
Хемотрофы это в биологии кратко
Обновлено: 06.07.2024
В хемотрофы или хемосинтетики - это группа организмов, которые для выживания используют восстановленные неорганические соединения в качестве сырья, откуда они получают энергию для последующего использования в дыхательном метаболизме.
Это свойство этих микроорганизмов получать энергию из очень простых соединений для образования сложных соединений также известно как хемосинтез, поэтому эти организмы иногда также называют хемосинтетиками.
Другой важной характеристикой является то, что эти микроорганизмы отличаются от остальных тем, что они растут в строго минеральных средах и без света, поэтому их иногда называют хемолиттрофами.
характеристики
Место обитания
Эти бактерии живут там, где проникает менее 1% солнечного света, то есть они процветают в темноте, почти всегда в присутствии кислорода.
Однако идеальным местом для развития хемосинтетических бактерий являются переходные слои между аэробными и анаэробными условиями.
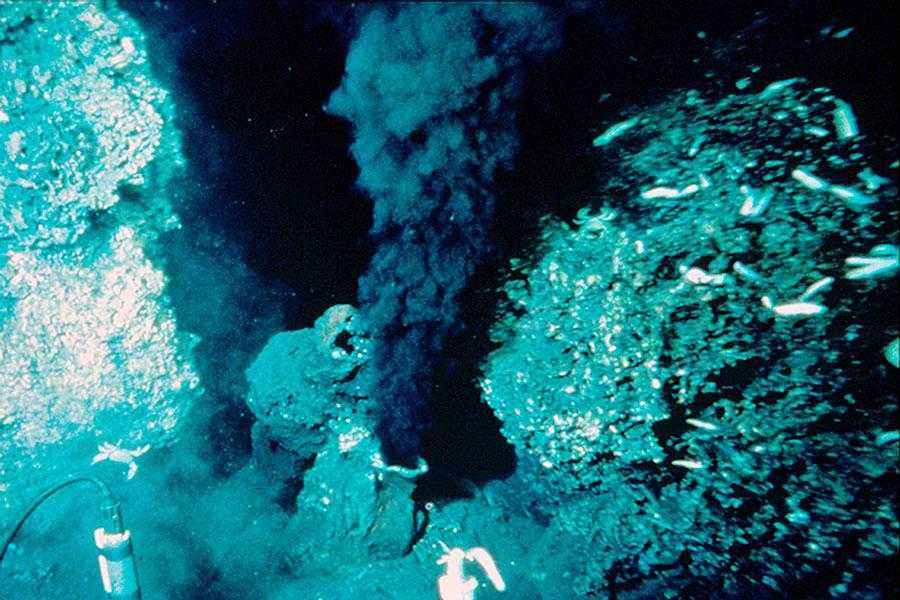
Чаще всего встречаются: глубокие отложения, окрестности подводных рельефов или подводные возвышенности, расположенные в средней части океанов, известные как срединно-океанические хребты.
Эти бактерии способны выжить в экстремальных условиях. На этих участках могут быть гидротермальные источники, из которых вытекает горячая вода или даже истечение магмы.
Функция в окружающей среде
Эти микроорганизмы необходимы для экосистемы, поскольку они превращают токсичные химические вещества, выходящие из этих отверстий, в пищу и энергию.
Вот почему хемосинтезирующие организмы играют фундаментальную роль в восстановлении минеральной пищи, а также спасают энергию, которая в противном случае была бы потеряна.
То есть они способствуют поддержанию трофической цепи или пищевой цепи.
Это означает, что они способствуют передаче питательных веществ через различные виды биологического сообщества, в котором каждый питается предыдущим и является пищей для следующего, что помогает поддерживать баланс экосистемы.
Эти бактерии также способствуют спасению или улучшению некоторых экологических сред, загрязненных в результате аварий. Например, в местах разлива нефти, то есть в этих случаях эти бактерии помогают обрабатывать токсичные отходы, превращая их в более безопасные соединения.
Классификация
Хемосинтетические или хемотрофные организмы подразделяются на хемоавтотрофов и хемогетеротрофов.
Хемоавтотрофы
Они используют CO2 как источник углерода, который ассимилируется в ходе цикла Кальвина и превращается в клеточные компоненты.
С другой стороны, они получают энергию от окисления восстановленных простых неорганических соединений, таких как: аммиак (NH3), дигидроген (H2), диоксид азота (NO2 – ), сероводород (H2S), сера (S), триоксид серы (S2ИЛИ3 – ) или ион железа (Fe2 + ).
То есть АТФ образуется в результате окислительного фосфорилирования во время окисления неорганического источника. Следовательно, они самодостаточны, им не нужно другое живое существо, чтобы выжить.
Хемогетеротрофы
В отличие от предыдущих, они получают энергию за счет окисления сложных восстановленных органических молекул, таких как глюкоза через гликолиз, триглицериды через бета-окисление и аминокислоты через окислительное дезаминирование. Таким образом они получают молекулы АТФ.
С другой стороны, хемогетеротрофные организмы не могут использовать CO.2 как источник углерода, как и хемоавтотрофы.
Типы хемотрофных бактерий
Бесцветные серные бактерии
Как следует из названия, это бактерии, окисляющие серу или ее восстановленные производные.
Эти бактерии строго аэробны и отвечают за преобразование сероводорода, образующегося при разложении органических веществ, в сульфат (SO4 -2 ), соединение, которое в конечном итоге будет использоваться растениями.
Сульфат подкисляет почву примерно до pH 2 из-за накопления протонов H. + образуется серная кислота.
Это делается путем внесения в почву порошковой серы, чтобы присутствующие специализированные бактерии (сульфобактерии) окисляли серу и, таким образом, уравновешивали pH почвы до значений, подходящих для сельского хозяйства.
Все хемолитрофные виды, окисляющие серу, являются грамотрицательными и принадлежат к типу Proteobacteria. Примером бактерий, окисляющих серу, являетсяAcidithiobacillus thiooxidans.
Некоторые бактерии могут накапливать элементарную серу (S 0 ) нерастворимые в виде гранул внутри ячейки, которые будут использоваться при исчерпании внешних источников серы.
Азотные бактерии
В этом случае бактерии окисляют восстановленные соединения азота. Есть два типа: нитрозирующие бактерии и нитрифицирующие бактерии.
Первые способны окислять аммиак (NH3), который образуется при разложении органических веществ, превращая их в нитриты (NO2), а последние превращают нитриты в нитраты (NO3 – ), соединения, используемые растениями.
Примером нитрозифицирующих бактерий является род Nitrosomonas, а в качестве нитрифицирующих бактерий - род Nitrobacter.
Железные бактерии
Эти бактерии являются ацидофильными, то есть им для выживания требуется кислый pH, поскольку при нейтральном или щелочном pH соединения двухвалентного железа окисляются самопроизвольно, без необходимости присутствия этих бактерий.
Следовательно, чтобы эти бактерии окисляли соединения двухвалентного железа (Fe 2+ ) на железо (Fe 3+ ) pH среды обязательно должен быть кислым.
Следует отметить, что железобактерии расходуют большую часть АТФ, образующегося в реакциях обратного переноса электронов, для получения необходимой восстанавливающей способности при фиксации CO.2.
Вот почему этим бактериям приходится окислять большое количество Fe. +2 чтобы иметь возможность развиваться, потому что в процессе окисления выделяется мало энергии.
Пример: бактерии Acidithiobacillus ferrooxidans он превращает карбонат железа, присутствующий в кислой воде, протекающей через угольные шахты, в оксид железа.
Все хемолиттрофные виды, окисляющие железо, являются грамотрицательными и принадлежат к типу Proteobacteria.
С другой стороны, все вещества, окисляющие железо, также способны окислять серу, но не наоборот.
Водородные бактерии
Эти бактерии используют молекулярный водород в качестве источника энергии для производства органических веществ и используют CO.2 как источник углерода. Эти бактерии являются факультативными хемоавтотрофами.
В основном они встречаются в вулканах. Никель необходим в среде его обитания, поскольку все гидрогеназы содержат это соединение в качестве металлического кофактора. У этих бактерий отсутствует внутренняя мембрана.
В своем метаболизме водород включается в гидрогеназу плазматической мембраны, перемещая протоны наружу.
Таким образом, внешний водород проходит внутрь, действуя как внутренняя гидрогеназа, превращая НАД. + в НАДН, который вместе с углекислым газом и АТФ переходят в цикл Кальвина.
Бактерии Hydrogenomonas Они также могут использовать ряд органических соединений в качестве источников энергии.
Хемотрофы – организмы, которые получают энергию главным образом из углекислого газа и других неорганических химических веществ через процесс, называемый хемосинтезом. Хемосинтез осуществляется хемотрофами путем окисления доноров электронов из окружающей среды. Хемотрофы могут быть как автотрофными (хемоавтотрофы), так и гетеротрофными (хемогетеротрофы).
Хемоавтотрофы – микроорганизмы, синтезирующие органические вещества из неорганических с помощью хемосинтеза. Хемосинтез – это процесс, посредством которого некоторые бактерии и археи, преобразовывают химическую энергию в питательные вещества. Они способны использовать в качестве восстановителей неорганические соединения, такие как сероводород, сера, аммоний и железо, а также синтезировать органические соединения из углекислого газа. Хемоавтотрофы встречаются в экстремальной среде обитания, например в глубоководных источниках, куда не проникает солнечный свет. К ним относятся метаногены, галофилы, нитрификаторы, термоацидофилы, сероокисляющие бактерии и другие экстремофилы.
Хемогетеротрофы являются гетеротрофами (организмы, которые не могут синтезировать органические вещества из неорганических путём фотосинтеза или хемосинтеза). Хемогетеротрофы подразделяются на хемолитогетеротрофов и хемоорганогетеротрофов:
Хемосинтезирующие бактерии (хемолитоавтотрофы) — бактерии, использующие диоксид углерода в качестве единственного источника углерода. Получают энергию в результате окислительно-восстановительных реакций, донором электронов являются неорганические соединения. К хемолитоавтотрофам относят представителей родов Nitrosospina, Nitrosococcus, Thiobacillus и других.
Связанные понятия
Фототрофы (др.-греч. φῶς, φωτός = свет, τροϕή = питание) — это организмы, которые используют свет для получения энергии. Они используют энергию света для поддержания различных метаболических процессов. Существует распространенное заблуждение, что фототрофы должны обязательно фотосинтезировать. Многие, хотя далеко не все, действительно фотосинтезируют: они используют энергию света, чтобы преобразовывать углекислый газ в органический материал, который служит для построения их тела, или в качестве источника.
Миксотро́фы (от др.-греч. μῖξις — смешение и τροφή — пища, питание) — организмы, способные использовать различные источники углерода и доноры электронов. Миксотрофы могут быть одновременно фототрофами и хемотрофами, литотрофами и органотрофами. Миксотрофами являются представители как прокариот, так и эукариот.Примером организма с миксотрофным получением углерода и энергии является бактерия Paracoccus pantotrophus из семейства Rhodobacteraceae — хемооргано-гетеротроф, также способная существовать.
Серобактерии (Тиобактерии) — весьма разнородная группа прокариотов, окисляющих восстановленные соединения серы.
Денитрификация (восстановление нитрата) — сумма микробиологических процессов восстановления нитратов до нитритов и далее до газообразных оксидов и молекулярного азота. В результате их азот возвращается в атмосферу и становится недоступным большинству организмов. Осуществляется только прокариотами (причём как бактериями, так и археями) в анаэробных условиях и связана с получением ими энергии.
Пурпурные бактерии (Purple bacteria) — разнородная группа фотосинтезирующих протеобактерий, обитающих в солёных и пресных водах. Пурпурные бактерии относятся к классам альфа-, бета-, и гамма-протеобактерий.
Связывание углерода — общее название совокупности процессов, при которых углекислый газ CO2 преобразуется в органические вещества. Такие процессы используют автотрофы, то есть организмы, которые сами вырабатывают необходимые для себя органические вещества. В частности, процесс связывания углерода является составной частью фотосинтеза.
Анаэробное дыхание — биохимический процесс окисления органических субстратов или молекулярного водорода с использованием в дыхательной ЭТЦ в качестве конечного акцептора электронов вместо O2 других окислителей неорганической или органической природы. Как и в случае аэробного дыхания, выделяющаяся в ходе реакции свободная энергия запасается в виде трансмембранного протонного потенциала, использующегося АТФ-синтазой для синтеза АТФ.
Хемосинтез — способ автотрофного питания, при котором источником энергии для синтеза органических веществ из CO2 служат реакции окисления неорганических соединений. Подобный вариант получения энергии используется только бактериями или археями. Это явление было открыто в 1889 году русским учёным С. Н. Виноградским. Микроорганизмы, способные к хемосинтезу, Виноградский называл аноргоксиданты. Название хемосинтез ввёл немецкий химик и ботаник Вильгельм Пфеффер в 1897 году.
Фикобили́ны (от греч. φύκος — водоросли и лат. bilis — желчь) — тетрапиррольные пигменты (билины) красных водорослей, криптофит и цианобактерий (синезелёных водорослей). Фикобилины являются хромофорной группой фикобилипротеинов — кислых водорастворимых глобулярных хромопротеинов светособирающего комплекса водорослей. Отдельными молекулами фикобилины, как правило, не представлены, а образуют комплексы с белками — фикобилипротеиды (хромопротеиды).
Автотро́фы (др.-греч. αὐτός — сам + τροφή — пища) — организмы, синтезирующие органические вещества из неорганических. Автотрофы составляют первый ярус в пищевой пирамиде (первые звенья пищевых цепей). Именно они являются первичными продуцентами органического вещества в биосфере, обеспечивая пищей гетеротрофов. Следует отметить, что иногда резкой границы между автотрофами и гетеротрофами провести не удаётся. Например, одноклеточная водоросль эвглена зелёная на свету является автотрофом, а в темноте.
Зелёные несе́рные бакте́рии, или зелёные ни́тчатые серобакте́рии, — филогенетически однородная группа факультативно анаэробных фотогетеротрофных бактерий, осуществляющих аноксигенный фотосинтез, использующих H2S, H2, сахара, аминокислоты и органические кислоты в качестве доноров электронов. В отличие от зелёных серобактерий, зелёные несерные бактерии при окислении сероводорода откладывают элементарную серу вне клеток, за что и получили своё название. Следует отметить, что зелёные несерные бактерии.
Обра́тный цикл Кре́бса, также известный как обра́тный цикл трикарбо́новых кисло́т, или цикл А́рнона — последовательность химических реакций, которую некоторые бактерии используют для синтеза органических соединений из диоксида углерода и воды.
Биоремедиация — комплекс методов очистки вод, грунтов и атмосферы с использованием метаболического потенциала биологических объектов — растений, грибов, насекомых, червей и других организмов.
Нитрогеназа (КФ 1.18.6.1) — комплекс ферментов (мультифермент), осуществляющий процесс фиксации атмосферного азота. Широко распространён у бактерий и архей, в то время как все эукариоты его лишены.
Нитрификация — микробиологический процесс окисления аммиака до азотистой кислоты или её самой далее до азотной кислоты, что связано либо с получением энергии (хемосинтез, автотрофная нитрификация), либо с защитой от активных форм кислорода, образующихся при разложении пероксида водорода (гетеротрофная нитрификация).
Зелёные серобактерии (лат. Chlorobiaceae) — семейство облигатно анаэробных (более строгих, чем пурпурные бактерии, в присутствии O2 не растут) фотолитоавтотрофных грамотрицательных бактерий, использующих сероводород (H2S), водород (H2) и элементарную серу (S0) в качестве доноров электронов. По происхождению они принадлежат к надтипу Bacteroidetes-Chlorobi, однако неоднородны и потому их классифицируют как отдельный тип.
Планктомице́ты (лат. Planctomycetes) — тип грамотрицательных бактерий, отличающихся уникальной клеточной структурой, а именно наличием сложной системы замкнутых мембран. В частности, у некоторых представителей нуклеоид находится в ядерном тельце, окружённом двойной мембраной. Некоторые виды осуществляют анаммокс — процесс анаэробного окисления аммиака, в ходе которого образуется элементарный азот).
Метаногены (Methanogens) — это археи, которые образуют метан как побочный продукт метаболизма в бескислородных условиях. Широко распространены в заболоченных территориях, где образуют метан (болотный газ) и в кишечниках жвачных млекопитающих и человека, и отвечают за метеоризм. В глубинах океанов биосинтез метана археями обычно пространственно располагается в местах выхода сульфатов. Некоторые являются экстремофилами и обитают в горячих источниках и на больших глубинах, а также на скалах и на глубине.
Анаммо́кс (сокр. от англ. anaerobic ammonium oxidation — анаэробное окисление аммония) — один из ключевых микробных процессов в круговороте азота. Бактерии, осуществляющие этот процесс, были открыты в 1999 году, и в своё время описание этого процесса стало большим сюрпризом для научного сообщества. Уравнение процесса.
Оксидазы — окислительные ферменты класса оксидоредуктаз. В настоящее время найдено очень много разнообразных окислительных ферментов, как растительного, так и животного происхождения. В живых клетках оксидазы служат катализаторами окислительно-восстановительных реакций и классифицируются на металлоферменты и флавопротеиды.
Бактериохлорофи́ллы — гетерогенная группа фотосинтетических тетрапиррольных пигментов, которые синтезируются различными аноксигенными фототрофными бактериями, осуществляющими фотосинтез без выделения кислорода.
Круговорот азота — биогеохимический цикл азота. Большая его часть обусловлена действием живых существ. Очень большую роль в круговороте играют почвенные микроорганизмы, обеспечивающие азотистый обмен почвы — круговорот в почве азота, который присутствует там в виде простого вещества (газа — N2) и ионов: нитритов (NO2-), нитратов (NO3-) и аммония (NH4+). Концентрации этих ионов отражают состояние почвенных сообществ, поскольку на эти показатели влияет состояние биоты (растений, микрофлоры), состояние.
Анаэробное окисление метана — процесс окисления метана до углекислого газа, производимый некультивируемыми (англ. VBNC) археями групп ANME-1, ANME-2 и ANME-3, близкими к Methanosarcinales при отсутствии в среде молекулярного кислорода. Биохимия и распространённость процесса в природе изучены пока недостаточно.
Факультативные анаэробы — организмы, энергетические циклы которых проходят по анаэробному пути, но способные существовать при доступе кислорода, в отличие от облигатных анаэробов, для которых кислород губителен.
Аэро́бы (от греч. αηρ — воздух и βιοζ — жизнь) — организмы, которые нуждаются в свободном молекулярном кислороде для процессов синтеза энергии, в отличие от анаэробов. К аэробам относятся подавляющее большинство животных, все растения, а также значительная часть микроорганизмов.
Фикобилисо́мы — светособирающие органеллы для фотосистемы II у цианобактерий, красных водорослей и глаукофитов. Стандартные фикобилисомы отсутствуют у криптофитовых и тех представителей прохлорофитовых, у которых имеются фикобилипротеины. У криптофитовых фикобилипротеины находятся во внутритилакоидном пространстве.
Биомолекулы — это органические вещества, которые синтезируются живыми организмами. В состав биомолекул включают белки, полисахариды, нуклеиновые кислоты, а также более мелкие компоненты обмена веществ. Биомолекулы состоят из атомов углерода, водорода, азота, кислорода, а также фосфора и серы. Другие атомы входят в состав биологически значимых веществ значительно реже.
Гамма-протеобактерии (лат. Gammaproteobacteria) — класс грамотрицательных бактерий типа протеобактерий, в который входят некоторые группы, важные в медицинском, экологическом и научном плане. Сюда же относят чрезвычайно большое количество патогенных микроорганизмов.
Хромофо́ры (др.-греч. χρῶμα — цвет и φέρω — несу) — ненасыщенные группы атомов, обуславливающие цвет химического соединения. В то же время поглощающие электромагнитное излучение независимо от наличия окраски. Так, карбонильная группа C=O является хромофором, поглощающим в области 280 нм, в то же время кетоны, содержащие С=O – бесцветные вещества. Хромофорная теория возникновения окраски была предложена в 1878 г. немецким учёным Виттом. К хромофорам относят азогруппу —N=N—, нитрогруппу —NO2, нитрозогруппу.
Вспомогательные пигменты — светопоглощающие соединения фотосинтезирующих организмов, которые работают в сочетании с хлорофиллом а, выполняя светособирающую или светозащитную функции. К ним относятся различные формы хлорофилла, например хлорофилл b в зеленых водорослях и светособирающих антеннах высших растений или хлорофиллы с или d у других водорослей. Кроме того есть много других вспомогательных пигментов таких как каротиноиды или фикобилипротеины, которые также поглощают свет и передают его энергию.
Метаногенез, биосинтез метана — процесс образования метана анаэробными археями, сопряжённый с получением ими энергии. Существует три типа метаногенеза.
Кренархео́ты (лат. Crenarchaeota, от др.-греч. κρήνη — ручей, ἀρχαῖος — древний) — тип в составе домена архей, включающий единственный класс Thermoprotei Reysenbach 2002. Тип выделен на основании анализа последовательностей 16S рРНК.
Карбоксисо́мы (полиэдральные тела) — микрокомпартменты в клетках бактерий, содержащие фиксирующие углерод ферменты. Они представляют собой многогранные однослойные белковые тела полиэдрической формы от 80 до 140 нанометров в диаметре. Они являются основной частью механизма концентрирования CO2, что помогает преодолеть неэффективность рибулозодифосфаткарбоксилазы (Рубиско) — главного фермента, лимитирующего скорость фиксации углерода в цикле Кальвина. Эти органеллы обнаружены во всех цианобактериях.
Гидрогеносомы трихомонад (наиболее изучены среди гидрогеносом-содержащих микроорганизмов) выделяют молекулярный водород, ацетат, углекислый газ и производит АТФ, комбинируя действие ферментов пируват:ферредоксин-оксидоредуктазы, гидрогеназы, ацетат-сукцинат-КоА-трансферазы и сукцинаттиокиназы. В них также содержатся супероксиддисмутаза, малатдегидрогеназа (декарбоксилирующая), ферредоксин, аденилаткиназа и НАДH:ферредоксин-оксидоредуктаза. Полагают, что эти органеллы произошли от эндосимбиотических.
Га́зовые вези́кулы (англ. Gas vesicle) — заполненные газом микрокомпартменты, обеспечивающие повышенную плавучесть клеток у примерно 150 видов планктонных бактерий и архей. Особенно часто газовые вакуоли встречаются у цианобактерий. Мембрана, ограничивающая газовую везикулу, имеет белковую природу, а внутри газовой везикулы находится атмосферный воздух. Газовые везикулы располагаются в цитоплазме поодиночке или образуют сотовидные скопления, которые иногда неправильно называют газовыми вакуолями.
Гетероцисты — дифференцированные клетки нитчатых цианобактерий, осуществляющие азотфиксацию. При недостатке соединений азота в среде они появляются регулярно вдоль трихомы из вегетативных клеток и акинет. Цианобактерии — фототрофы, осуществляющие оксигенный фотосинтез, однако кислород, атмосферный и выделяемый при фотосинтезе, ингибирует фермент нитрогеназу, необходимую для азотфиксации, поэтому у нитчатых цианобактерий в процессе эволюции возникли специализированные клетки для азотфиксации.
Хлоросо́мы (англ. Chlorosome от др.-греч. χλωρός — зелёный) — обогащённые липидами везикулы зелёных серныx бактерий и нитчатых аноксигенных фототрофных бактерий, локализованные в цитоплазме и связанные с клеточной мембраной кристаллической базальной пластинкой. Внутри хлоросомы находятся пучки палочковидных структур, содержащих молекулы бактериохлорофиллов c, d или e. Таким образом, в хлоросомах собраны светособирающие системы.
Ауксотрофы — организмы, которые не способны синтезировать определенное органическое соединение, необходимое для роста этого организма. Ауксотрофия — характеристика подобных организмов, этот термин противоположен прототрофии.
Азотоба́ктер (лат. Azotobacter) — род бактерий, живущих в почве и способных в результате процесса азотфиксации переводить газообразный азот в растворимую форму, доступную для усваивания растениями.
Под эволюцией фотосинтеза понимают исторический путь происхождения и последующего развития фотосинтеза или последовательное становление и изменение процесса преобразования солнечной энергии в химическую для синтеза сахаров из углекислого газа, с выделением кислорода в качестве побочного продукта.
Восстановительный пентозофосфатный цикл, или цикл Кальвина, — серия биохимических реакций, осуществляемая при фотосинтезе растениями (в строме хлоропластов), цианобактериями, прохлорофитами и пурпурными бактериями, а также многими бактериями-хемосинтетиками, является наиболее распространённым из механизмов автотрофной фиксации CO2. Цикл Кальвина назван в честь американского биохимика Мелвина Кальвина (1911—1997). Часто используются альтернативные названия, указывающие на роль коллег Кальвина в открытии.
Оксидоредукта́зы (КФ1) — отдельный класс ферментов, катализирующих лежащие в основе биологического окисления реакции, сопровождающиеся переносом электронов с одной молекулы (восстановителя — акцептора протонов или донора электронов) на другую (окислитель — донор протонов или акцептор электронов).
Хламидомона́да (лат. Chlamydómonas) — род одноклеточных зелёных водорослей из семейства Хламидомонадовые (Chlamydomonadaceae).

Хемотрофы организмы, которые получают энергия посредством окисление из доноры электронов в их среде. [1] Эти молекулы могут быть органический (хемоорганотрофы) или же неорганический (хемолитотрофы). Обозначение хемотрофа в отличие от фототрофы, которые используют солнечную энергию. Хемотрофы могут быть либо автотрофный или же гетеротрофный. Хемотрофы встречаются на дне океана, где солнечный свет не может достичь их из-за большой глубины и всех уровней воды между ними и солнцем. Они эволюционировали, чтобы не зависеть от солнечной энергии. Дно океана часто содержит подводные вулканы, которые могут обеспечивать тепло вместо солнечного тепла.
Содержание
Хемоавтотроф
Хемоавтотрофы (или хемотрофный автотроф) (Греческий: Химиотерапия (χημεία) = химическая, авто (εαυτός) = self, troph (τροφή) = питание), в дополнение к получению энергии из химические реакции, синтезировать все необходимые органические соединения из углекислый газ. Хемоавтотрофы могут использовать неорганические источники электронов, такие как сероводород, элементаль сера, двухвалентное железо, молекулярный водород, и аммиак или органических источников. Большинство хемоавтотрофов экстремофилы, бактерии или же археи которые живут во враждебной среде (например, глубоководные жерла) и являются первичные производители в таком экосистемы. Хемоавтотрофы обычно делятся на несколько групп: метаногены, окислители серы и редукторы, нитрификаторы, анаммокс бактерии и термоацидофилы. Примером одного из этих прокариот может быть Сульфолобус. Хемолитотрофный рост может быть очень быстрым, например, Hydrogenovibrio crunogenus с время удвоения около часа. [2] [3]
Период, термин "хемосинтез", изобретенный в 1897 г. Вильгельм Пфеффер, первоначально было определено как производство энергии путем окисления неорганических веществ в сочетании с автотрофия- то, что сегодня назвали бы хемолитоавтотрофией. Позже этот термин будет включать также хемоорганоавтотрофию, то есть его можно рассматривать как синоним химиоавтотрофия. [4] [5]
Хемогетеротроф
Бактерии, окисляющие железо и марганец
В глубоких океанах бактерии, окисляющие железо, получают свои потребности в энергии за счет окисления двухвалентного железа (Fe 2+ ) в трехвалентное железо (Fe 3+ ). Электрон, законсервированный из этой реакции, уменьшает дыхательная цепь и, таким образом, может быть использован в синтезе АТФ вперед электронный транспорт или же НАДН обратным электронный транспорт, заменяя или дополняя традиционные фототрофизм.
- Как правило, бактерии, окисляющие железо, могут существовать только в областях с высокими концентрациями двухвалентного железа, таких как новые слои лавы или области гидротермальной активности. Большая часть океана лишена двухвалентного железа из-за окислительного эффекта растворенного кислорода в воде и склонности бактерий поглощать железо.
- Слои лавы снабжают бактерии двухвалентным железом прямо из мантии Земли, но только новообразованными. огненный горные породы имеют достаточно высокий уровень двухвалентного железа. Кроме того, поскольку для реакции необходим кислород, эти бактерии гораздо чаще встречаются в верхних слоях океана, где кислорода больше.
- Пока неизвестно, как именно железобактерии извлекают железо из породы. Принято считать, что существует некий механизм, разъедающий породу, возможно, через специализированные ферменты или соединения, которые выводят на поверхность больше FeO. Давно обсуждается, какая часть выветривания скалы происходит из-за биотические компоненты и сколько можно отнести к абиотические компоненты.
- Гидротермальные источники также выбрасывают большое количество растворенного железа в глубину океана, позволяя бактериям выжить. Кроме того, высокий температурный градиент вокруг вентиляционных систем означает, что могут сосуществовать самые разные бактерии, каждая из которых имеет свою специализированную температурную нишу.
- Независимо от каталитический При использовании метода хемоавтотрофные бактерии являются важным, но часто упускаемым из виду источником пищи для глубоководных экосистем, которые иначе получают ограниченный солнечный свет и органические питательные вещества.
Марганец- окисляющие бактерии также используют вулканические породы таким же образом; окислением марганца марганца (Mn 2+ ) в марганец (Mn 4+ ) марганец. Марганца гораздо меньше, чем в океанической коре железа, но бактериям гораздо легче извлечь его из магматического стекла. Кроме того, каждое окисление марганца отдает ячейке два электрона по сравнению с одним при каждом окислении железа, хотя количество АТФ или же НАДН которые могут быть синтезированы в сочетании с этими реакциями, зависит от pH и термодинамики конкретной реакции с точки зрения того, сколько Свободная энергия Гиббса изменения, происходящие во время реакций окисления, по сравнению с изменением энергии, необходимым для образования АТФ или же НАДНВсе они зависят от концентрации, pH и т. д. Многое еще остается неизвестным о марганцево-окисляющих бактериях, поскольку они не культивировались и не документировались в значительной степени.
Читайте также: