
Электрический синапс это кратко
Обновлено: 05.07.2024
Используемый довод статьи (аксиома):
Электрический синапс судя по всему эволюционно один из первых вариантов передачи сигналов между клетками, позволяющий развить активность нейронов максимально быстро, но из за сопротивления мембран лишь на ограниченное расстояние. Такие синапсы обычно двунаправленного действия, что идеально подходит для одновременного возбуждения нейронов, выполняющих одинаковую функцию. Однако и тут эволюция пыталась провести модернизацию, создавая потенциалзависимые ионные каналы. С появлением более продвинутых в плане возможностей химических синапсов доля электрических в нервной системе неуклонно падала по мере ее усложнения.
Вес уверенности:
Электрический синапс - место высокоспециализированных щелевых контактов между нейрон ами , где происходит прямое перетекание электрических токов от одного нейрон а к другому . В щелевых контактах мембраны соседних клеток находятся на расстоянии около 3,8 нм , в то время как в химическом синапс е расстояние между двумя нейрон ами составляет от 20 до 40 нм . У многих животных в нервной системе имеются как химические, так и электрические синапс ы . По сравнению с химическими синапс ами, электрические синапс ы проводят нервные импульсы быстрее, однако, в отличие от химических синапс ов, сигнал на постсинаптическом нейрон е оказывается равным или меньше первоначального сигнала. Электрические синапс ы активно используют животные, которым необходима способность развивать наиболее быстрый ответ, какой возможно, например, в случае защитных рефлексов . Как правило, электрические синапс ы двунаправленны, то есть нервный импульс может проходить по ним в обоих направлениях .

Строение щелевого контакта: а — коннексон в закрытом состоянии; b — коннексон в открытом состоянии; с — коннексон, встроенный в мембрану; d — мономер коннексина, е — плазматическая мембрана; f — межклеточное пространство; g — промежуток в 2—4 нанометра в электрическом синапс е; h — гидрофильный канал коннексона.
Функции
Простота устройства электрических синапс ов позволяет им проводить сигнал очень быстро, однако они участвуют лишь в простых поведенческих реакциях, в отличие от более сложно устроенных химических синапс ов. Поскольку для передачи сигнала через электрический синапс не нужно связывание рецептор а с сигнальной молекулой-лигандом, при работе электрических синапс ов не происходит задержки, которая у химических синапс ов может составлять от 0,5 до 4 миллисекунд. Однако у млекопитающих различия в скоростях проведения сигнала электрическим и химическим синапс ом не различаются так сильно, как у холоднокровных животных. Благодаря высокой скорости проведения сигнала электрическим синапс ам несколько соседних нейрон ов развивают потенциал действия практически одновременно. Ответ постсинаптического нейрон а имеет тот же знак, что и изменения в пресинаптическом нейрон е. Так, деполяризация пресинаптической мембраны всегда вызовет деполяризацию постсинаптической мембраны, то же самое имеет место для гиперполяризации . Как правило, ответ постсинаптического нейрон а меньше, чем амплитуда исходного сигнала; это обусловлено сопротивлением пре- и постсинаптической мембран.
Распространение
Электрические синапс ы встречаются по всей центральной нервной системе. Они были детально изучены в неокортексе, гиппокампе, таламическом ретикулярном ядре , голубом пятне, нижнем оливковом ядре , мезенцефалическом ядре тройничного нерва , обонятельных луковицах, сетчатке и спинном мозге позвоночных. Электрические синапс ы были обнаружены также в полосатых телах, мозжечке и супрахиазматическом ядре. Как правило, электрические синапс ы опосредуют очень быстрые поведенческие ответы, как, например, выпускание чернил морским зайцем Aplysia при опасности. Электрические синапс ы были обнаружены и вне нервной системы. Электрические синапс ы связывают рецептор ные клетки , кардиомиоциты, гладкомышечные клетки, клетки печени, глиальные и эпителиальные клетки.
Электрические синапс ы наиболее характерны для низкоорганизованных животных. В ходе эволюции доля электрических синапс ов уменьшалась, и в ЦНС млекопитающих (в том числе человека) на долю электрических синапс ов приходится около 1 % связей между нейрон ами.
Источник: Электрический синапс
Дата создания: 29.10.2020
Последнее редактирование: 21.12.2020
Чтобы оставить комментарии нужно авторизоваться: Авторизация пользователя
Область контакта между двумя нейронами называют синапсом.
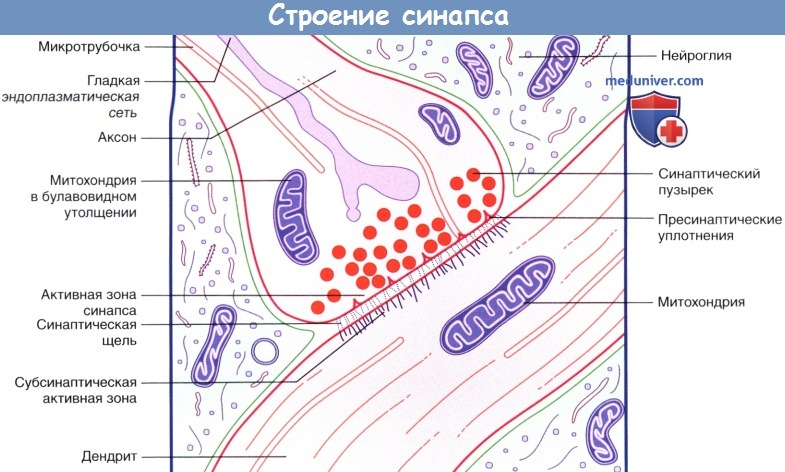
Внутреннее строение аксодендритического синапса.
а) Электрические синапсы. Электрические синапсы в нервной системе млекопитающих встречаются редко. Они образованы щелевидными контактами (нексусами) между дендритами или сомами соприкасающихся нейронов, которые соединяются с помощью цитоплазматических каналов диаметром 1,5 нм. Процесс передачи сигнала происходит без синаптической задержки и без участия медиаторов.
Посредством электрических синапсов возможно распространение электротонических потенциалов от одного нейрона к другому. Вследствие тесного синаптического контакта модуляция проведения сигнала невозможна. Задача этих синапсов — осуществление одновременного возбуждения нейронов, выполняющих одинаковую функцию. Примером служат нейроны дыхательного центра продолговатого мозга, которые во время вдоха синхронно генерируют импульсы. Кроме того, примером могут служить нейронные цепи, управляющие саккадами, при которых точка фиксации взора перемещается от одного объекта внимания к другому.
б) Химические синапсы. Большинство синапсов нервной системы — химические. Функционирование таких синапсов зависит от высвобождения медиаторов. Классический химический синапс представлен пресинаптической мембраной, синаптической щелью и постсинаптической мембраной. Пресинаптическая мембрана — часть булавовидного расширения нервного окончания клетки, передающей сигнал, а постсинаптическая мембрана — часть клетки, получающей сигнал.
Медиатор высвобождается из булавовидного расширения посредством экзоцитоза, проходит через синаптическую щель и связывается с рецепторами на постсинаптической мембране. Под постсинаптической мембраной расположена субсинаптическая активная зона, в которой после активации рецепторов постсинаптической мембраны происходят разнообразные биохимические процессы.
В булавовидном расширении расположены содержащие медиаторы синаптические пузырьки, а также большое количество митохондрий и цистерны гладкой эндоплазматической сети. Применение традиционных методик фиксации при исследовании клеток позволяет различить на пресинаптической мембране пресинаптические уплотнения, ограничивающие активные зоны синапса, к которым при помощи микротрубочек направляются синаптические пузырьки.
Аксодендритический синапс.
Срез препарата спинного мозга: синапс между концевым участком дендрита и, предположительно, двигательным нейроном.
Наличие округлых синаптических пузырьков и постсинаптического уплотнения характерно для возбуждающих синапсов.
Срез дендрита проведен в поперечном направлении, о чем свидетельствует наличие множества микротрубочек.
Кроме того, видны некоторые нейрофиламенты. Участок синапса окружен протоплазматическим астроцитом. Процессы, происходящие в нервных окончаниях двух типов.
(А) Синаптическая передача небольших молекул (например, глутамата).
(1) Транспортные пузырьки, содержащие мембранные белки синаптических пузырьков, направляются вдоль микротрубочек к плазматической мембране булавовидного утолщения.
В это же время происходит перенос молекул ферментов и глутамата путем медленного транспорта.
(2) Мембранные белки пузырьков выходят из плазматической мембраны и формируют синаптические пузырьки.
(3) Глутамат погружается в синаптические пузырьки; происходит накопление медиатора.
(4) Пузырьки, содержащие глутамат, подходят к пресинаптической мембране.
(5) В результате деполяризации происходит экзоцитоз медиатора из частично разрушенных пузырьков.
(6) Высвобождающийся медиатор распространяется диффузно в области синаптической щели и активирует специфические рецепторы на постсинаптической мембране.
(7) Мембраны синаптических пузырьков транспортируются обратно в клетку путем эндоцитоза.
(8) Происходит частичный обратный захват глутамата в клетку для повторного использования.
(Б) Передача нейропептидов (например, субстанции Р), осуществляющаяся одновременно с синаптической передачей (например, глутамата).
Совместная передача этих веществ происходит в центральных нервных окончаниях униполярных нейронов, обеспечивающих болевую чувствительность.
(1) Синтезированные в комплексе Гольджи (в области перикариона) пузырьки и предшественники пептидов (пропептиды) транспортируются к булавовидному расширению путем быстрого транспорта.
(2) При их попадании в область булавовидного утолщения завершается процесс формирования молекулы пептида, и пузырьки транспортируются к плазматической мембране.
(3) Деполяризация мембраны и перенос содержимого пузырьков в межклеточное пространство путем экзоцитоза.
(4) Одновременно с этим происходит высвобождение глутамата.
1. Активация рецепторов. Молекулы медиаторов проходят через синаптическую щель и активируют рецепторные белки, расположенные парами на постсинаптической мембране. Активация рецепторов запускает ионные процессы, которые приводят к деполяризации постсинаптической мембраны (возбуждающее постсинаптическое действие) или гиперполяризации постсинаптической мембраны (тормозящее постсинаптическое действие). Изменение электротонуса передается в сому в виде затухающего по мере распространения электротонического потенциала, за счет которого происходит изменение потенциала покоя в начальном сегменте аксона.
Ионные процессы подробно описаны в отдельной статье на сайте. При преобладании возбуждающих постсинаптических потенциалов начальный сегмент аксона деполяризуется до порогового уровня и генерирует потенциал действия.
Наиболее распространенный возбуждающий медиатор ЦНС — глутамат, а тормозной — гамма-аминомасляная кислота (ГАМК). В периферической нервной системе медиатором для двигательных нейронов поперечно-полосатой мускулатуры служит ацетилхолин, а для чувствительных нейронов — глутамат.
Последовательность процессов, происходящих в глутаматергических синапсах, показана на рисунке ниже. При передаче глутамата совместно с другими пептидами высвобождение пептидов осуществляется внесинаптическим путем.
Большинство чувствительных нейронов помимо глутамата выделяет и другие пептиды (один или несколько), высвобождающиеся в различных участках нейрона; однако основная функция этих пептидов — модуляция (повышение или снижение) эффективности синаптической передачи глутамата.
Кроме того, нейротрансмиссия может происходить путем диффузной внесинаптической передачи сигнала, характерной для моноаминергических нейронов (нейронов, использующих биогенные амины для обеспечения нейротрансмиссии). Выделяют две разновидности моноаминергических нейронов. В одних нейронах осуществляется синтез катехоламинов (норадреналина или дофамина) из аминокислоты тирозина, а в других — серотонина из аминокислоты триптофана. Например, дофамин высвобождается как в синаптической области, так и из варикозных утолщений аксона, в которых также происходит синтез этого нейромедиатора.
Дофамин проникает в межклеточную жидкость ЦНС и до момента деградации способен активировать специфические рецепторы на расстоянии до 100 мкм. Моноаминергические нейроны присутствуют во многих структурах ЦНС; нарушение передачи импульса этими нейронами приводит к различным заболеваниям, среди которых выделяют болезнь Паркинсона, шизофрению и глубокую депрессию.
Оксид азота (газообразная молекула) также участвует в диффузной нейропередаче в глутаматергической системе нейронов. Избыточное влияние оксида азота оказывает цитотоксическое действие, особенно в тех участках, кровоснабжение которых нарушено за счет тромбоза артерий. Глутамат также является потенциально цитотоксическим нейромедиатором.
Существует три вида химических межнейрональных взаимодействий: синаптическое (например, глутаматергическое), внесинаптическое (пептидергическое) и диффузное (например, моноаминергическое, серотонинергическое).
Химические синапсы классифицируют по анатомическому строению на аксодендритические, аксосоматические, аксоаксональные и дендро-дендритические. Синапс представлен пре- и постсинаптическими мембранами, синаптической щелью и субсинаптической активной зоной.
Электрические синапсы обеспечивают одновременную активацию целых групп нейронов, образуя между ними электрические связи за счет щелевидных контактов (нексусов).
Диффузная нейротрансмиссия в головном мозге.
Аксоны глутаматергического (1) и дофаминергического (2) нейронов образуют плотные синаптические контакты с отростком звездчатого нейрона (3) полосатого тела.
Дофамин высвобождается не только из пресинаптической области, но и из варикозного утолщения аксона, откуда диффузно распространяется в межклеточное пространство и активирует дофаминовые рецепторы дендритного ствола и стенки перицита капилляра. Растормаживание.
(А) Возбуждающий нейрон 1 активирует тормозной нейрон 2, который в свою очередь затормаживает нейрон 3.
(Б) Появление второго тормозного нейрона (2б) оказывает противоположное влияние на нейрон 3, поскольку происходит торможение нейрона 2б.
Спонтанно-активный нейрон 3 генерирует сигналы в условиях отсутствия тормозных влияний.
3. Торможение и растормаживание. Функционирование спонтанно-активных нейронов сдерживается под влиянием тормозных нейронов (обычно, ГАМКергических). Деятельность тормозных нейронов, в свою очередь, может быть ингибирована воздействующими на них другими тормозными нейронами, в результате чего происходит растормаживание клетки-мишени. Процесс растормаживания — важная особенность нейрональной активности в базальных ганглиях.
4. Редкие виды химических синапсов. Выделяют два типа аксоаксональных синапсов. В обоих случаях булавовидное утолщение образует тормозной нейрон. Синапсы первого типа образуются в области начального сегмента аксона и передают мощное ингибирующее влияние тормозного нейрона. Синапсы второго типа образуются между булавовидным утолщением тормозного нейрона и булавовидными утолщениями возбуждающих нейронов, что приводит к угнетению высвобождения медиаторов. Этот процесс получил название пресинаптического торможения. В этом плане традиционный синапс обеспечивает постсинаптичсекое торможение.
Дендро-дендритические (Д-Д) синапсы образуются между дендритными шипиками дендритов смежных шипиковых нейронов. Их задача — не генерирование нервного импульса, а изменение электротонуса клетки-мишени. В последовательных Д-Д-синапсах синаптические пузырьки располагаются только в одном дендритном шипике, а в реципрокном Д-Д-синапсе— в обоих. Возбуждающие Д-Д-синапсы изображены на рисунке ниже. Тормозные Д-Д-синапсы широко представлены в переключающих ядрах таламуса.
Кроме того, выделяют немногочисленные сомато-дендритические и сомато-соматические синапсы.
Аксоаксональные синапсы коры головного мозга.
Стрелками указано направление проведения импульсов. (1) Пресинаптическое и (2) постсинаптическое торможение спинномозгового нейрона, направляющегося к головному мозгу.
Стрелками указано направление проведения импульсов (возможно торможение переключательного нейрона под действием тормозных влияний). Возбуждающие дендро-дендритические синапсы. Изображены дендриты трех нейронов.
Реципрокный синапс (справа). Стрелками указано направление распространения электрото-нических волн.
Электрический синапс — механическая и электропроводящая связь между двумя примыкающими нейронами, которая образуется в узкой щели между пре- и постсинаптическими нейронами (т. н. щелевой контакт).
Различают пре- и постсинаптическую часть. В эфапсе I типа они разделены щелью в 2 нм (обычное межклеточное — 20 нм), во II типе щель отсутствует [1] .
В отличие от химического синапса, в электрическом синапсе клетки соединяются высокопроницаемыми контактами с помощью особых коннексонов (каждый коннексон состоит из шести белковых субъединиц — коннексинов).
В эфапсе I типа при деполяризации ток пропускается в одном направлении, при гиперполяризации — в обратном.
В эфапсе II типа потенциал действия следует в обоих направлениях.
Содержание
Распространение

Строение электрического синапса: а — коннексон в закрытом состоянии; b — коннексон в открытом состоянии; с — коннексон, встроенный в мембрану; d — мономер коннексина, е — плазматическая мембрана; f — межклеточное пространство; g — промежуток в 2-4 нанометра в электрическом синапсе; h — гидрофильный канал коннексона.
В первой половине ХХ века считалось, что передача нервного сигнала с помощью химического синапса присуща периферийним нервно-мускульным контактам позвоночных (автономная мускулатура и моторные контакты), но, в основном из-за нехватки опытных данных, считалось, что синапсы в Центральной нервной системе (ЦНС) используют непосредственно электрическую передачу. Но, после отработки методик внутриклеточной записи электрических потенциалов в 1950-х годах, было показано, что соединение нейронов химическими синапсами является правилом для ЦНС позвоночных.
В дальнейшем некоторые синапсы в нервной системе ракообразных были идентифицированы как электрические. В настоящее время обнаружено небольшое количество электрических синапсов в мозге млекопитающих, и большая их доля - в ЦНС низших позвоночных, особенно рыб. У беспозвоночных электрические синапсы довольно обычны в нервной системе аннелид и ракообразных. Электрические синапсы обычно расположены на таких нервных путях, где требуется соблюдение точно определенного времени прохождения нервного импульса — учитывая, что электрический синапс не вызывает заметной на опыте задержки сигнала.
Строение
Указанные свойства отражаются и на микроструктуре электрического синапса, обнаруженной с помощью электронного микроскопа.
В отличие от химического синапса, синаптическая щель в электрическом синапсе чрезвычайно узка (около 3,5 нанометров [2] ). Через синаптическую щель данного типа синапсов проходят пространственно упорядоченные гидрофильные протеиновые туннели, каждый примерно 5 нанометров шириной, которые перфорируют пре- и постсинаптическую мембрану и называются коннексонами. У первичноротых организмов (нематоды, моллюски, членистоногие) коннексоны сформированы протеинами специфической структуры, называемые паннексинами (англ.) или иннексинами (англ.); у вторичноротых (иглокожие, асцидии, позвоночные) коннексоны построены из протеинов другого типа — коннексинов, которые кодируются другой группой генов.
Млекопитающие являются в этом плане менее дифференцированными вторичноротыми, и в их организме наряду с коннексинамы производятся также и паннексины, но до сих пор у позвоночных не выявлено ни одного электрического синапса, где межклеточные каналы были бы сформированы паннексинами.
Электрические синапсы позволяют осуществлять электрическую проводимость в обоих направлениях (в отличие от химических); тем не менее, в последнее время у некоторых ракообразных были открыты направляющие электрические синапсы, то есть такие, которые позволяют осуществлять прохождение нервного сигнала только в одном направлении.
Функции
Описанные свойства электрических синапсов определяют их функции в нервной системе различных организмов.
Благодаря прямому переходу ионов через такой синапс нервный сигнал передается ими практически без задержки. Это позволяет ракообразным, у которых такие синапсы совмещают нейроны, отвечающие за двигательную активность, минимизировать время между появлением опасности и моторной реакцией на нее — что часто является критическим при бегстве от хищника.
Тот факт, что размер поры коннексонов электрических синапсов делает возможным переход из клетки в клетку молекул АТФ и вторичных мессенджеров, также играет важную роль в обеспечении синхронизации возбуждения и метаболизма нейронов — последнее особенно важно для клеток нейроглии, сочетание которых электрическими синапсами было открыто недавно.
Примечания
Литература
Сома · Аксон (Аксонный холмик, Терминаль аксона, Аксоплазма, Аксолемма, Нейрофиламенты)
Строение щелевого контакта: а — коннексон в закрытом состоянии; b — коннексон в открытом состоянии; с — коннексон, встроенный в мембрану; d — мономер коннексина, е — плазматическая мембрана; f — межклеточное пространство; g — промежуток в 2—4 нанометра в электрическом синапсе; h — гидрофильный канал коннексона.
Электри́ческий си́напс (англ. electrical synapse ) — место высокоспециализированных контактов (щелевых контактов) между нейронами, где происходит прямое перетекание электрических токов от одного нейрона к другому [1] . В щелевых контактах мембраны соседних клеток находятся на расстоянии около 3,8 нм, в то время как в химическом синапсе расстояние между двумя нейронами составляет от 20 до 40 нм [2] . У многих животных в нервной системе имеются как химические, так и электрические синапсы. По сравнению с химическими синапсами, электрические синапсы проводят нервные импульсы быстрее, однако, в отличие от химических синапсов, сигнал на постсинаптическом нейроне оказывается равным или меньше первоначального сигнала. Электрические синапсы активно используют животные, которым необходима способность развивать наиболее быстрый ответ, какой возможно, например, в случае защитных рефлексов. Как правило, электрические синапсы двунаправленны, то есть нервный импульс может проходить по ним в обоих направлениях [3] .
Читайте также: