кодонам мРНК. Антикодон образует водородные связи с кодоном, что помещает тРНК в положение, способствующее образованию пептидной связи между последней аминокислотой образованного пептида и аминокислотой, присоединённой к тРНК [30] .
Рибосомальные РНК (рРНК) — каталитическая составляющая рибосом. Эукариотические рибосомы содержат четыре типа молекул рРНК: 18S, 5.8S, 28S и 5S. Три из четырёх типов рРНК синтезируются в ядрышке. В цитоплазме рибосомальные РНК соединяются с рибосомальными белками и формируют нуклеопротеин, называемый рибосомой [29] . Рибосома присоединяется к мРНК и синтезирует белок. рРНК составляет до 80 % РНК, обнаруживаемой в цитоплазме эукариотической клетки [33] .
Необычный тип РНК, который действует в качестве тРНК и мРНК (тмРНК) обнаружен во многих бактериях и пластидах. При остановке рибосомы на дефектных мРНК без стоп-кодонов тмРНК присоединяет небольшой пептид, направляющий белок на деградацию [34] .
Участвующие в регуляции генов
В живых клетках обнаружено несколько типов РНК, которые могут уменьшать степень выражения гена при комплементарности мРНК или самому гену. Микро-РНК (21-22 нуклеотида в длину) найдены у эукариот и оказывают воздействие через механизм РНК-интерференции. При этом комплекс микро-РНК и ферментов может приводить к метилированию нуклеотидов в ДНК промотора гена, что служит сигналом для уменьшения активности гена. При использовании другого типа регуляции мРНК, комплементарная микро-РНК, деградируется. [35] . Однако есть и миРНК, которые увеличивают, а не уменьшают экспрессию генов. [36] . Малые интерферирующие РНК (миРНК, 20-25 нуклеотидов) часто образуются в результате расщепления вирусных РНК, но существуют и эндогенные клеточные миРНК [37] . Малые интерферирующие РНК также действуют через РНК-интерференцию по сходным с микро-РНК механизмам [38] . У животных найдены так называемыме РНК, взаимодействующие с Piwi (piRNA, 29-30 нуклеотидов), действующие в половых клетках против транспозиции и играющие роль в образовании гамет [39] [40] . Кроме того, piRNA могут эпигенетически наследоваться по материнской линии, передавая потомству своё свойство ингибировать экспрессию транспозонов [41] .
Антисмысловые РНК широко распространены у бактерий, многие из них подавляют выражение генов, но некоторые активируют экспрессию [42] . Действуют антисмысловые РНК, присоединяясь к мРНК, что приводит к образованию двуцепочечных молекул РНК, которые деградируются ферментами. [43] . У эукариот обнаружены высокомолекулярные, мРНК-подобные молекулы РНК. Эти молекулы также регулируют выражение генов, [44] . В качетве примера можно привести Xist, присоединяющуюся и инактивирующую одну из двух Х-хромосом у самок млекопитающих. [45] .
Кроме роли отдельных молекул в регуляции генов, регуляторные элементы могут формироваться в 5' и 3' нетранслируемых участках мРНК. Эти элементы могут действовать самостоятельно, предотвращая инициацию трансляции, либо присоединять белки, например, ферритин или малые молекулы, например, биотин. [46] .
В процессинге РНК
Многие РНК принимают участие в модификации других РНК. Интроны вырезаются из пре-мРНК сплайсосомами, которые, кроме белков, содержат несколько малых ядерных РНК (мяРНК) [31] . Кроме того, интроны могут катализировать собственное вырезание. [47] . Синтезированая в результате транскрипции РНК также может быть химически модифицирована. У эукариот химические модификации нуклеотидов РНК, например, их метилирование, выполняется малыми ядерными РНК (мяРНК, 60-300 нуклеотидов). Этот тип РНК локализуется в ядрышко и тельцах Кахаля [30] . После ассоциации мяРНК с ферментами, мяРНК связываются с РНК-мишенью путём образования пар между основаниями двух молекул, а ферменты модифицируют нуклеотиды РНК-мишени. Рибосомальные и транспортные РНК содержат много подобных модификаций, конкретное положение которых часто сохраняется в процессе эволюции. Также могут быть модифицированы мяРНК и сами мяРНК [48] [49] Гидовые РНК осуществляют процесс редактирования РНК в кинетопласте - особом участке митохондрии протистов-кинетопластид (например, трипаносом).
Геномы, состоящие из РНК

Как и ДНК, РНК может хранить информацию о биологических процессах. РНК может использоваться в качестве генома вирусов и вирусоподобных частиц. РНК-геномы можно разделить на те, которые не имеют промежуточной стадии ДНК и которые для размножения копируются в ДНК-копию и обратно (ретровирусы).
РНК-содержащие вирусы
Многие вирусы, например, вирус гриппа, на всех стадиях содержат геном, состоящий исключительно из РНК. РНК содержится внутри обычно белковой оболочки и реплицируется с помощью закодированных в ней РНК-зависимых РНК-полимераз. Вирусные геномы, состоящие из РНК разделяются на
Вироиды — другая группа патогенов, содержащих РНК-геном и не содержащих белок. Они реплицируются РНК-полимеразами организма хозяина [50] .
Ретровирусы и ретротранспозоны
У других вирусов РНК-геном есть в течение только одной из фаз жизненного цикла. Вирионы так называемых ретровирусов содержат молекулы РНК, которые при попадании в клетки хозяина служат матрицей для синтеза ДНК-копии. В свою очередь, с матрицы ДНК считывается РНК-геном. Кроме вирусов обратную транскрипции применяют и класс мобильных элементов генома — ретротранспозоны [51]
Гипотеза РНК-мира
РНК - класс нуклеиновых кислот, линейных полимеров нуклеотидов, в состав которых входят остаток фосфорной кислоты, рибоза (в отличие от ДНК, содержащей дезоксирибозу) и азотистые основания - аденин, цитозин, гуанин и урацил (в отличие от ДНК, содержащий вместо урацила тимин). Эти молекулы содержатся в клетках всех живых организмов, а также в некоторых вирусов.
РНК содержатся главным образом в цитоплазме клеток. Эти молекулы синтезируются в клетках всех клеточных живых организмов, а также содержатся в вироидах и некоторых вирусах.
Основные функции РНК в клеточных организмах - это шаблон для трансляции генетической информации в белки и поставка соответствующих аминокислот к рибосомам. В вирусах является носителем генетической информации (кодирует белки оболочки и ферменты вирусов). Вироиды состоят из кольцевой молекулы РНК и не содержат в себе других молекул. Существует гипотеза мира РНК, согласно которой, РНК возникли перед белками и были первыми формами жизни.
Клеточные РНК образуются в ходе процесса, называемого транскрипцией, то есть синтеза РНК на матрице ДНК, осуществляемого специальными ферментами - РНК-полимеразы. Затем матричные РНК (мРНК) участвуют в процессе, называемом трансляцией. Трансляция - это синтез белка на матрице мРНК при участии рибосом. Другие РНК после транскрипции подвергаются химическим модификациям, и после образования вторичной и третичной структур выполняют функции, зависящие от типа РНК.
Для одноцепочечной РНК характерны разнообразные пространственные структуры, в которых часть нуклеотидов одной и той же цепи спарены между собой. Некоторые высокоструктурированные РНК принимают участие в синтезе белка клетки, например, транспортные РНК служат для узнавания кодонов и доставки соответствующих аминокислот к месту синтеза белка, а матричные РНК служат структурной и каталитической основой рибосом.
Однако функции РНК в современных клетках не ограничиваются их ролью в трансляции. Так мРНК участвуют в сплайсинге эукариотических матричных РНК и других процессах.
Геномы ряда вирусов состоят из РНК, то есть у них она играет роль, которую в высших организмах выполняет ДНК. На основании разнообразия функций РНК в клетке была выдвинута гипотеза, согласно которой РНК - первая молекула, способная к самовоспроизведению в добиологических системах.
История изучения РНК
Значение РНК в синтезе белков было предположено в 1939 году в работе Торберна Оскара Касперссона, Жана Брачета и Джека Шульца. Джерард Маирбакс выделил первую матричную РНК, кодирующую гемоглобин кролика и показал, что при ее введении в ооциты образуется тот же самый белок.
В Советском Союзе в 1956-57 годах проводились работы (А. Белозерский, А. Спирин, Э. Волкин, Ф. Астрахан) по определению состава РНК клеток, которые привели к выводу, что основную массу РНК в клетке составляют рибосомные РНК.
В 1959 году Северо Очоа получил Нобелевскую премию по медицине за открытие механизма синтеза РНК. Последовательность из 77 нуклеотидов одной из тРНК дрожжей S. cerevisiae была определена в 1965 году в лаборатории Роберта Холле, за что в 1968 году он получил Нобелевскую премию по медицине.
В 1967 Карл Везе предположил, что РНК имеют каталитические свойства. Он выдвинул так называемую Гипотезу РНК-мира, в котором РНК прото-организмов служили и как молекулы хранения информации (сейчас эта роль выполняется ДНК) и как молекулы, которые катализировали метаболические реакции (сейчас это делают ферменты).
В 1976 Уолтер Фаерс и его группа из Гентского университета (Голландия) впервые определили последовательность генома РНК - содержащегося в вирусе, бактериофага MS2.
В начале 1990-х было обнаружено, что введение чужеродных генов в геном растений приводит к подавлению выражения аналогичных генов растения. Примерно в это же время было показано, что РНК длиной около 22 оснований, которые сейчас называются микро-РНК, играют регуляторную роль в онтогенезе круглых червей.
Гипотеза о значении РНК в синтезе белков была высказана Торбьерном Касперссоном (Torbjörn Caspersson) на основе исследований 1937-1939 гг., в результате которых было показано, что клетки, активно синтезирующие белок, содержат большое количество РНК. Подтверждение гипотезы было получено Юбером Шантренном (Hubert Chantrenne).
Особенности строения РНК
Нуклеотиды РНК состоят из сахара - рибозы, к которой в положении 1 'присоединена одна из основ: аденин, гуанин, цитозин или урацил. Фосфатная группа объединяет рибозы в цепочку, образуя связи с 3 'атомом углерода одной рибозы и в 5' положении другого. Фосфатные группы при физиологическом рН отрицательно заряжены, поэтому РНК - можно назвать полианионом.
Еще одной модифицированной основой, о которой стоит сказать является - гипоксантин, деаминованний гуанин, нуклеозид которого носит название инозин. Инозин играет важную роль в обеспечении вырожденности генетического кода.
Роль многих других модификаций не до конца изучена, но в рибосомальной РНК много пост-транскрипционных модификаций находятся в важных для функционирования рибосомы участках. Например, на одном из рибонуклеотидов, участвующих в образовании пептидной связи. Азотистые основания в составе РНК могут образовывать водородные связи между цитозином и гуанином, аденином и урацилом, а также между гуанином и урацилом. Однако возможны и другие взаимодействия, например, несколько аденинов могут образовывать петлю, или петля, состоящая из четырех нуклеотидов, в которой есть пара оснований аденин - гуанин.
Важная структурная особенность РНК, отличающая ее от ДНК - наличие гидроксильной группы в 2 'положении рибозы, которая позволяет молекуле РНК существовать в А, а не В-конформации, которая наиболее часто наблюдается в ДНК. В А-форме глубокая и узкая большая бороздка и неглубокая и широкая малая бороздка. Второе последствие наличия 2 'гидроксильной группы состоит в том, что конформационно пластичные, то есть, не принимающие участие в образовании двойной спирали, участки молекулы РНК могут химически атаковать другие фосфатные связи и их расщеплять.
Примером зависимости функций молекул РНК от их вторичной структуры являются участки внутренней посадки рибосомы (IRES). IRES - структура на 5 'конце информационной РНК, которая обеспечивает присоединение рибосомы в обход обычного механизма инициации синтеза белка, требует наличия особого модифицированного основания (кэпа) на 5' конце и белковых факторов инициации. Сначала IRES были обнаружены в вирусных РНК, но сейчас накапливается все больше данных о том, что клеточные мРНК также используют IRES-зависимый механизм инициации в условиях стресса. Многие типы РНК, например, рРНК и мяРНК (мяРНК) в клетке функционируют в виде комплексов с белками, которые ассоциируют с молекулами РНК после их синтеза или (у эукариот) экспорта из ядра в цитоплазму. Такие РНК-белковые комплексы называются рибонуклеопротеиновыми комплексами или рибонуклеопротеидами.
Подавляющее большинство РНК не кодируют белок. Эти некодирующие РНК могут транскрибировать из отдельных генов (например, рибосомальные РНК) или быть производными интронов. Классические, хорошо изученные типы некодирующих РНК - это транспортные РНК (тРНК) и рРНК, участвующие в процессе трансляции. Существуют также классы РНК, ответственные за регуляцию генов, процессинг мРНК и другие роли. Кроме того, есть и молекулы некодирующих РНК, способные катализировать химические реакции, такие, как разрезание и лигирование молекул РНК. По аналогии с белками, способными катализировать химические реакции - энзимами (ферментами), каталитические молекулы РНК называются рибозимами.
Транспортные (тРНК) - малые, состоящие из примерно 80 нуклеотидов, молекулы с консервативной третичной структурой. Они переносят специфические аминокислоты к месту синтеза пептидной связи в рибосоме. Каждая тРНК содержит участок для присоединения аминокислоты и антикодон для узнавания и присоединения к кодону мРНК. Антикодон образует водородные связи с кодоном, что помещает тРНК в положение, способствующее образованию пептидной связи между последней аминокислотой образованного пептида и аминокислотой, присоединенной к тРНК.
Рибосомальные РНК (рРНК) - каталитическая составляющая рибосом. Эукариотические рибосомы содержат четыре типа молекул рРНК: 18S, 5.8S, 28S и 5S. Три из четырех типов рРНК синтезируются на полисомах. В цитоплазме рибосомальные РНК соединяются с рибосомальными белками и формируют нуклеопротеины, называемые рибосомами. Рибосома присоединяется к мРНК и синтезирует белок. рРНК составляет до 80% РНК, обнаруживается в цитоплазме эукариотической клетки.
Необычный тип РНК, который действует в качестве тРНК и мРНК (тмРНК) обнаружен во многих бактериях и пластидах. При остановке рибосомы на дефектных мРНК без стоп-кодонов тмРНК присоединяет небольшой пептид, направляющий белок на деградацию.
Микро-РНК (21-22 нуклеотида в длину) найдены у эукариот и влияют через механизм РНК-интерференции. При этом комплекс микро-РНК и ферментов может приводит к метилированию нуклеотидов в ДНК промотора гена, что служит сигналом для уменьшения активности гена. При использовании другого типа регуляции мРНК, комплементарная микро-РНК, деградирует. Однако есть и миРНК, которые увеличивают, а не уменьшают экспрессию генов.
Малые интерферирующие РНК (миРНК, 20-25 нуклеотидов) часто образуются в результате расщепления вирусных РНК, но существуют и эндогенные клеточные миРНК. Малые интерферирующие РНК также действуют через РНК-интерференцию по сходным с микро-РНК механизмам.
Сравнение с ДНК
Между ДНК и РНК есть три основных отличия:
1. ДНК содержит сахар дезоксирибозу, РНК - рибозу, у которой есть дополнительная, по сравнению с дезоксирибозой, гидроксильная группа. Эта группа увеличивает вероятность гидролиза молекулы, то есть уменьшает стабильность молекулы РНК.
2. Нуклеотид, комплементарный аденину, в РНК не тимин, как в ДНК, а урацил - неметилированная форма тимина.
3. ДНК существует в форме двойной спирали, состоящей из двух отдельных молекул. Молекулы РНК, в среднем, намного короче и преимущественно одноцепочечные. Структурный анализ биологически активных молекул РНК, включая тРНК, рРНК мяРНК и другие молекулы, которые не кодируют белков, показал, что они состоят не из одной длинной спирали, а из многочисленных коротких спиралей, расположенных близко друг к другу и образуют нечто, похожее на третичную структуру белка. В результате этого РНК может катализировать химические реакции, например, пептид-трансферазный центр рибосомы, участвующий в образовании пептидной связи белков, полностью состоит из РНК.
Особенности функций:
1. Процессинг
Многие РНК принимают участие в модификации других РНК. Интроны вырезают из про-мРНК сплайсосомы, которые, кроме белков, содержат несколько малых ядерных РНК (мяРНК). Кроме того, интроны могут катализировать собственное вырезание. Синтезированная в результате транскрипции РНК также может быть химически модифицирована. У эукариот химические модификации нуклеотидов РНК, например, их метилирование, выполняется малыми ядерными РНК (мяРНК, 60-300 нуклеотидов). Этот тип РНК локализуется в ядрышке и тельцах Кахаля. После ассоциации мяРНК с ферментами, мяРНК связывается с РНК-мишенью путем образования пар между основаниями двух молекул, а ферменты модифицируют нуклеотиды РНК-мишени. Рибосомальные и транспортные РНК содержат много подобных модификаций, конкретное положение которых часто сохраняется в процессе эволюции. Также могут быть модифицированы мяРНК и сами мяРНК.
2. Трансляция
тРНК присоединяют определенные аминокислоты в цитоплазме и направляется к месту синтеза белка на иРНК где связывается с кодоном и отдает аминокислоту которая используется для синтеза белка.
3. Информационная функция
У некоторых вирусов РНК выполняет те функции которые ДНК выполняет у эукариот. Также информационную функцию выполняет иРНК которая переносит информацию о белках и является местом его синтеза.
4. Регуляция генов
Некоторые типы РНК участвуют в регуляции генов увеличивая или уменьшая его активность. Это так называемые миРНК (малые интерферирующие РНК) и микро-РНК.
5. Каталитическая функция
Есть так называемые ферменты которые относятся к РНК они называются рибозимы. Эти ферменты выполняют различные функции и имеют своеобразное строение
Читайте также: