
Ионный состав цитоплазмы кратко
Обновлено: 04.07.2024
Состав плазмы и межклеточной жидкости. Компоненты внутриклеточной жидкости
Сравнение состава внутриклеточной жидкости с внеклеточной, состоящей из плазмы и межклеточной жидкости, приведено на рисунках и таблице ниже.
Основные катионы и анионы внутри- и внеклеточной жидкостей. Концентрации ионов Ca 2+ и Mg 2+ на схеме представлены в виде суммы. На рисунке приведено общее содержание свободных и связанных ионов Неэлектролиты плазмы
а) Ионный состав плазмы и межклеточной жидкости одинаков. Поскольку плазма и межклеточная жидкость разделены высокопроницаемой мембраной капилляров, ионный состав этих жидкостей почти одинаков. Наиболее важное различие между этими жидкими средами заключается в более высокой концентрации белка в плазме. Вследствие низкой проницаемости капилляров в межклеточное пространство большинства тканей проникает лишь небольшое количество белка.
Концентрация положительно заряженных ионов (катионов) в плазме немного выше (примерно на 2%) по сравнению с межклеточной жидкостью, что обусловлено эффектом Доннана. Белки плазмы в сумме заряжены отрицательно, следовательно, они способствуют присоединению катионов, таких как ионы Na+ и К+, удерживая их в плазме в несколько большем количестве. Напротив, в межклеточной жидкости отрицательно заряженные ионы (анионы) присутствуют в слегка повышенной по сравнению с плазмой концентрации, т.к. отрицательный заряд белков плазмы отталкивает анионы. На практике, однако, принято считать, что концентрации ионов в межклеточной жидкости и плазме одинаковы.
Возвращаясь к рисунку выше, можно видеть, что внеклеточная жидкость содержит большое количество ионов Na+ и Cl- и достаточно высокое число бикарбонатов; содержание же калия, кальция, магния, фосфатов и ионов органических кислот в ней незначительно.
Состав внеклеточной жидкости тщательно регулируется с помощью различных механизмов, в особенности почечных, которые будут рассмотрены далее. Благодаря этим механизмам клетки постоянно находятся в среде с концентрацией электролитов и питательных веществ, необходимой для жизнедеятельности.
б) Основные компоненты внутриклеточной жидкости. Внутриклеточная жидкость> отделена от внеклеточной цитоплазматической мембраной, высокопроницаемой для воды и практически непроницаемой для большинства электролитов. Внутриклеточная жидкость в отличие от внеклеточной содержит лишь небольшое количество ионов натрия и хлора, а ионы кальция в ней практически отсутствуют. Внутри клетки, напротив, содержится очень большое количество ионов калия, а также умеренное число ионов магния и сульфатов; концентрация всех перечисленных веществ вне клетки низка. Кроме того, в клетках содержится большое количество белка, в 4 раза превышающее его содержание в плазме.
в) Измерение объемов жидких сред организма на основе принципа разведения индикатора. Объем жидкой среды можно рассчитать, помещая в нее вещество-индикатор. Позволив индикатору равномерно распределиться во всем объеме жидкости, устанавливают степень изменения его концентрации в растворе.
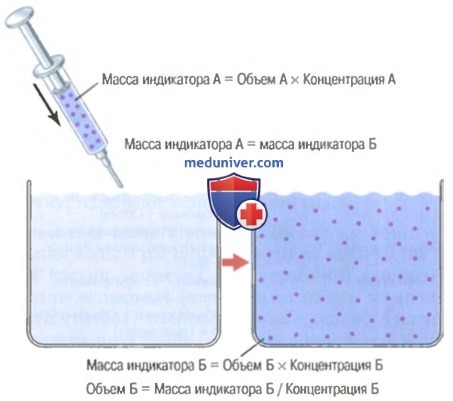
Метод разведения индикатора для измерения объемов жидкости
На рисунке выше показано, как объем жидкой среды измеряют с помощью метода разведения индикатора, основанного на законе сохранения массы. Согласно данному закону, общее количество вещества после перемешивания в жидкой среде будет равно массе вещества, введенной в данный объем.
На рисунке выше показано, как к емкости, содержащей неизвестный объем жидкости, с помощью шприца добавляют небольшое количество красителя или другого индикатора. Веществу дают возможность равномерно распределиться в жидкости, пока в любой точке объема его концентрация не станет одинаковой. Затем в образце отобранной жидкости измеряют концентрацию индикатора в растворе с помощью химического, фотоэлектроколориметрического или иного метода. Если утечки индикаторного вещества из данного объема не происходит, то его общее количество (объем Б х концентрация Б) будет равна количеству вещества, введенного шприцем (объем А х концентрация А).
Простое преобразование этого уравнения позволяет рассчитать неизвестный объем Б по формуле:
Объем Б = (объем А*концентрация А)/концентрацию Б.
Таким образом, для расчета необходимо знать два параметра: (1) общее количество вещества, введенного в объем жидкости (в числителе); (2) концентрацию индикаторного вещества в жидкости после перемешивания (в знаменателе).
Например, если 1 мл индикатора в концентрации 10 мг/мл ввели в емкость Б и после его разведения концентрация в каждом миллилитре раствора составит 0,01 мг/мл, то неизвестный объем будет равен: 1000 мл.
Объем Б = (1 мл*10 мг/мл)/0,01 мг/мл = 1000 мл.
Данный метод можно применять для измерения объема практически любой жидкой среды организма, если при этом соблюдаются следующие условия: (1) индикатор распределяется во всем объеме жидкости; (2) индикатор находится только в той среде, где измеряют его концентрацию; (3) индикатор не метаболизируется и не выводится из организма. Требованиям, необходимым для измерения объемов жидких сред организма, удовлетворяют несколько таких веществ.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Поскольку плазма и межклеточная жидкость разделены высокопроницаемой мембраной капилляров, ионный состав этих жидкостей почти одинаков. Однако, проницаемость клеточной мембраны для различных ионов различна.
Концентрация положительно заряженных ионов (катионов) в плазме немного выше (примерно на 2%) по сравнению с межклеточной жидкостью. Белки плазмы в сумме заряжены отрицательно, следовательно, они способствуют присоединению катионов, таких как ионы Na+ и К+, удерживая их в плазме в несколько большем количестве. Напротив, в межклеточной жидкости отрицательно заряженные ионы (анионы) присутствуют в слегка повышенной по сравнению с плазмой концентрации, т.к. отрицательный заряд белков плазмы отталкивает анионы. Внеклеточная жидкость содержит большое количество ионов Na+ и Cl- и достаточно высокое число бикарбонатов; содержание же калия, кальция, магния, фосфатов и ионов органических кислот в ней незначительно. Внутриклеточная жидкость, в отличие от внеклеточной, содержит лишь небольшое количество ионов натрия и хлора, а ионы кальция в ней практически отсутствуют. Внутри клетки, напротив, содержится очень большое количество ионов калия, а также умеренное число ионов магния и сульфатов; концентрация всех перечисленных веществ вне клетки низка. Кроме того, в клетках содержится большое количество белка, в 4 раза превышающее его содержание в плазме.
Мембранный потенциал — разность электрических потенциалов между наружной и внутренней поверхностями биологической мембраны, обусловленная неодинаковой концентрацией ионов. Внутренняя поверхность заряжена отрицательно по отношению к наружной. Чем больше разность потенциалов между сторонами мембраны, тем больше ток.
В состоянии покоя мембрана клетки мало проницаема для ионов Na°. При возбуждении проницаемость мембраны увеличивается, и она пропускает положительно заряженные ионы натрия внутрь клетки, что приводит к понижению мембранной разности потенциалов (деполяризации мембраны) и даже к появлению разности потенциалов противоположного знака.
71. Потенциал покоя клетки. Уравнение Гольдмана-Ходжкина-Катца.
Потенциал покоя — мембранный потенциал возбудимой клетки в невозбужденном состоянии. Он представляет собой разность электрических потенциалов, имеющихся на внутренней и наружной сторонах мембраны и составляет у теплокровных от -60 до -100 мВ. -60мВ – порог возбудимости.
Условия возникновения потенциала покоя:
Наличие градиентов концентрации ионов на цитомембране.
Различная проницаемость мембраны для различных ионов.
Мембрана возбудимой клетки способна быстро менять свою проницаемость для определённого вида катионов, вызывая переход положительного заряда с внешней стороны на внутреннюю.

Уравнение Гольдмана-Ходжина-Катца.
ЭДС=(RT/Fz)*ln*(pK +[Ko]+pNa+[Nao]+…+pA - [Ai - ])/(pK+[Ki]+pNa+[Nai]+…+pA – [Ao - ])
R - универсальная газовая постоянная, Т - абсолютная температура растворов, F - число Фарадея, p - проницаемость мембраны, [] – концентрация иона, i – внутри клетки, о – снаружи, А – анион.
Итак, мембранный потенциал покоя - это дефицит положительных электрических зарядов внутри клетки, возникающий за счёт утечки из неё положительных ионов калия и электрогенного действия натрий-калиевого насоса.
Для поддержания внутреннего баланса, поступающие из внешней среды ионы должны удалятся из цитоплазмы и связываться вне клетки в нерастворимые минеральные соединения, которые не могли бы снова попасть в цитоплазму. Роль связывания ионов в цитоплазме и их транспортировки во внешнюю среду выполняют низкомолекулярные белки типа кальций-связывающих протеинов В состав цитоплазмы входят все виды органических и неорганических веществ. Например,
Натрий — участвует в поддержании мембранного потенциала, генерации нервного импульса, процессы осморегуляции и создании буферной системы крови.
Калий — участвует в поддержании мембранного потенциала , генерации нервного импульса, регуляции сокращения сердечной мышцы.
Хлор — поддерживает электронейтральность клетки
В ней присутствуют также нерастворимые отходы обменных процессов и запасные питательные вещества. Основное вещество цитоплазмы — вода.
В интерстициальном внутритканевом пространстве между животными клеткаминаходится сложное межклеточноевещество, экстрацеллюлярный матрикс. У многихтканей, например в мышцах ипечени, матрикс заполняет только тонкие промежутки междуклетками, тогда как в другихтканях, таких, как соединительная, хрящевая икостная ткани, на межклеточный матрикс приходится большой объем и именно он выполняет основные функции . Три главных компонента межклеточного матрикса: прочныеколлагены, сетчатые адгезивныебелкии основноевещество,протеогликаны.
Клеточные мембраны обладают избирательной проницаемостью: через них медленно диффундируют глюкоза,аминокислоты,жирные кислоты,глицеролиионы, причем сами мембраны в известной мере активно регулируют этот процесс — одни вещества пропускают, а другие нет. Существует четыре основных механизма для поступления веществ в клетку или вывода их из клетки наружу: диффузия, осмос, активный транспорт и экзо- или эндоцитоз.
Избирательная проницаемость мембраны при пассивном транспорте обусловлена специальными каналами — интегральными белками. Они пронизывают мембрану насквозь, образовывая своего рода проход. Для элементов K, Na и Cl есть свои каналы. Относительно градиента концентрации молекулы этих элементов движутся в клетку и из неё. При раздражении каналы натриевых ионов раскрываются, и происходит резкое поступление в клетку ионов натрия. При этом происходит дисбаланс мембранного потенциала. После чего мембранный потенциал восстанавливается. Каналы калия всегда открыты, через них в клетку медленно попадают ионы калия.
В состоянии покоя внешняя поверхность клеточной мембраны имеет положительный потенциал , а внутренняя отрицательный.В момент возбуждения полярность мембраны меняется на противоположную. В результате этого между возбужденным и невозбужденным участками мембраны возникает разность потенциалов
6. Потенциал покоя клетки. Уравнение Гольдмана-Ходжкина-Катца
Потенциал покоя - это разность электрических зарядов между внутренней и наружной сторонами мембраны, когда клетка находится в состоянии физиологического покоя. Его средняя величина составляет -60 милливольт.
Потенциал, существующий на мембранах невозбужденных клеток (потенциал покоя) обусловлен полупроницаемыми свойствами клеточной мембраны и неравномерным распределением ионов между клеткой и окружающей средой. Это распределение поддерживается механизмами активного переноса, локализованными в самой мембране.
Основной вклад в создание и поддержание потенциала покоя вносят ионы натрия, калия и хлора.

φм=
i,o- концентрации ионов вне и внутри клетки. Потенциал покоя, рассчитанный по формуле Гольдмана-Ходжкина-Катца, составляет 60мВ со знаком минус со стороны внутриклеточного пространства.
Состав плазмы и межклеточной жидкости. Компоненты внутриклеточной жидкости
Сравнение состава внутриклеточной жидкости с внеклеточной, состоящей из плазмы и межклеточной жидкости, приведено на рисунках и таблице ниже.
Основные катионы и анионы внутри- и внеклеточной жидкостей. Концентрации ионов Ca 2+ и Mg 2+ на схеме представлены в виде суммы. На рисунке приведено общее содержание свободных и связанных ионов Неэлектролиты плазмы
а) Ионный состав плазмы и межклеточной жидкости одинаков. Поскольку плазма и межклеточная жидкость разделены высокопроницаемой мембраной капилляров, ионный состав этих жидкостей почти одинаков. Наиболее важное различие между этими жидкими средами заключается в более высокой концентрации белка в плазме. Вследствие низкой проницаемости капилляров в межклеточное пространство большинства тканей проникает лишь небольшое количество белка.
Концентрация положительно заряженных ионов (катионов) в плазме немного выше (примерно на 2%) по сравнению с межклеточной жидкостью, что обусловлено эффектом Доннана. Белки плазмы в сумме заряжены отрицательно, следовательно, они способствуют присоединению катионов, таких как ионы Na+ и К+, удерживая их в плазме в несколько большем количестве. Напротив, в межклеточной жидкости отрицательно заряженные ионы (анионы) присутствуют в слегка повышенной по сравнению с плазмой концентрации, т.к. отрицательный заряд белков плазмы отталкивает анионы. На практике, однако, принято считать, что концентрации ионов в межклеточной жидкости и плазме одинаковы.
Возвращаясь к рисунку выше, можно видеть, что внеклеточная жидкость содержит большое количество ионов Na+ и Cl- и достаточно высокое число бикарбонатов; содержание же калия, кальция, магния, фосфатов и ионов органических кислот в ней незначительно.
Состав внеклеточной жидкости тщательно регулируется с помощью различных механизмов, в особенности почечных, которые будут рассмотрены далее. Благодаря этим механизмам клетки постоянно находятся в среде с концентрацией электролитов и питательных веществ, необходимой для жизнедеятельности.
б) Основные компоненты внутриклеточной жидкости. Внутриклеточная жидкость> отделена от внеклеточной цитоплазматической мембраной, высокопроницаемой для воды и практически непроницаемой для большинства электролитов. Внутриклеточная жидкость в отличие от внеклеточной содержит лишь небольшое количество ионов натрия и хлора, а ионы кальция в ней практически отсутствуют. Внутри клетки, напротив, содержится очень большое количество ионов калия, а также умеренное число ионов магния и сульфатов; концентрация всех перечисленных веществ вне клетки низка. Кроме того, в клетках содержится большое количество белка, в 4 раза превышающее его содержание в плазме.
в) Измерение объемов жидких сред организма на основе принципа разведения индикатора. Объем жидкой среды можно рассчитать, помещая в нее вещество-индикатор. Позволив индикатору равномерно распределиться во всем объеме жидкости, устанавливают степень изменения его концентрации в растворе.
Метод разведения индикатора для измерения объемов жидкости
На рисунке выше показано, как объем жидкой среды измеряют с помощью метода разведения индикатора, основанного на законе сохранения массы. Согласно данному закону, общее количество вещества после перемешивания в жидкой среде будет равно массе вещества, введенной в данный объем.
На рисунке выше показано, как к емкости, содержащей неизвестный объем жидкости, с помощью шприца добавляют небольшое количество красителя или другого индикатора. Веществу дают возможность равномерно распределиться в жидкости, пока в любой точке объема его концентрация не станет одинаковой. Затем в образце отобранной жидкости измеряют концентрацию индикатора в растворе с помощью химического, фотоэлектроколориметрического или иного метода. Если утечки индикаторного вещества из данного объема не происходит, то его общее количество (объем Б х концентрация Б) будет равна количеству вещества, введенного шприцем (объем А х концентрация А).
Простое преобразование этого уравнения позволяет рассчитать неизвестный объем Б по формуле:
Объем Б = (объем А*концентрация А)/концентрацию Б.
Таким образом, для расчета необходимо знать два параметра: (1) общее количество вещества, введенного в объем жидкости (в числителе); (2) концентрацию индикаторного вещества в жидкости после перемешивания (в знаменателе).
Например, если 1 мл индикатора в концентрации 10 мг/мл ввели в емкость Б и после его разведения концентрация в каждом миллилитре раствора составит 0,01 мг/мл, то неизвестный объем будет равен: 1000 мл.
Объем Б = (1 мл*10 мг/мл)/0,01 мг/мл = 1000 мл.
Данный метод можно применять для измерения объема практически любой жидкой среды организма, если при этом соблюдаются следующие условия: (1) индикатор распределяется во всем объеме жидкости; (2) индикатор находится только в той среде, где измеряют его концентрацию; (3) индикатор не метаболизируется и не выводится из организма. Требованиям, необходимым для измерения объемов жидких сред организма, удовлетворяют несколько таких веществ.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021

Обзор
Потенциал покоя. В мембране открыты калиевые ионные каналы постоянного тока, закрыты натриевые каналы, работает насос-обменник (Na + /K + -АТФаза).
Автор
Редакторы

Давайте разбираться подробнее.
Зачем нам нужно знать, что такое потенциал покоя и как он возникает?
На эти вопросы можно ответить только в том случае, если узнать, как клетка создаёт себе разность электрических потенциалов (потенциал покоя) на мембране.
Совершенно очевидно, что для понимания того, как работает нервная система, необходимо вначале разобраться, как работает её отдельная нервная клетка — нейрон. Главное, что лежит в основе работы нейрона — это перемещение электрических зарядов через его мембрану и появление вследствие этого на мембране электрических потенциалов. Можно сказать, что нейрон, готовясь к своей нервной работе, вначале запасает энергию в электрической форме, а затем использует ее в процессе проведения и передачи нервного возбуждения.
Таким образом, наш самый первый шаг к изучению работы нервной системы — это понять, каким образом появляется электрический потенциал на мембране нервных клеток. Этим мы и займёмся, и назовём этот процесс формированием потенциала покоя.
В норме, когда нервная клетка находится в физиологическом покое и готова к работе, у неё уже произошло перераспределение электрических зарядов между внутренней и наружной сторонами мембраны. За счёт этого возникло электрическое поле, и на мембране появился электрический потенциал — мембранный потенциал покоя.
Таким образом, мембрана оказывается поляризованной. Это означает, что она имеет разный электрический потенциал наружной и внутренней поверхностей. Разность между этими потенциалами вполне возможно зарегистрировать.
В этом можно убедиться, если ввести внутрь клетки микроэлектрод, соединённый с регистрирующей установкой. Как только электрод попадает внутрь клетки, он мгновенно приобретает некоторый постоянный электроотрицательный потенциал по отношению к электроду, расположенному в окружающей клетку жидкости. Величина внутриклеточного электрического потенциала у нервных клеток и волокон, например, гигантских нервных волокон кальмара, в покое составляет около −70 мВ. Эту величину называют мембранным потенциалом покоя (МПП). Во всех точках аксоплазмы этот потенциал практически одинаков.
Ноздрачёв А.Д. и др. Начала физиологии [5].
Ещё немного физики. Макроскопические физические тела, как правило, электрически нейтральны, т.е. в них в равных количествах содержатся как положительные, так и отрицательные заряды. Зарядить тело можно, создав в нем избыток заряженных частиц одного вида, например, трением о другое тело, в котором при этом образуется избыток зарядов противоположного вида. Учитывая наличие элементарного заряда (e), полный электрический заряд любого тела можно представить как q = ±N×e, где N — целое число.
Потенциал электростатического поля φ определяется как отношение потенциальной энергии W пробного заряда q к величине этого заряда: φ = W/q, откуда следует, что потенциал численно равен потенциальной энергии, которой обладает в данной точке поля единичный положительный заряд. Единицей измерения потенциала служит вольт (1 В) [4]. В некоторых случаях потенциал электрического поля нагляднее определяется как физическая величина, численно равная работе внешних сил против сил электрического поля E при перемещении единичного положительного точечного заряда из бесконечности в данную точку. Последнее определение удобно записать следующим образом:

В электрофизиологии кроме потенциала покоя рассматриваются и другие электрические потенциалы: локальные постсинаптические и рецепторные потенциалы (возбуждающие и тормозные), электротонические и следовые потенциалы, миниатюрные потенциалы концевой пластинки, концентрационный потенциал и потенциал действия [5].
Потенциал покоя — это разность электрических потенциалов, имеющихся на внутренней и наружной сторонах мембраны, когда клетка находится в состоянии физиологического покоя. Его величина измеряется изнутри клетки, она отрицательна и составляет в среднем −70 мВ (милливольт), хотя в разных клетках может быть различной: от −35 мВ до −90 мВ.
Важно учитывать, что в нервной системе электрические заряды представлены не электронами, как в обычных металлических проводах, а ионами — химическими частицами, имеющими электрический заряд. И вообще в водных растворах в виде электрического тока перемещаются не электроны, а ионы. Поэтому все электрические токи в клетках и окружающей их среде — это ионные токи.
Итак, изнутри клетка в покое заряжена отрицательно, а снаружи — положительно. Это свойственно всем живым клеткам, за исключением, разве что, эритроцитов, которые, наоборот, заряжены отрицательно снаружи. Если говорить конкретнее, то получается, что снаружи вокруг клетки будут преобладать положительные ионы (катионы Na + и K + ), а внутри — отрицательные ионы (анионы органических кислот, не способные свободно перемещаться через мембрану, как Na + и K + ).
Теперь нам всего лишь осталось объяснить, каким же образом всё получилось именно так. Хотя, конечно, неприятно сознавать, что все наши клетки кроме эритроцитов только снаружи выглядят положительными, а внутри они — отрицательные.
Сущность формирования потенциала покоя
Попробуем разобраться, откуда берётся электрический заряд нервных клеток, хотя их никто не трёт, как это делают физики в своих опытах с электрическими зарядами.
Здесь исследователя и студента поджидает одна из логических ловушек: внутренняя отрицательность клетки возникает не из-за появления лишних отрицательных частиц (анионов), а, наоборот, из-за потери некоторого количества положительных частиц (катионов)!
Так куда же деваются из клетки положительно заряженные частицы? Напомню, что это покинувшие клетку и скопившиеся снаружи ионы натрия — Na + — и калия — K + .
Главный секрет появления отрицательности внутри клетки
Сразу откроем этот секрет и скажем, что клетка лишается части своих положительных частиц и заряжается отрицательно за счёт двух процессов:
Эти два процесса нам и надо объяснить.
Первый этап создания внутренней отрицательности: обмен Na + на K +
Но ведь при обмене одного положительного заряда (Na + ) на другой такой же положительный заряд (K + ) никакого дефицита положительных зарядов в клетке возникать не может! Правильно. Но, тем не менее, из-за этого обмена в клетке остаётся очень мало ионов натрия, потому что они почти все ушли наружу. И в то же время клетка переполняется ионами калия, которые в неё накачали молекулярные насосы. Если бы мы могли попробовать на вкус цитоплазму клетки, мы бы заметили, что в результате работы насосов-обменников она превратилась из солёной в горько-солёно-кислую, потому что солёный вкус хлорида натрия сменился сложным вкусом довольно-таки концентрированного раствора хлорида калия. В клетке концентрация калия достигает 0,4 моль/л. Растворы хлорида калия в пределах 0,009–0,02 моль/л имеют сладкий вкус, 0,03–0,04 — горький, 0,05–0,1 — горько-солёный, а начиная с 0,2 и выше — сложный вкус, состоящий из солёного, горького и кислого [8].
Кстати, интересно, что клетка не рождается с готовым потенциалом покоя. Ей его ещё надо создать. Например, при дифференцировке и слиянии миобластов потенциал их мембраны изменяется от −10 до −70 мВ, т.е. их мембрана становится более отрицательной — поляризуется в процессе дифференцировки. А в экспериментах на мультипотентных мезенхимальных стромальных клетках костного мозга человека искусственная деполяризация, противодействующая потенциалу покоя и уменьшающая отрицательность клеток, даже ингибировала (угнетала) дифференцировку клеток [1].
- любовь клетки к калию (поэтому клетка насильно затаскивает его к себе);
- любовь калия к свободе (поэтому калий покидает захватившую его клетку).
Механизм насыщения клетки калием мы уже объяснили (это работа насосов-обменников), а механизм ухода калия из клетки объясним ниже, когда перейдём к описанию второго этапа создания внутриклеточной отрицательности. Итак, результат деятельности мембранных ионных насосов-обменников на первом этапе формирования потенциала покоя таков:
- Дефицит натрия (Na + ) в клетке.
- Избыток калия (K + ) в клетке.
- Появление на мембране слабого электрического потенциала (−10 мВ).
Можно сказать так: на первом этапе ионные насосы мембраны создают разность концентраций ионов, или градиент (перепад) концентрации, между внутриклеточной и внеклеточной средой.
Второй этап создания отрицательности: утечка ионов K + из клетки
Итак, что начинается в клетке после того, как с ионами поработают её мембранные натрий-калиевые насосы-обменники?
Из-за образовавшегося дефицита натрия внутри клетки этот ион при каждом удобном случае норовит устремиться внутрь: растворённые вещества всегда стремятся выровнять свою концентрацию во всём объёме раствора. Но это у натрия получается плохо, поскольку ионные натриевые каналы обычно закрыты и открываются только при определённых условиях: под воздействием специальных веществ (трансмиттеров) или при уменьшении отрицательности в клетке (деполяризации мембраны).
В то же время в клетке имеется избыток ионов калия по сравнению с наружной средой — потому что насосы мембраны насильно накачали его в клетку. И он, тоже стремясь уравнять свою концентрацию внутри и снаружи, норовит, напротив, выйти из клетки. И это у него получается!
Ионы калия K + покидают клетку под действием химического градиента их концентрации по разные стороны мембраны (мембрана значительно более проницаема для K + , чем для Na + ) и уносят с собой положительные заряды. Из-за этого внутри клетки нарастает отрицательность.
К + -каналы утечки постоянно открыты при нормальных значениях мембранного потенциала покоя и проявляют взрывы активности при сдвигах мембранного потенциала, которые длятся несколько минут и наблюдаются при всех значениях потенциала. Усиление К + -токов утечки ведёт к гиперполяризации мембраны, тогда как их подавление — к деполяризации. . Однако, существование канального механизма, ответственного за токи утечки, долгое время оставалось под вопросом. Только сейчас стало ясно, что калиевая утечка — это ток через специальные калиевые каналы.
Зефиров А.Л. и Ситдикова Г.Ф. Ионные каналы возбудимой клетки (структура, функция, патология) [7].
От химического — к электрическому
А теперь — ещё раз самое главное. Мы должны осознанно перейти от движения химических частиц к движению электрических зарядов.
Для этой составной части потенциала покоя есть даже специальное название — концентрационный потенциал [5]. Концентрационный потенциал — это часть потенциала покоя, созданная дефицитом положительных зарядов внутри клетки, образовавшимся за счёт утечки из неё положительных ионов калия.
Ну, а теперь немного физики, химии и математики для любителей точности.
Электрические силы связаны с химическими по уравнению Гольдмана. Его частным случаем является более простое уравнение Нернста, по формуле которого можно рассчитать трансмембранную диффузионную разность потенциалов на основе различной концентрации ионов одного вида по разные стороны мембраны. Так, зная концентрацию ионов калия снаружи и внутри клетки, можно рассчитать калиевый равновесный потенциал EK:

где Ек — равновесный потенциал, R — газовая постоянная, Т — абсолютная температура, F — постоянная Фарадея, К + внеш и K + внутр — концентрации ионов К + снаружи и внутри клетки, соответственно. По формуле видно, что для расчёта потенциала между собой сравниваются концентрации ионов одного вида — K + .
Более точно итоговая величина суммарного диффузионного потенциала, который создаётся утечкой нескольких видов ионов, рассчитывается по формуле Гольдмана-Ходжкина-Катца. В ней учтено, что потенциал покоя зависит от трех факторов: (1) полярности электрического заряда каждого иона; (2) проницаемости мембраны Р для каждого иона; (3) [концентраций соответствующих ионов] внутри (внутр) и снаружи мембраны (внеш). Для мембраны аксона кальмара в покое отношение проводимостей РK : PNa :PCl = 1 : 0,04 : 0,45 [5].

Заключение
Итак, поте нциал покоя состоит из двух частей:
Вот теперь мембранный потенциал покоя окончательно сформирован.
Читайте также: