
Генетическая структура популяции кратко
Обновлено: 30.06.2024
Одной из важных отраслей генетики является генетика популяций. Эта отрасль науки посвящена изучению генетической структуры природных популяций, а также генетических процессов, происходящих в ней. Она играет исключительно важную роль для развития эволюционной теории. Благодаря достижениям генетики популяций установлено, что популяция это не просто единица вида, но и единица эволюции.
Основоположником работ в этом направлении первой половине $XX$ века ($20$-е годы) стал С.С. Четвериков, русский генетик. Также, весомый вклад в развитие этой науки внесли:
- С.М. Гершинзон,
- Н.П. Дубинин,
- Д.Д.Ромашов,
- Р.Райт,
- Р.Фишер,
- Ф.Г.Добржанский.
Что же такое популяция? Современное определение этого понятия выглядит так:
Закон Харди-Вайнберга
Так как популяция состоит из особей одного вида, то все они обладают одинаковым набором генов. Однако любой ген может быть представлен разными аллелями, количество которых может быть значительным. Значит, популяция – это совокупность неодинаковых в генетическом отношении особей, которые отличаются разными состояниями свойственных им признаков. В природе разные популяции одного и того же вида могут отличаться частотами встречаемости определенных аллельных генов.
Исследование закономерностей, которые распределяют аллели в популяциях, является важной задачей популяционной генетики. Над этой проблемой работали немецкий ученый В. Вайнберг и английский исследователь Г. Харди. Они сделали вывод, что у достаточно многочисленной популяции, особи которой свободно скрещиваются между собой свободно, и без влияния какого-либо внешнего фактора на определенные сочетания аллелей, новые мутации не возникают , и обмен генетической информацией с другими популяциями в результате миграции особей в другие популяции не происходит, соотношение аллелей стабилизируется на протяжении нескольких поколений, затем на длительное время остается постоянным.
Готовые работы на аналогичную тему
Закономерность, установленная Г. Харди и В. Вайнбергом, говорит о том, что при условии постоянства внешних и внутренних факторов частоты встречаемости аллелей у достаточно многочисленной популяции, которая изолированна от других, остается достаточно устойчивым на протяжении длительного периода.
Ученые предложили формулу, с помощью которой можно описать распределение частот в панмиксической популяции по одной паре аллельных генов ($A – a$). Эту формулу используют в качестве простейшей модели, служащей основой для проведения более сложных популяционно-генетических исследований.
$p^2AA + 2pqAa + q^2aa = 1$
Уравнение Харди-Вайнберга
- $q$ - частота встречаемости рецессивного гена;
- $p$ - частота встречаемости доминантного гена;
- $q^2$ - частоста встречаемости генотипа $aa$;
- $p^2$ - частоста встречаемости генотипа $AA$;
- $2pq$ - частота встречаемости генотипа $Aa$.
Исследования показали, что только при отсутствии свободного скрещивания, можно существенно повлиять на распределение частот встречаемости генотипов потомков.
Кроме того на изменения частот могут повлиять интенсивные миграции особей из между популяциями. Это приводит к активному обмену разными аллелями.
Мутации, происходящие в популяциях, также дают отклонение в показателях частот встречаемости аллелей.
Дрейф генов
Среди главных причин изменений генетической структуры популяций можно выделить дрейф генов.
Дрейф генов – это случайное ненаправленное изменение частоты встречаемости в популяциях аллелей.
Очень четко это явление проявляется в популяциях с немногочисленными особями, благодаря существующему ограничению свободного спаривания во время размножения. Но чем больше численность популяции, тем меньшее влияние на изменение частот встречаемости аллелей имеет дрейф генов.
Генетическая структура популяций
Популяции между собой отличаются частотой различных сочетаний аллельных генов.
Совокупность всех генов и их аллелей особей определенной популяции именуется генофондом.
Особое значение в генетике имеют рецессивные мутации. Рецессивные признаки распространяются в популяции в гетерозиготном состоянии, в фенотипе не проявляются. Но со временем увеличивается вероятность образования гомозигот по рецессивному признаку. Тогда эти признаки проявляются в фенотипе. Если признак окажется полезным или нейтральным, он сохранится, если вредным – его носители просто погибнут. Поэтому рецессивные мутации называют резервом наследственной изменчивости популяций.
Генетика популяций (лекция) - раздел Биотехнологии, Формировать умения использовать генетическую символику при решении генетических задач Популяция – Совокупность Особей Одного Вида, Занимающих Определённую Территор.
Популяция – совокупность особей одного вида, занимающих определённую территорию, свободно скрещивающихся друг с другом, имеющих общее происхождение, определённую генетическую структуру и в той или в иной степени изолированных от других таких же совокупностей.
Популяция – совокупность особей одного вида, занимающих определённую территорию, свободно скрещивающихся друг с другом, имеющих общее происхождение, определённую генетическую структуру и в той или в иной степени изолированных от других таких же совокупностей.
Каждая популяция обладает генофондом и генетической структурой.
1. Генофонд – совокупность генотипов всех особей популяции.
2. Генетическая структура популяции – соотношение в ней различных генотипов и аллелей.
Изучением генетической структуры, закономерностей наследования признаков, динамикой популяции заОсновные понятия популяционной генетики: частота аллелей и частота генотипов.
1. Частота аллели равна доли данной аллели к общему числу аллелей в популяции
2. Частота генотипа равна доли данного генотипа к общему числу генотипов в популяции
нимается популяционная генетика.
Основные понятия популяционной генетики: частота аллелей и частота генотипов.
1. Частота аллели равна доли данной аллели к общему числу аллелей в популяции
2. Частота генотипа равна доли данного генотипа к общему числу генотипов в популяции
Большое влияние на генетическую структуру популяций оказывает способ размножение.
Популяции самоопыляющихся растений и перекрёстноопыляющихся растений резко отличаются друг от друга. В самоопыляющейся популяции с каждым поколением число гомозигот увеличивается, число гетерозигот уменьшается, следовательно постепенно распадается на две генотипические разнородные группы – чистые линии (гомозиготы доминантные и гомозиготы рецессивные).
В популяциях большинства растений и животных, размножающихся половым путём, осуществляется свободное скрещивание, обеспечивающее равномерную встречаемость гамет. Популяцию, в которой наблюдается равновероятная встреча гамет называется панмиктической.
Генетическая структура панмиктической популяции подчиняется закону Харди – Вайнберга.
Закон Харди – Вайнберга(1908 г) :
В идеальной популяции из поколения в поколение сохраняется строго определённое соотношение частот доминантных и рецессивных генов (p + q =1), а также соотношение частот генотипических классов (p² +2pq + q² = 1)
Где p– частота встречаемости доминантного гена А
q - частота встречаемости рецессивного гена а
p² - частота встречаемости гомозигот по доминанте (АА)
2 pq-частота встречаемости гетерозигот (Аа)
q² -частота встречаемости гомозигот по рецессиву (аа)
Условия существования идеальной популяции:
· Популяция состоит из большого числа генов
· Особи в популяции скрещиваются свободно (панмиксия)
· Отсутствует естественный отбор
· Популяция полностью изолирована от других популяций
У человека выделяют три типа популяций
Ø Изоляты – это эндогенные популяции. 1,5 тыс. (труднодоступные районы близкородственные браки больше 90%)
Ø Демы – небольшие популяции от 1,5 тыс. до 4 тыс. (близкородственные браки больше 80%)
Ø Большие популяции(панмиктические) – популяции с населением крупных городов
Генетическая структура популяций. Типы популяций
На всех уровнях организации живой материи (молекулярном, хромосомном, клеточном, организменном) действуют одинаковые, механизмы и законы наследственности и есть факторы, нарушающие эти закономерности и приводящие к изменениям признаков и свойств организмов. Но все виды на Земле существуют в форме не разрозненных особей, а их совокупностей. Группа, или совокупность особей, относящихся к одному виду, заселяющих определенное пространство и имеющих сходные механизмы адаптации к условиям обитания, называется популяцией. Изучением генетических процессов на уровне популяции занимается популяционная генетика.
Генетическая структура популяции самоопыляющихся организмов (сем. злаковых — ячмень, пшеница; сем. бобовых) характеризуется известной степенью гомозиготности особей по ряду генов.Самоопыление, приводя к гомозиготации особей, вовсе не делает популяцию генетически однородной. При самоопылении особей, гетерозиготных по одной паре аллелей, например Аа, в ряду поколений будет постоянно уменьшаться число гетерозигот, по при этом выщепятся по крайней мере две генетически неодинаковые гомозиготные чистые линии: АА и аа. Если учесть, что гомозиготация идет не по одной, а по многим парам аллелей, легко предположить, сколько чистых линий с различной комбинацией гомозигот по каждой паре аллелей может образоваться в популяции. Например, гомозиготация по двум парам аллелей приведет к формированию четырех чистых линий: ААВВ, ААвв, ааВВ и аавв.
Генетическая структура популяции – соотношение в популяциях различных генов и аллелей. Она определяется богатством генофонда популяции (совокупность генов всех особей популяции).Любая популяция представляет собой непрерывный поток поколений благодаря обмену генами, который происходит в результате скрещивания особей друг с другом. Признаки, появившиеся в ходе независимого комбинирования генов, определяют формирование фенотипа организмов и обусловливают изменчивость в популяции. В ходе естественного отбора адаптивные фенотипы сохраняются, а неадаптивные исчезают. Так формируется генетическая реакция всей популяции, которая определяет выживание данного вида. Только те особи популяции, которые выжили и оставили потомство, вносят вклад в будущее своего вида.
Популяция включает огромное количество разнообразных генов, которые образуют ее генофонд. Каждый ген может существовать в нескольких формахназываемых аллелями. Число особей в конкретной популяции, несущих определенный аллель, определяет частоту данного аллеля.
Причиной того, что отдельно взятый генотип не может служить адекватной еденицей процесса эволюции, является то, что его генотип остается неизменным на протяжении всей его жизни, а время жизни организма ограничено. С другой стороны, популяция представляет собой непрерывный ряд поколений, а генетическая структура популяции может изменяться, т.е. эволюционировать, от поколения к поколению. Непрерывность существования популяции обеспечивается механизмом биологической наследственности.
При изучении процесса эволюции важное значение имеет представление о генофонде. Генофондом называется совокупность генотипов всех особей популяции. Для диплоидных организмов генофонд популяции, насчитывает N особей, состоящих их 2N гаплоидных генотипов. Таким образом, генофонд популяции из N особей включает в себя N пар гомологичных хромосом и 2N аллелей каждого локуса (4N, т.к. у диплоидного организма 2 локуса). Исключение составляют половые хромосомы и сцепленные с полом гены, представленные в каждом гетерогаметном организме в одном экземпляре.
Непосредственно мы наблюдаем лишь фенотипы, а не генотипы или гены. Изменчивость генофонда может быть описана либо частотами генов, либо частотами генотипов. Если мы знаем соотношение между генотипами и соответствующими им фенотипами, то по частотам наблюдаемых фенотипов мы можем рассчитать частоты соответствующих генотипов.
Частоты аллелей можно рассчитать по частотам генотипов, учитывая, что в гомозиготах содержится по два одинаковых аллеля, а в гетерозиготах – по одному аллелю каждого типа. Таким образом, что бы получить частоту аллелей каждого типа, нужно к частоте индивидуумов, гомозиготных по данному аллелю, прибавить половину частоты гетерозигот по этому аллелю. Если частоты генотипов представить как: гомозиготных (АА) – D, (аа) – R, гетерозиготного (Аа) – H, то частоты аллелей считаются как:
p = D + 1/2 H
q = R + 1/2 H
Закон Харди — Вайнберга . свидетельствует о том, что наследование как таковое не меняет частоты аллелей в популяции.
Если обозначить частоту аллели А через р, а частоту аллели а через q, то при наличии по данному локусу только двух аллелей в популяции: рА + qa = 1. Соотношение генотипов в таком случае будет: (рА -- qa)2 = р2АА + 2pqAa -- q2aa= 1, в чем легко убедиться, воспользовавшись решеткой Пеннета:
Если вам нужна помощь в написании работы, то рекомендуем обратиться к профессионалам. Более 70 000 авторов готовы помочь вам прямо сейчас. Бесплатные корректировки и доработки. Узнайте стоимость своей работы.
Если в популяции для данного гена присутствуют три аллели с частотами р, q и г, то частоты генотипов также соответствуют формуле биномиального распределения:
(р + q + г)2 = р2 + q2 + г2 + 2pq + 2pr + 2qr = 1
и т. д. при большем числе аллелей.
Представим, что аллели А и а. встречаются с частотами 0,5, тогда в fi частоты генотипов будут:
0,25 АА 0,25Ла
Таким образом, 0,25АА + 0,50Ла + 0,25аа = 1. Нетрудно видеть, что при этом сохраняется исходная частота аллелей: 0,5А и 0,5а. В следующем поколении получают то же соотношение генотипов, как и в последующих поколениях. Пользуясь этим приемом и принимая иные значения р и q, легко убедиться, что уже в первом поколении в популяции устанавливается равновесие, сохраняющееся и во всех последующих.
Эта закономерность была очевидна для математика X. Харди, и он хотел, чтобы она стала известна биологам, считавшим, что частота доминантной аллели в смешанной популяции может автоматически возрастать.
В строгом смысле закон Харди — Вайнберга приложим к бесконечно большим популяциям, в которых осуществляется панмиксия и на которые не действуют никакие внешние факторы. Только при этих условиях популяция находится в равновесии, т. е. частоты аллелей и генотипов в ней постоянны. Очевидно, что это идеальное представление о популяции никогда не реализуется в природных условиях. На популяцию постоянно действуют многочисленные внешние и внутренние факторы, нарушающие генетическое равновесие, которые будут рассмотрены далее.
Как определить частоту аллелей в популяции при полном доминировании? Поскольку, согласно формуле Харди—Вайнберга, (рА.->- то, зная частоту рецессивных гомозигот, достаточно извлечь квадратный корень из полученной величины, и мы найдем частоту рецессивной аллели q. Частота доминантной аллели составит р = 1 — q. Определив таким образом частоты аллелей А и а, можно выяснить частоты всех генотипов в популяции.
Важным следствием закона Харди—Вайнберга является то, что редкие аллели в популяции присутствуют преимущественно в ге-терозиготе. Например, если аллель а встречается с частотой 0,01, а аллель А — соответственно с частотой 0,99, то частота гетеро-зигот составит 0,0198, т. е. около 0,02, а рецессивных гомозигот — 0,0001. Таким образом, в гетерозиготном состоянии рецессивная аллель встречается более чем в 100 раз чаще, нежели в гомози-готном.
В медицинской генетике закон Харди — Вайнберга позволяет оценить популяционный риск генетически обусловленных заболеваний, поскольку каждая популяция обладает собственным аллелофондом и, соответственно, разными частотами неблагоприятных аллелей. Зная частоты рождения детей с наследственными заболеваниями, можно рассчитать структуру аллелофонда. В то же время, зная частоты неблагоприятных аллелей, можно предсказать риск рождения больного ребёнка.
В селекции — позволяет выявить генетический потенциал исходного материала (природных популяций, а также сортов и пород народной селекции), поскольку разные сорта и породы характеризуются собственными аллелофондами, которые могут быть рассчитаны с помощью закона Харди — Вайнберга. Если в исходном материале выявлена высокая частота требуемого аллеля, то можно ожидать быстрого получения желаемого результата при отборе. Если же частота требуемого аллеля низка, то нужно или искать другой исходный материал, или вводить требуемый аллель из других популяций (сортов и пород).
В работах 1920-х годах С. С. Четвериков обосновывает три основные посылки популяционной генетики:Мутационный процесс в природных условиях протекает точно так же, как и в условиях лаборатории. Поэтому мы вправе распространять по крайней мере некоторые выводы, полученные в лаборатории, на природные ситуации.
Один из таких выводов — непрерывное во времени возникновение новых мутаций у всех видов живых организмов, другой — рецессивность большинства вновь появляющихся мутаций по отношению к аллелям дикого типа, распространенным в природных популяциях.
Характернейшей чертой природных популяций является преобладание в них панмиксии( свободные случайные скрещивания), что делает возможным приложение Закон Харди — Вайнберга.
Генетическая гетерогенность популяций- природные модели популяции представлены в основном гомозиготами по доминантным аллелям. Частота дом аллелей близка к единице. С это точки зрения эволюционный сдвиг в популяции основывается на отборе редких благоприятных аллелей.
Основные факторы генетической эволюции в популяциях – С. С. Четвериков, Р. Фишер, С. Райт, Н. II. Дубинин, Д. Д. Ромашов и др. заложили основы современных идей о факторах, определяющих генетическую эволюцию популяций. Использование формул Харди - Вайнберга позволяет рассчитать генетический состав в популяции в данный момент и определить тенденции его изменений в ряду поколений. В целом популяции видов испытывают постоянную эволюцию их генетической структуры. Основными факторами такой эволюции являются: 1) мутации; 2) отбор (естественный и искусственный); 3) генетико-автоматические процессы, или, по-другому, дрейф генов – процессы чисто случайных изменений концентраций аллелей или зависимых от других генетических процессов – сопряженный дрейф аллелей; 4) миграции – естественные процессы смешения популяций или искусственное скрещивание друг с другом разных пород, сортов и видов. 1. Мутации изменяют частоту генов в популяциях. Частота мутирования гена — 10-5 – 10-7 на поколение. Учитывая большое количество генов у человека (порядка 30 0000), до 6% его гамет несут мутантные гены. Доминантные мутации проявляются уже в первом поколении и сразу же подвергаются действию естественного отбора. Рецессивные мутации (возникают значительно чаще) сначала накапливаются в популяции и только с появлением рецессивных гомозигот начинают проявляться фенотипически и подвергаться действию естественного отбора. Насыщенность природных популяций рецессивными мутациями называется генетическим грузом и имеет большое значение для выживания вида. Генетическим грузом в человеческих популяциях объясняется появление до 5% потомков с генетическими дефектами. Накопление мутантных аллелей способствует комбинативной изменчивости, приводящей к генетической гетерогенности (генетическому полиморфизму) природных популяций. Средняя степень гетерозиготности в популяциях растений составляет 17%, у беспозвоночных — 13,4%, у позвоночных — 6,6%, у человека — около 6,7%. Мутационный процесс обеспечивает разнообразие эволюционного материала. 2. Дрейф генов – это случайные колебания частот генов в малых популяциях. Предположим, что на необитаемый остров попало зерно гетерозиготного самоопыляемого растения. Исходная популяция будет состоять на 100% из гетерозиготных особей (Аа). В первом поколении уже будет содержаться только 50% гетерозиготных особей: Р: Аа х Аа, F1 будет: АА + 2Аа + аа. Гомозиготы (АА и аа) дадут только гомозиготных потомков, а гетерозиготы — расщепление 1:1 (поровну гомо- и гетерозигот), поэтому в F2 уже будет 25% гетерозигот. 3. Изоляция – это ограничение свободы скрещивания. Она способствует дивергенции — разделению популяций на отдельные группы и изменению частот генотипов. Различают географический (горные хребты, реки, проливы и т.п.), генетический (неполноценность гибридов, различные наборы хромосом), экологический (различные экологические ниши, размножение при разных температурах) и морфофизиологический (различия в строении половых органов) типы изоляции. В человеческих популяциях наиболее существенной является эколого-этологическая изоляция, включающая религиозные и морально-этические ограничения браков. В малых человеческих популяциях (демах, изолятах) наблюдаются дрейф генов и инбридинг (родственные браки). Эти браки нежелательны, они приводят к инбредной депрессии, так как у родственников высокая степень вероятности гетерозиготности по одному и тому же рецессивному патологическому гену. Например, частота больных фенилкетонурией при неродственных браках составляет 1:15000, а при родственных — 1:7000, альбинизмом — 1:40000 и 1:3000 соответственно. Мерой генетических последствий инбридинга служит коэффициент инбридинга – это вероятность того, что у какой-либо особи в данном локусе окажутся два аллеля, идентичные по происхождению. У детей одной супружеской пары вероятность одинаковых аллелей в одном локусе равна 1/2. У их детей эта вероятность становится 1/4 (1/2xl/2). При вступлении в брак двоюродных сибсов коэффициент инбридинга равен 1/16 (1/4х l/4). Аутбридииг — неродственные браки. Они поддерживают высокий уровень гетерозиготности. Повышению гeтерозиготности человеческих популяций способствует миграция, масштабы которой огромны особенно в последний десятилетия. Иммиграция поставляет новые аллели или новые комбинации генотипов, а эмиграция изменяет со отношение различных генотипов в популяции. Повышение уровня гетерозиготности является одной из причин акселерации (ускорения развитияи повышения массы тела и роста людей). 4. Естественный отбор элиминирует из популяции менее удачные комбинации генов и избирательно сохраняет более удачные генотипы, тем самым, изменяя частоту генов в популяциях. Интенсивность естественного отбора даже в современных человеческих популяциях довольно высокая: спонтанные аборты составляют примерно 50% всех зачатий; мертворождения — 3%; ранняя детская смертность— 2%; не вступают в брак около 20% людей; примерно 10% браков бесплодны. Таким образом, около 75% людей не вносят свой вклад в генофонд будущих поколений. Помимо естественного отбора в популяциях (в том числе и человеческих) может действовать и контротбор — это отбор неблагоприятных в обычных условиях среды признаков. Например, в странах Западной Африки частота патологического гена серповидно-клеточной анемии довольно высока, в то время как в странах умеренного климата он не встречается. Такая распространенность данного гена объясняется устойчивостью гетерозигот к тропической малярии. Гомозиготы по гемоглобину А (АА) имеют нормальный гемоглобин А, способный связывать и переносить кислород, но болеют и умирают от тропической малярии. Гомозиготы рецессивные (аа) болеют серповидно-клеточной анемией, их гемоглобин 8 не связывает кислород и они погибают в раннем детском возрасте от его недостатка. Гетерозиготы (Аа) содержат и гемоглобин А (переносит кислород) и гемоглобин S (обеспечивает устойчивость к тропической малярии) и выживают в эндемичных по тропической малярии зонах.
Полиморфи́зм— способность некоторых организмов существовать в состояниях с различной внутренней структурой или в разных внешних формах.

Из видеоурока вы узнаете о том, что такое популяция. О том, что популяциям свойственна изменчивость, которая приводит не только к разнообразию в популяции, но и к образованию новых подвидов и видов. Также вы узнаете о том, что в популяциях имеются большие запасы аллелей, которые не приносят им какой-либо пользы в данном месте или в данное время. Они сохраняются в популяции в гетерозиготном состоянии, пока в результате изменения условий среды вдруг не окажутся полезными. В данном уроке приводятся следующие понятия: популяция, естественный отбор, популяционная генетика, генофонд, мутационный процесс, гетерозиготность, изменчивость популяции.
В данный момент вы не можете посмотреть или раздать видеоурок ученикам
Чтобы получить доступ к этому и другим видеоурокам комплекта, вам нужно добавить его в личный кабинет, приобретя в каталоге.
Получите невероятные возможности



Конспект урока "Популяция. Генетический состав популяций"
Вид — это исторически сложившаяся совокупность популяций, особи которых обладают наследственным сходством морфологических, физиологических и биохимических особенностей, могут свободно скрещиваться и давать плодовитое потомство, приспособлены к определённым условиям жизни и занимают определённую область — ареал.
В природе совокупность особей вида, группируются в популяции. Популяции населяют какие-то отдельные небольшие участки ареала.
Популяция — это группа одновидовых организмов, занимающих определённый участок территории внутри ареала вида, свободно скрещивающихся между собой и частично или полностью изолированных от других популяций.

Вильгельм Людвиг Иогансен
Численность популяций, да и количество особей в популяциях из-за изменения каких-либо факторов, например климатических, может меняться. Но сами популяции как таковые все равно сохраняются.
Конечно, природа не стабильна и внутри популяций могут возникать мелкие группировки, в которые объединяются особи со сходным поведением или на основе родственных связей. Такие группировки известны нам как стайки рыб или воробьёв прайды львов. Напомним, что прайды ― это семейные группы львов. В которые входят один или нескольких взрослых самцов (обычно не более 3-х), нескольких половозрелых самок и их детёнышей (львят обоих полов). Численность прайда может достигать до 30−40 животных. В прайдах всегда доминируют львы — альфа-самцы.
Но, как ни странно, охотой занимаются только самки. Молодые самцы по достижении половой зрелости изгоняются из прайда. Живя отдельно, они крепнут, взрослеют, а затем создают свои прайды.

В отличие от популяции, группы не способны устойчиво поддерживать сами себя. Они могут смешиваться с другими или распадаться под влиянием каких-либо факторов.
Популяция же занимает ту часть ареала, в которой отсутствуют изоляционные барьеры.
Каждая популяция приспосабливалась (адаптировалась) к среде своего обитания. Характерные черты этого приспособления передаются из поколения в поколение.

Взаимоотношения организмов в популяциях
Составляющие популяцию организмы связаны друг с другом различными взаимоотношениями.
Часто особи одной популяции конкурируют друг с другом за те или иные виды ресурсов. Например, самка гиены рожает от 1 до 3 детенышей. Если оба детеныша самки, то они начинают сражение за доминирование, используя зубы. Эта конкуренция ведет к гибели почти 25% всех гиен на их первом месяце жизни. В результате оставшийся в живых детёныш получает больше различных ресурсов и созревает быстрее.
Особи одной популяции могут поедать друг друга. Например, речной окунь в отсутствии корма питается собственной молодью.
Также особи одного вида могут группироваться и вместе обороняться от хищника.
Таким образом, можно сказать что внутренние взаимоотношения в популяциях парой очень сложны и противоречивы.
В природе действует естественный отбор. Ослабленные организмы погибают. Это улучшает качественный состав популяции (в том числе качество наследственного материала, которым располагает популяция). Естественный отбор повышает способность популяции к выживанию в меняющихся условиях среды.
Сильные организмы занимают место погибших, размножаются и дают начало новому поколению.

В пределах каждой популяции организмов, размножающихся половым путём, идёт постоянный обмен генетическим материалом. А вот скрещивание особей из разных популяций происходит гораздо реже, поэтому генетический обмен между разными популяциями ограничен.
Существование в форме популяции повышает внутреннее разнообразие вида, его устойчивость к местным изменениям условий жизни, позволяет ему закрепляться в новых для себя условиях.
Генетический состав популяции
Научные генетические открытия и теории Дарвина привели к рождению особого направления исследований — популяционной генетики.
Популяционная генетика позволяет объяснить процессы изменения генетического состава популяций, возникновения новых свойств организмов и их закрепление под воздействием естественного отбора.
Каждая популяция характеризуется своим специфическим набором генов (генофондом) с присущим только данной популяции соотношением частот встречаемости разных аллелей.
Генофонд — это совокупное количество генетического материала, который слагается из генотипов отдельных особей.
Как уже отмечалось, природные популяции в разных частях ареала вида обычно более или менее различны.
Почему популяции одного вида могут отличаться? Дело в том, что чаще всего скрещивание между особями одного вида происходит в пределах одной популяции, которая находится на определённой территории.
В каждой достаточно долго существующей совокупности особей могут спонтанно возникать различные мутации, которые в дальнейшем комбинируются (перемешиваются) с разными, уже имеющимися наследственными свойствами.
Под действием мутаций, которые закрепляются в генофондах популяций, изменяются виды, что ведёт к эволюции организмов.
Поэтому можно сказать, что мутационный процесс — это постоянный источник наследственной изменчивости.
В популяции, состоящей из нескольких миллионов особей, в каждом поколении может возникать по несколько мутаций практически каждого гена, который имеется в этой популяции.
Наследственная изменчивость обусловлена возникновением разных типов мутаций и их комбинаций, которые передаются по наследству и впоследствии проявляются у потомства.
Природные популяции насыщены самыми разнообразными мутациями. На это обратил внимание ещё русский учёный Сергей Сергеевич Четвериков, который установил, что значительная часть изменчивости генофонда скрыта от глаз, так как подавляющее большинство возникающих мутаций рецессивны и не проявляются внешне.
Вспомним, что гетерозиготность ─ это присущее всякому гибридному организму состояние, при котором его гомологичные хромосомы несут разные формы (аллели) того или иного гена или различаются по взаиморасположению генов.

Ген, который способен подавлять действие другого гена в онтогенезе, называется доминантным.
Ген, который оказывается подавленным и не может проявить себя в фенотипе, называется рецессивным.
Обычно доминантный ген обозначается прописной буквой ─ А-большое, рецессивный ген (а-малое). Рецессивный ген проявляется в фенотипе только в тех случаях, когда он находится в паре с аналогичным геном (а-малое, а-малое), т.е. когда оба аллельных гена являются рецессивными.
Если один аллельный ген является доминантным, а второй рецессивным (А-большое, а-малое, то в фенотипе проявляются только признаки, свойственные доминантному гену (А-большое).

Рецессивные мутации можно выявить в экспериментах со скрещиванием близкородственных особей.
При таком скрещивании некоторые рецессивные аллели, находившиеся в гетерозиготном и потому скрытом состоянии, перейдут в гомозиготное состояние и смогут проявиться.
Таким образом, благодаря мутациям популяции могут изменяться.
Изменчивость популяции — это важный фактор эволюции. Она повышает внутреннее разнообразие вида. Позволяет закрепиться виду в новых местах обитания.
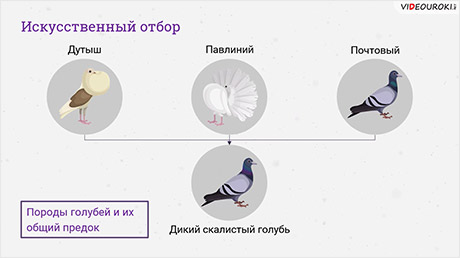
Генетическую изменчивость можно увидеть в ходе искусственного отбора. Когда из популяции выбирают тех особей, у которых какие-либо ценные в хозяйственном отношении признаки выражены наиболее сильно, и скрещивают этих особей между собой.
В фенотипе первого поколения в результате такого скрещивания проявляются необходимые наилучшие выбранные признаки.
Например, у моркови и свёклы ценятся только сладкие корнеплоды, у картофеля — крупные клубни, у мака ― нераскрывающиеся коробочки.
Следовательно, в популяциях возможна генетическая изменчивость буквально по каждому признаку данного организма.
Ещё Сергей Сергеевич Четвериков показал, что большинство вновь возникших мутаций оказываются вредными и, как правило, снижают жизнеспособность особей.
Они сохраняются в популяциях лишь благодаря отбору в пользу гетерозигот.
Однако мутации, вредные в одних условиях, могут повысить жизнеспособность в других условиях.

Так, мутация, вызывающая недоразвитие или полное отсутствие крыльев у насекомых, безусловно, вредна в обычных условиях, и бескрылые особи быстро вытесняются нормальными. Но на островах, где дуют сильные ветры, такие насекомые имеют преимущества перед особями с нормально развитыми крыльями.
Поскольку всякая популяция обычно хорошо приспособлена к своей среде обитания, крупные изменения обычно снижают эту приспособленность.
В популяциях имеются большие запасы таких аллелей, которые не приносят ей какой-либо пользы в данном месте или в данное время; они сохраняются в популяции в гетерозиготном состоянии, пока в результате изменения условий среды вдруг не окажутся полезными.
Как только условия среды изменяются, частота новых аллелей под действием отбора начинает возрастать, и в конечном счёте они становятся основным генетическим материалом.
Таким образом, благодаря приспособлениям к новым условиям среды, популяции адаптируются.
Например, к изменению климата, появлению нового хищника или конкурента и даже к загрязнению среды человеком.
Примером подобной адаптации служит эволюция видов насекомых, устойчивых к инсектицидам. При введении в практику нового инсектицида (яда, действующего на насекомых) для успешной борьбы с насекомым-вредителем бывает достаточно небольшого его количества.
С течением времени концентрацию инсектицида приходится повышать, пока, наконец, он не оказывается недейственным.
Гены, способные обеспечить устойчивость к инсектицидам, очевидно, имелись в каждой из популяций этих видов; их действие и обеспечило в конечном итоге снижение эффективности ядов, использованных для борьбы с вредителями.
Таким образом, мутационный процесс создаёт материал для эволюционных преобразований, формируя резерв наследственной изменчивости в генофонде каждой популяции и виде в целом. Поддерживая высокую степень генетического разнообразия популяций, он создаёт основу для действия естественного отбора и микроэволюции.
Генетика популяций — важнейшая отрасль генетики, которая занимается изучением генетической структуры природных популяций, а также различных происходящих в ней генетических процессов.
Большое значение генетика популяций имеет в развитии эволюционной теории. Достижения генетики популяций позволили установить, что популяция является не просто единицей вида, но также и полноценной единицей эволюции.
Первым в этом направлении стал работать русский генетик С. С. Четвериков — это были 20-е годы 20 века. Развитие этой науки также связано с такими именами как:
- С. М. Гершинзон;
- Д. Д. Ромашов;
- Р. Фишер;
- Н. П. Дубинин;
- Ф. Г. Добржанский;
- Р. Райт.
В современном понимании популяция представляет собой специфическую и принадлежащую одному виду совокупность особей, которая характеризуется наличием общего генофонда и общей территории.
Закон Харди-Вайнберга
Популяция включает в себе особей одного вида. Каждая из этих особей отличается одинаковым набором генов. Но каждый ген может быть представлен разными аллелями — их число может быть довольно большим. Поэтому стоит сделать замечание, что популяция представляет собой все же совокупность неодинаковых в генетическом плане особей, отличающихся различными состояниями свойственными им признаками.
Популяции одного и того же вида в природе нередко отличаются частотами встречаемости конкретных аллельных генов.
Важная задача популяционной генетики — изучение закономерностей распределения аллелей в популяциях. Именно этим вопросом занимались такие ученые как В. Вайнберг и Г. Харди. Согласно их исследованиям, в достаточно больших популяциях не возникают новые мутации, даже при условии, что особи свободно скрещиваются между собой, а какой-либо внешний фактор, влияющий на определенное сочетание аллелей, отсутствует. Также не происходит обмен генетической информацией с другими популяциями в ходе миграции особей.
Происходит стабилизация соотношения аллелей на протяжении нескольких поколений. И длительное время такое соотношение остается неизменным.
Из установленной Г. Харди и В. Вайнбергом закономерности следует, что при постоянстве внешних и внутренних факторов частота встречаемости аллелей у больших популяций, находящихся в изоляции от других, длительное время остается устойчивой.
Для описания распределения частот в панмиксической популяции по одной паре аллельных генов (А-а) ученые придумали формулу (формула Харди-Вайнберга). Эта формула служит простейшей моделью и основой для проведения более сложных популяционно-генетических исследований.
Читайте также: