
Биосинтез нуклеиновых кислот кратко
Обновлено: 05.07.2024
Раздел ЕГЭ: 2.6. Генетическая информация в клетке. Гены, генетический код и его свойства. Матричный характер реакций биосинтеза. Биосинтез белка и нуклеиновых кислот
К реакциям матричного синтеза относят репликацию ДНК, синтез и-РНК на ДНК (транскрипцию) и синтез белка на и-РНК (трансляцию), а также синтез РНК или ДНК на РНК вирусов.
Биосинтез белка — это один из видов пластического обмена, в ходе которого наследственная информация, закодированная в генах ДНК, реализуется в определенную последовательность аминокислот в белковых молекулах. В биосинтезе белка выделяют два основных этапа: транскрипцию и трансляцию.
Репликация ДНК
Структура молекулы ДНК, установленная Дж. Уотсоном и Ф. Криком в 1953 г., отвечала тем требованиям, которые предъявлялись к молекуле-хранительнице и передатчику наследственной информации. Молекула ДНК состоит из двух комплементарных цепей. Эти цепи удерживаются слабыми водородными связями, способными разрываться под действием ферментов. Процесс удвоения ДНК происходит полуконсервативным способом: молекула ДНК расплетается, и на каждой из цепей синтезируется новая цепь по принципу комплементарности. Процесс самовоспроизведения молекулы ДНК, обеспечивающий точное копирование наследственной информации и передачу ее из поколения в поколение, называется репликацией.
Передача информации и синтез белка идут по матричному принципу, сравнимому с работой печатного станка в типографии. Информация от ДНК многократно копируется. Если при копировании произойдут ошибки, то они повторятся во всех последующих копиях. Правда, некоторые ошибки при копировании информации молекулой ДНК могут исправляться. Этот процесс устранения ошибок называется репарацией. Первой из реакций в процессе передачи информации является репликация молекулы ДНК и синтез новых цепей ДНК.
Репликация — это процесс самоудвоения молекулы ДНК, осуществляемый под контролем ферментов. На каждой из цепей ДНК, образовавшихся после разрыва водородных связей, при участии фермента ДНК-полимеразы синтезируется дочерняя цепь ДНК. Материалом для синтеза служат свободные нуклеотиды, имеющиеся в цитоплазме клеток.
Биологический смысл репликации заключается в точной передаче наследственной информации от материнской молекулы к дочерним, что в норме и происходит при делении соматических клеток.
Биосинтез белка и нуклеиновых кислот
В биосинтезе белка выделяют два основных этапа: транскрипцию и трансляцию.
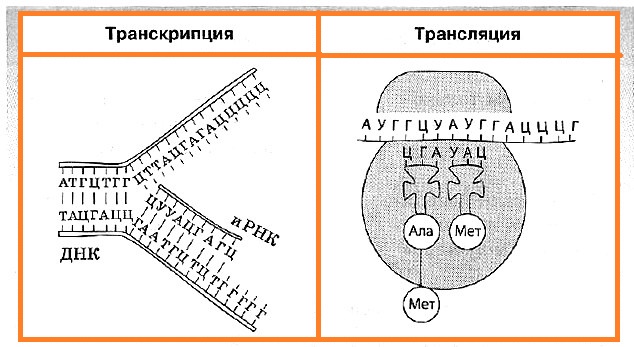
Транскрипция — это биосинтез молекул иРНК на соответствующих участках ДНК. Транскрипция происходит только на одной цепи ДНК, которая называется кодирующей, в отличие от другой — некодирующей, или кодогенной. Обеспечивает процесс переписывания специальный фермент РНК-полимераза, который подбирает нуклеотиды РНК по принципу комплементарности.
Синтезированные в процессе транскрипции в ядре молекулы иРНК покидают его через ядерные поры, а митохондриальные и пластидные иРНК остаются внутри органоидов. После транскрипции происходит процесс активации аминокислот, в коде которой аминокислота присоединяется к соответствующей свободной тРНК.
Трансляция — это биосинтез полипептидной цепи на молекуле иРНК, при котором происходит перевод генетической информации в последовательность аминокислот полипептидной цепи.
Второй этап синтеза белка чаще всего происходит в цитоплазме, например на шероховатой ЭПС. Для его протекания необходимы наличие рибосом, активация тРНК, в ходе которой они присоединяют соответствующие аминокислоты, присутствие ионов Mg 2+ , а также оптимальные условия среды (температура, рН, давление и т. д.).
Для начала транскрипции (инициации) к молекуле иРНК присоединяется малая субъединица рибосомы, а затем по принципу комплементарности к первому кодону АУГ подбирается тРНК, несущая аминокислоту метионин. Лишь после этого присоединяется большая субъединица рибосомы. В пределах собранной рибосомы оказываются два кодона иРНК, первый из которых уже занят. К соседнему с ним кодону присоединяется вторая тРНК, также несущая аминокислоту, после чего между остатками аминокислот с помощью ферментов образуется пептидная связь.
Когда рибосома передвигается на один кодон иРНК, первая из тРНК, освободившаяся от аминокислоты, возвращается в цитоплазму за следующей аминокислотой, а фрагмент будущей полипептидной цепи как бы повисает на оставшейся тРНК. К новому кодону, оказавшемуся в пределах рибосомы, присоединяется следующая тРНК, процесс повторяется, и шаг за шагом полипептидная цепь удлиняется, то есть происходит ее элонгация.
Окончание синтеза белка (терминация) происходит, как только в молекуле иРНК встретится специфическая последовательность нуклеотидов, которая не кодирует аминокислоту (стоп-кодон). После этого рибосома, иРНК и полипептидная цепь разделяются, а вновь синтезированный белок приобретает соответствующую структуру и транспортируется в ту часть клетки, где он будет выполнять свои функции.
Трансляция является весьма энергоемким процессом, поскольку на присоединение одной аминокислоты к тРНК расходуется энергия одной молекулы АТФ, еще несколько используются для продвижения рибосомы по молекуле иРНК.
Репликация ДНК и синтез белка в клетке протекают по принципу матричного синтеза, поскольку новые молекулы нуклеиновых кислот и белков синтезируются в соответствии с программой, заложенной в структуре ранее существовавших молекул тех же нуклеиновых кислот (ДНК или РНК).
Субстратом для синтеза нуклеиновых кислот в клетках и тканях могут служить продукты гидролиза ДНК и РНК, а также продукты обмена белков и углеводов.
Биосинтез пуриновых нуклеотидов происходит из углекислого газа, глицина, муравьиной кислоты, аспарагиновой кислоты и глутамина. В результате поэтапного синтеза из этих компонентов (всего 11 реакций) образуется инозиновая кислота, из которой затем АМФ и ГМФ. Из указанных монофосфатов затем синтезируются соответствующие ди- и трифосфаты.
Синтез пиримидиновых нуклеотидов происходит также поэтапно, вначале образуется карбомоилфосфат из NH3, CO2, АТФ. Затем с участием аспарагиновой кислоты образуется дигидрооротовая кислота, затем оротовая кислота.
Оротовая кислота далее взаимодействует с 5'-фосфорибозил-1-пирофосфатом, при этом синтезируется оротидин-5 -фосфат, который затем декарбоксилируется с образованием уридинмонофосфата. Из уридиловой кислоты при аминировании углеродов за счет глутамина образуется цитидинмонофосфат, а при метилировании – тимидиловая кислота:
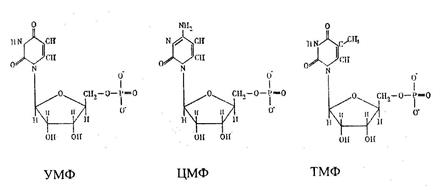
Эти монофосфаты затем фосфорилируются с участием АТФдо образования трифосфатов.
Синтез ДНК (или репликация ДНК). В клетках животных синтез (репликация) ДНК происходит только в определенный период жизни клетки – в синтетической S-фазе. Эта фаза отделена от митоза (M) предсинтетическим (G1 ) и постсинтетическим (G2) периодами (рис. 12.1).
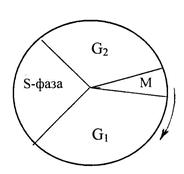
Рис. 12.1. Цикл деления клеток млекопитающих.
Фаза синтеза ДНК (S-фаза) отделена от митоза (M) периодами G1 и G2. Стрелкой указано направление синтеза.
Все этапы синтеза ДНК эукариот осуществляются с помощью репликативного мультиферментного комплекса (праймосомы), состоящего из ферментов и белковых факторов, осуществляющих узнавание точки ori, расплетание родительского дуплекса, удержание материнских цепей ДНК на достаточном расстоянии друг от друга, инициацию новых дочерних цепей, закручивание цепей в спираль и, наконец, терминацию репликации ДНК. В составе комплекса присутствуют новосинтезированная ДНК, не менее 30 белков с Мм от 15 до 300 кДа, ДНК-полимеразы 5, а, праймаза, 3'-5'экзонуклеаза, ДНК-лигаза I, РНКаза H, ДНК-топоизомераза I, ДНК-хеликаза, белок PCNA (ядерный антиген пролиферирующих клеток) и другие факторы.
В праймасоме репликативный белок A (RPA) связывается с однонитевой ДНК и существует в виде комплекса из трех белков с Мм 70, 34 и 11 кДа. Репликативный фактор С (RFC) состоит из пяти белковых елиниц и играет важную роль при связывании репликативных полимераз во время образования репликативной вилки. Белок PCNA, так называемый ДНК-полимеразный зажим, функционирует как кофактор ДНК-полимеразы, образуя закрытое кольцо, окружающее ДНК с каналом в центре. Репликативный фактор С совместно с PCNA продвигается вдоль ДНК по репликативной вилке. С участием праймасомы, ферментов образуются новые полинуклеотидные цепи. Синтез новых цепей всегда идет в направлении от 5'-конца к 3'-концу. Поэтому на одной из ветвей репликативной вилки новая цепь наращивается непрерывно по мере раскручивания ДНК-матрицы, а на другой ветви образуются короткие фрагменты новой цепи ДНК, называемые фрагменты Оказаки; затем концы этих фрагментов соединяются между собой в результате действия фермента ДНК-лигазы (рис. 12.2.).
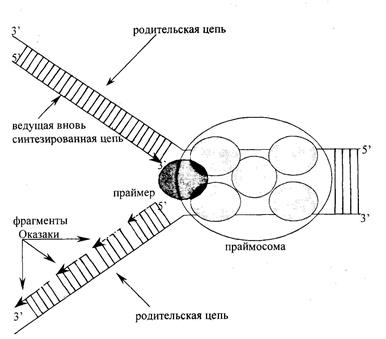
Рис. 12.2. Схематическое изображение синтеза ДНК в репликативной вилке.

На одной ветви репликативной вилки нуклеотидная цепь синтезируется непрерывно, на другой – прерывисто, с образованием фрагментов Оказаки. Фрагменты Оказаки соединяются между собой в результате действия ДНК-лигазы. По мере роста новых цепей репликативная вилка перемещается по ДНК и в результате происходит репликация (удвоение) молекулы ДНК. Последовательности нуклеотидов новой цепи ДНК строго комплементарны последовательностям родительской цепи, благодаря этому соблюдается передача наследственной информации из поколения в поколение (см. рис.3.5.). Точность репликации очень высока – возможна одна ошибка на 10 трансферазных реакций.
Синтез ДНК на матрице РНК происходит под действием фермента ревертазы или обратной транскриптазы (РНК-зависимой ДНК-полимеразы), содержащейся в составе онковирусов. Доказано, что многие РНК-содержащие онкогенные вирусы (онковирусы) содержат этот фермент в составе покровных белков. При этом механизм синтеза ДНК на матрице РНК включает 3 стадии: на первом этапе фермент ревертаза синтезирует на матрице вирусной РНК комплементарную цепь ДНК, что приводит к формированию гибридной молекулы. Затем исходная молекула РНК отделяется от ДНК и только после этого на матрице ДНК синтезируется комплементарная цепь ДНК.
Субстратом для синтеза нуклеиновых кислот в клетках и тканях могут служить продукты гидролиза ДНК и РНК, а также продукты обмена белков и углеводов.
Биосинтез пуриновых нуклеотидов происходит из углекислого газа, глицина, муравьиной кислоты, аспарагиновой кислоты и глутамина. В результате поэтапного синтеза из этих компонентов (всего 11 реакций) образуется инозиновая кислота, из которой затем АМФ и ГМФ. Из указанных монофосфатов затем синтезируются соответствующие ди- и трифосфаты.
Синтез пиримидиновых нуклеотидов происходит также поэтапно, вначале образуется карбомоилфосфат из NH3, CO2, АТФ. Затем с участием аспарагиновой кислоты образуется дигидрооротовая кислота, затем оротовая кислота.
Оротовая кислота далее взаимодействует с 5'-фосфорибозил-1-пирофосфатом, при этом синтезируется оротидин-5 -фосфат, который затем декарбоксилируется с образованием уридинмонофосфата. Из уридиловой кислоты при аминировании углеродов за счет глутамина образуется цитидинмонофосфат, а при метилировании – тимидиловая кислота:
Эти монофосфаты затем фосфорилируются с участием АТФдо образования трифосфатов.
Синтез ДНК (или репликация ДНК). В клетках животных синтез (репликация) ДНК происходит только в определенный период жизни клетки – в синтетической S-фазе. Эта фаза отделена от митоза (M) предсинтетическим (G1 ) и постсинтетическим (G2) периодами (рис. 12.1).
Рис. 12.1. Цикл деления клеток млекопитающих.
Фаза синтеза ДНК (S-фаза) отделена от митоза (M) периодами G1 и G2. Стрелкой указано направление синтеза.
Все этапы синтеза ДНК эукариот осуществляются с помощью репликативного мультиферментного комплекса (праймосомы), состоящего из ферментов и белковых факторов, осуществляющих узнавание точки ori, расплетание родительского дуплекса, удержание материнских цепей ДНК на достаточном расстоянии друг от друга, инициацию новых дочерних цепей, закручивание цепей в спираль и, наконец, терминацию репликации ДНК. В составе комплекса присутствуют новосинтезированная ДНК, не менее 30 белков с Мм от 15 до 300 кДа, ДНК-полимеразы 5, а, праймаза, 3'-5'экзонуклеаза, ДНК-лигаза I, РНКаза H, ДНК-топоизомераза I, ДНК-хеликаза, белок PCNA (ядерный антиген пролиферирующих клеток) и другие факторы.
В праймасоме репликативный белок A (RPA) связывается с однонитевой ДНК и существует в виде комплекса из трех белков с Мм 70, 34 и 11 кДа. Репликативный фактор С (RFC) состоит из пяти белковых елиниц и играет важную роль при связывании репликативных полимераз во время образования репликативной вилки. Белок PCNA, так называемый ДНК-полимеразный зажим, функционирует как кофактор ДНК-полимеразы, образуя закрытое кольцо, окружающее ДНК с каналом в центре. Репликативный фактор С совместно с PCNA продвигается вдоль ДНК по репликативной вилке. С участием праймасомы, ферментов образуются новые полинуклеотидные цепи. Синтез новых цепей всегда идет в направлении от 5'-конца к 3'-концу. Поэтому на одной из ветвей репликативной вилки новая цепь наращивается непрерывно по мере раскручивания ДНК-матрицы, а на другой ветви образуются короткие фрагменты новой цепи ДНК, называемые фрагменты Оказаки; затем концы этих фрагментов соединяются между собой в результате действия фермента ДНК-лигазы (рис. 12.2.).
Рис. 12.2. Схематическое изображение синтеза ДНК в репликативной вилке.
На одной ветви репликативной вилки нуклеотидная цепь синтезируется непрерывно, на другой – прерывисто, с образованием фрагментов Оказаки. Фрагменты Оказаки соединяются между собой в результате действия ДНК-лигазы. По мере роста новых цепей репликативная вилка перемещается по ДНК и в результате происходит репликация (удвоение) молекулы ДНК. Последовательности нуклеотидов новой цепи ДНК строго комплементарны последовательностям родительской цепи, благодаря этому соблюдается передача наследственной информации из поколения в поколение (см. рис.3.5.). Точность репликации очень высока – возможна одна ошибка на 10 трансферазных реакций.
Синтез ДНК на матрице РНК происходит под действием фермента ревертазы или обратной транскриптазы (РНК-зависимой ДНК-полимеразы), содержащейся в составе онковирусов. Доказано, что многие РНК-содержащие онкогенные вирусы (онковирусы) содержат этот фермент в составе покровных белков. При этом механизм синтеза ДНК на матрице РНК включает 3 стадии: на первом этапе фермент ревертаза синтезирует на матрице вирусной РНК комплементарную цепь ДНК, что приводит к формированию гибридной молекулы. Затем исходная молекула РНК отделяется от ДНК и только после этого на матрице ДНК синтезируется комплементарная цепь ДНК.
ЛЕКЦИЯ № 3. Нуклеиновые кислоты. Биосинтез белка
Нуклеиновые кислоты – это фосфорсодержащие биополимеры, мономерами которых являются нуклеотиды. Цепи нуклеиновых кислот включают от нескольких десятков до сотен миллионов нуклеотидов.
Существует 2 вида нуклеиновых кислот – дезоксирибо-нуклеиновая кислота (ДНК) и рибонуклеиновая кислота (РНК). Нуклеотиды, входящие в состав ДНК, содержат углевод, дезокси-рибозу, в состав РНК – рибозу.
Как правило, ДНК представляет собой спираль, состоящую из двух комплиментарных полинуклеотидных цепей, закрученных вправо. В состав нуклеотидов ДНК входят: азотистое основание, дезоксирибоза и остаток фосфорной кислоты. Азотистые основания делят на пуриновые (аденин и гуанин) и пиримидиновые (ти-мин и цитозин). Две цепи нуклеотидов соединяются между собой через азотистые основания по принципу комплементарности: между аденином и тимином возникают две водородные связи, между гуанином и цитозином – три.
1) обеспечивает сохранение и передачу генетической информации от клетки к клетке и от организма к организму, что связано с ее способностью к репликации;
2) регуляция всех процессов, происходящих в клетке, обеспечиваемая способностью к транскрипции с последующей трансляцией.
Процесс самовоспроизведения (авто-репродукции) ДНК называется репликацией. Репликация обеспечивает копирование генетической информации и передачу ее из поколения в поколение, генетическую идентичность дочерних клеток, образующихся в результате митоза, и постоянство числа хромосом при митоти-ческом делении клетки.
РНК – одноцепочечный полимер, в состав мономеров которого входят пуриновые (аденин, гуанин) и пиримидиновые (урацил, цитозин) азотистые основания, углевод рибоза и остаток фосфорной кислоты.
Различают 3 вида РНК: информационную, транспортную и рибо-сомальную.
Информационная РНК (и-РНК) располагается в ядре и цитоплазме клетки, имеет самую длинную полинуклеотидную цепь среди РНК и выполняет функцию переноса наследственной информации из ядра в цитоплазму клетки.
Транспортная РНК (т-РНК) также содержится в ядре и цитоплазме клет-ки, ее цепь имеет наиболее сложную структуру, а также является самой короткой (75 нуклеотидов). Т-РНК доставляет аминокислоты к рибосомам в процессе трансляции – биосинтеза белка.
Рибосомальная РНК (р-РНК) содержится в ядрышке и рибосомах клетки, имеет цепь средней длины. Все виды РНК образуются в процессе транскрипции соответствующих генов ДНК.
3. Биосинтез белка
Биосинтез белка в организме эукариот происходит в несколько этапов.
1. Транскрипция – это процесс синтеза и-РНК на матрице ДНК. Цепи ДНК в области активного гена освобождаются от ги-стонов. Водородные связи между комплементарными азотистыми основаниями разрываются. Основной фермент транскрипции РНК-полимераза присоединяется к промотору – специальному участку ДНК. Транскрипция проходит только с одной (кодоген-ной) цепи ДНК. По мере продвижения РНК-полимеразы по кодо-генной цепи ДНК рибонуклеотиды по принципу комплементарности присоединяются к цепочке ДНК, в результате образуется незрелая про-и-РНК, содержащая как кодирующие, так и некоди-рующие нуклеотидные последовательности.
Транскрипция и процессинг происходят в ядре клетки. Затем зрелая и-РНК через поры в мембране ядра выходит в цитоплазму, и начинается трансляция.
3. Трансляция – это процесс синтеза белка на матрице и РНК. В начале и-РНК 3-концом присоединяется к рибосоме. Т-РНК доставляют к акцепторному участку рибосомы аминокислоты, которые соединяются в полипептидную цепь в соответствии с шифрующими их кодонами. Растущая полипептидная цепь перемещается в донорный участок рибосомы, а на акцепторный участок приходит новая т-РНК с аминокислотой. Трансляция прекращается на кодонах-терминаторах. Генетический код
Это система кодирования последовательности аминокислот белка в виде определенной последовательности нуклеотидов в ДНК и РНК.
Единица генетического кода (кодон) – это триплет нуклеоти-дов в ДНК или РНК, кодирующий одну аминокислоту.
Всего генетический код включает 64 кодона, из них 61 кодирующий и 3 некодирующих (кодоны-терминаторы, свидетельствующие об окончании процесса трансляции).
Кодоны-терминаторы в и-РНК: УАА, УАГ, УГА, в ДНК: АТТ, АТЦ, АЦТ.
Начало процесса трансляции определяет кодон-инициатор (АУГ, в ДНК – ТАЦ), кодирующий аминокислоту метионин. Этот кодон первым входит в рибосому. Впоследствии метионин, если он не предусмотрен в качестве первой аминокислоты данного белка, отщепляется.
Генетический код обладает характерными свойствами.
1. Универсальность – код одинаков для всех организмов. Один и тот же триплет (кодон) в любом организме кодирует одну и ту же аминокислоту.
2. Специфичность – каждый кодон шифрует только одну аминокислоту.
3. Вырожденность – большинство аминокислот могут кодироваться несколькими кодонами. Исключение составляют 2 аминокислоты – метионин и триптофан, имеющие лишь по одному варианту кодона.

Лекция № 12 Биохимия нуклеиновых кислот. Матричные биосинтезы.

1. Структура и функции ДНК (закономерности нуклеотидного состава, структура, гистоны, укладка ДНК в хроматине).

Химическая организация гена А. Строение нуклеозида. Нуклеозиды состоят из азотистых оснований и углевода – пентоза. Название пуриновых нуклеозидов имеют окончание – ОЗИН, а пиримидиновых нуклеозидов окончание – ИДИН. Б. Строение нуклеотидов. Нуклеотиды являются мономерами нуклеиновых кислот они состоят из азотистых оснований углевода – пентоза и фосфорной кислоты. Нуклеотиды хорошо растворимы в воде и обладают кислотными свойствами → нуклеотиды являются кислотами. Разные нуклеотиды отличатся между собой природой углеводов и азотистых оснований.

В. Строение и виды азотистых оснований: Азотистые основания нуклеотидов делятся на 2 типа: 1. Пиримидиновые – они состоят из 1 шестичленного кольца; 2. Пуриновые - состоят из 2 конденсированных 5 и 6-членных колец.
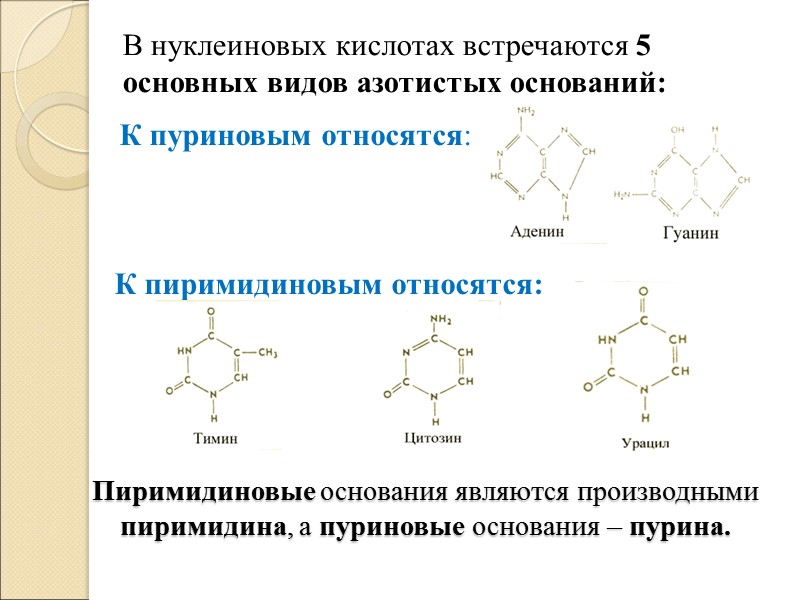
Пиримидиновые основания являются производными пиримидина, а пуриновые основания – пурина. В нуклеиновых кислотах встречаются 5 основных видов азотистых оснований: К пуриновым относятся: К пиримидиновым относятся:
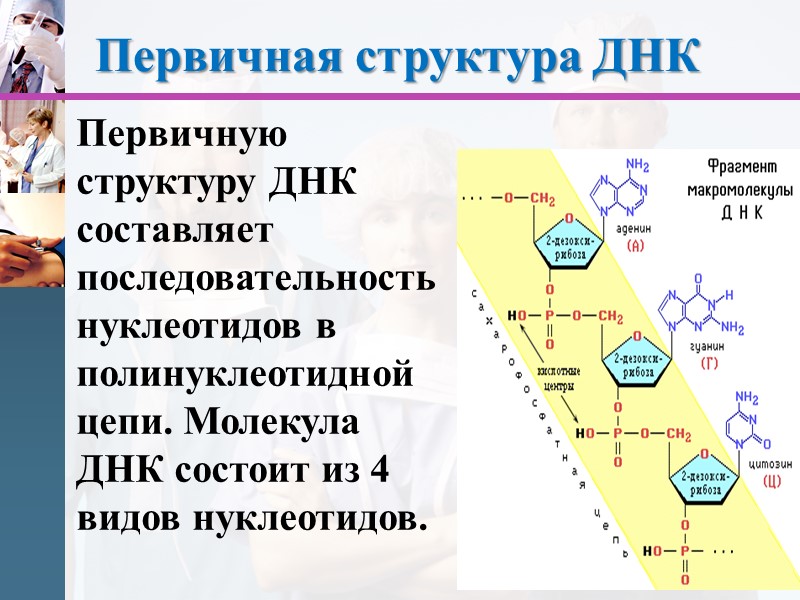
Первичная структура ДНК Первичную структуру ДНК составляет последовательность нуклеотидов в полинуклеотидной цепи. Молекула ДНК состоит из 4 видов нуклеотидов.

Больших успехов в определении структуры ДНКдостигли Эрвин Чаргафф и его сотрудники (1950г.) Используя метод хроматографии, они впервые определили нуклеотидный состав ДНК. Они установили, что соотношение азотистых оснований ДНК подчиняется универсальным. Правила Чаргаффа: 1. Сумма пуриновых нуклеотидов = сумме пиримидиновых нуклеотидов. 2. Молярное содержание аденинов = молярному содержанию тиминов и их отношение =1. 3. Количество аденина = количеству цитозина, а количество гуанина = количеству тимина, сумма аденина и цитозина = сумме гуанина и тимина.

Это супер спираль или кольцо более высокого порядка, представляет собой дальнейшую спирализацию и суперспирализацию молекулы ДНК. Третичная структура ДНК

Хромосомы эукариот представляют собой линейную молекулу ДНК. Эукариотическая ДНК обматывает белковые частицы- гистоны, располагающиеся вдоль ДНК.

2. Репликация ДНК (механизмы, ферменты). Повреждения ДНК. Репарация повреждений и ошибок репликации (механизмы, ферменты).
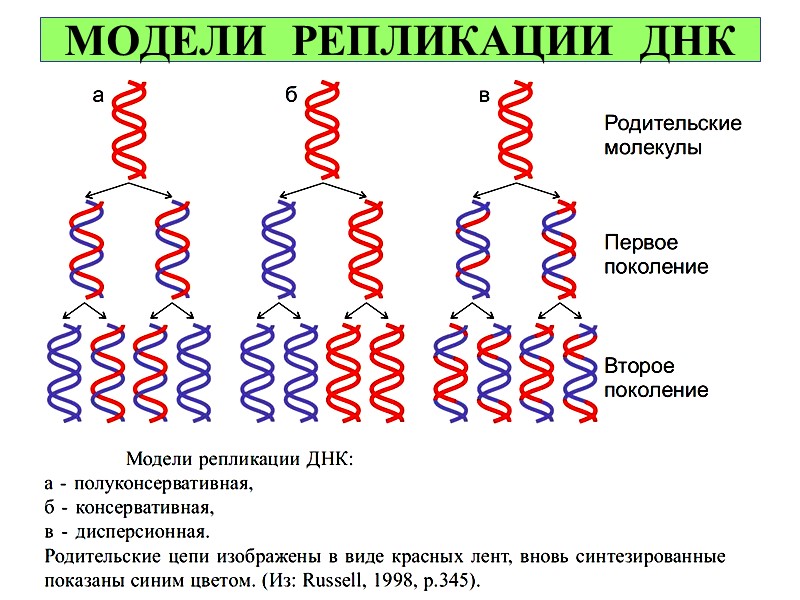
МОДЕЛИ РЕПЛИКАЦИИ ДНК

Репликон Единица, с помощью которой клетка контролирует отдельные акты репликации, получила название репликона. Каждый репликон в каждом клеточном цикле активизируется только один раз. В нем обязательно должны присутствовать необходимые для репликации контролирующие элементы: точка начала (origin), в которой инициируется репликация, точка окончания (terminus), в которой репликация останавливается.

ЭЛОНГАЦИЯ РЕПЛИКАЦИИ РНК-праймер ДНК-хеликаза РНК-праймер РНК-праймер Фрагменты Оказаки Праймаза (ДНК-полимераза α) ДНК-полимераза δ ДНК полимераза ε узнает РНК праймер и начинает синтезировать ДНК SSB-белки Лидирующая ДНК Отстающая ДНК

3. Структура и функции РНК (разновидности, структура). Биосинтез РНК и ее процессинг.

www.themegallery.com Нить ДНК, которая служит матрицей для синтеза РНК при транскрипции называется смысловой или матричной. Вторая нить ДНК называется некодирующей (комплементарная ей РНК не кодирует белки, она "бессмысленная").

Специфическая последовательность ДНК (сайт), в которой РНК-полимераза связывается с матрицей и начинает синтез РНК, называется промотором, а последовательность, на которой завершается синтез РНК, - сайтом терминации. Участок ДНК, ограниченный промотором и сайтом терминации, представляет собой единицу транскрипции -транскриптон. У эукариотов в состав транскриптона, как правило, входит только один ген.

Структура РНК-полимеразы РНК-полимераза — достаточно большая молекула. Основной фермент содержит 5 субъединиц (~400 кДа): α2: две α-субъединицы связывают остальные элементы фермента и распознают регулирующие факторы. Каждая субъединица состоит из двух доменов: αСКД (С-концевой домен) связывает первый элемент промотора, и αNКД (N-концевой домен) связывается с остальными компонентами полимеразы. β: эта субъединица обладает собственно полимеразным действием, катализируя синтез РНК. Она осуществляет инициацию процесса и управляет элонгацией. β': неспецифически связывается с ДНК. ω: восстанавливает денатурированную РНК-полимеразу обратно в дееспособную форму in vitro. Также обнаружено ее защитное/шаперонное действие на β'-субъединицу у Mycobacterium smegmatis. Для связывания с промоторными областями ДНК, основной фермент нуждается в еще одной субъединице — сигма (σ). Сигма-фактор значительно снижает сродство РНК-полимеразы к неспецифичным областям ДНК, и в то же время повышает ее чувствительность к определенным промоторам, в зависимости от своей структуры. С его помощью транскрипция начинается с нужного участка ДНК.

Эукариоты обладают различными типами РНК-полимераз, классифицируемыми по типам РНК, которые они производят: РНК-полимераза I, синтезирующая 45S-предшественника рРНК, превращающуюся затем в рРНК 28S, 18S и 5,8S, которые уже образуют главные РНК-секциирибосомы. РНК-полимераза II, производящая предшественников для мРНК, а также для большинства мяРНК и миРНК. Это наиболее хорошо изученный тип РНК-полимеразы. Ввиду того, что транскрипция должна происходить под строгим контролем, РНК-полимеразе II для связывания с промоторами требуется целый набор факторов транскрипции. РНК-полимераза III, синтезирующая тРНК, 5S рРНК и другие малые РНК, присутствующее в ядре и цитозоле.

Модификации пре-тРНК В процессе посттранскрипционных модификаций первичных транскриптов тРНК: • молекулы укорачиваются с 5'- и 3'-концов и удаляется интрон; • 10-15% азотистых оснований в молекулах модифицируется; • на 3'-конце формируется акцепторный участок (-ССА) для присоединения аминокислот, а в средней части антикодон - триплет нуклеотидов, обеспечивающий взаимодействие тРНК с кодоном мРНК.

Обратная транскрипция Некоторые вирусы (такие как ВИЧ, вызывающий СПИД), имеют возможность транскрибировать РНК в ДНК. ВИЧ имеет РНК-геном, который встраивается в ДНК. В результате, ДНК вируса может быть объединено с геномом клетки-хозяина. Главный фермент, ответственный за синтез ДНК из РНК, называется ревертазой. Одной из функций ревертазы является создание комплементарной ДНК (кДНК) из вирусного генома. Ассоциированый фермент рибонуклеаза H расщепляет РНК, а ревертаза синтезирует кДНК из двойной спирали ДНК. кДНК интегрируется в геном клетки-хозяина с помощью интегразы. Результатом является синтез вирусных протеинов клеткой-хозяином, которые образуют новые вирусы. В случае с ВИЧ так же программируется апоптоз (смерть клетки) Т-лимфоцитов. В иных случаях клетка может остаться распростанителем вирусов. Некоторые клетки эукариотов содержат фермент теломеразу, так же проявляющую активность обратной транскрипции. С её помощью синтезируются повторяющиеся последовательности в ДНК. Теломераза часто активирутся в раковых клетках для бесконечной дупликации генома без потери кодирующей протеины последовательности ДНК.

Классификация интронов Существует четыре группы интронов: Ядерные интроны Интроны группы I Интроны группы II Интроны группы III Иногда интроны группы III также относят к группе II, потому что они похожи по структуре и функции. Ядерные, или спалайсосомные интроны подвергаются сплайсингу при помощи сплайсосомы и малых ядерных РНК (мяРНК, snRNA). В последовательности РНК, содержащей ядерные интроны, есть специальные сигнальные последовательности, которые узнаются сплайсосомой. Интроны I, II и III группы способны к автосплайсингу и встречаются реже, чем сплайсосомные интроны.

4. Обмен нуклеотидов (состав, биосинтез, катаболизм). Нарушение обмена азотистых оснований (подагра).

Метаболизм нуклеотидов

ФУНКЦИИ НУКЛЕОТИДОВ

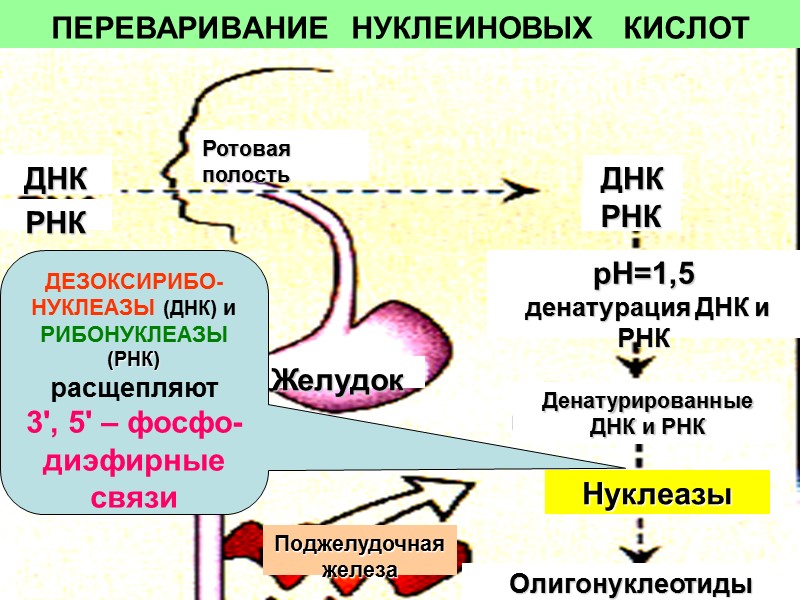
ПЕРЕВАРИВАНИЕ НУКЛЕИНОВЫХ КИСЛОТ Ротовая полость ДНК РНК ДНК РНК Желудок Поджелудочная железа рН=1,5 денатурация ДНК и РНК Денатурированные ДНК и РНК Нуклеазы Олигонуклеотиды ДЕЗОКСИРИБО-НУКЛЕАЗЫ (ДНК) и РИБОНУКЛЕАЗЫ (РНК) расщепляют 3', 5' – фосфо-диэфирные связи

Поджелудочная железа Олигонуклеотиды Фосфодиэстеразы Мононуклеотиды Нуклеотидазы Нуклеозиды Нуклеозидазы Пиримидины Пурины (Дезокси) рибоза Кровоток Мочевая кислота Моча Тонкий кишечник Ф Энтероцит

МЕТАБОЛИЗМ ПУРИНОВ

ИСТОЧНИКИ АТОМОВ В ПУРИНОВЫХ ОСНОВАНИЯХ АСПАРТАТ ГЛИЦИН ГЛУТАМИН ФОРМИЛ ТГФК СЕРИН

Особенность синтеза пуринов Атомы С и N поэтапно добавляются к рибозе 5-фосфат.

Синтез ФРПФ (ФРДФ) Активатор Ингибиторы Пуриновые нуклеотиды Рибозо-5-фосфат 5-фосфорибозил-1-пирофосфат (ФРПФ) АТФ АМФ ФРПФ синтетаза

ЛИМИТИРУЮЩАЯ РЕАКЦИЯ 5-фосфорибозиламин Глутамин Глутамат Ингибиторы АМФ, ГМФ, ИМФ 5-фосфорибозил 1-пирофосфат (ФРПФ) Глутамин фосфорибозил пирофосфат амидо- трансфераза [S] 3.

Полисома Одновременно несколько рибосом могут участвовать в трансляции одной мРНК. Каждая рибосома занимает участок, равный примерно 80 нуклеотидам мРНК. Таким образом, рибосомы располагаются на мРНК с интервалами около 100 нуклеотидов, образуя комплекс, называемый полисомой.

Функционально активные белки образуются в результате посттрансляционных модификаций полипептидных цепей, синтезированных на рибосомах. Они включают: частичный протеолиз; фолдинг, или формирование пространственной структуры, в котором принимают участие белки-шапероны, обеспечивающие образование функционально активной конформации полипептидной цепи; модификации аминокислот: карбоксилирование, фосфорилирование, йодирование, гидроксилирование, ацилирование и гликозилирование; образование дисульфидных связей между остатками цистеина, участвующими в формировании трехмерной структуры белка; присоединение простетических групп; образование олигомерных структур, которое также осуществляется при участии шаперонов.

Структурная часть гена Промотор Начало транскрипции Старт-кодон Стоп - кодон 5’ 3’ 3’ 5’ Терминатор транскрипции Строение гена Участок связывания фермента РНК-полимеразы (место начала транскрипции) Участок, кодирующий последовательность аминокислот в молекуле белка. Триплет ТАЦ в ДНК и АУГ в РНК иницирующий начало синтеза белка Триплеты – бессмысленные кодоны на которых завершается трансляция Участок, сигнализирующий о прекращении транскрипции

Виды оперонов Оперон — это тесно связанная последовательность структурных генов, определяющих синтез группы белков, которые участвуют в одной цепи биохимических преобразований. К наиболее хорошо изученным оперонам бактерий относят: Лактозный (lac)-оперон Галактозный (gal)-оперон Триптофановый (trp)-оперон

Лактозный оперон Оперон – участок бактериальной хромосомы, включающий следующие участки ДНК: Р – промотор, О – оператор, Z, Y, А – структурные гены, Т –терминатор. (В состав других оперонов может входить до 10 структурных генов.) Промотор – место присоединения РНК- полимеразы. Оператор – участок оперона, к которому присоединяются белки-репрессоры или активаторы транскрипции. Структурные гены – гены кодирующие ферменты, участвующие в метаболизме лактозы. Терминатор служит для отсоединения РНК-полимеразы после окончания синтеза иРНК, соответствующей ферментам Z, Y, А, необходимым для усвоения лактозы.
Читайте также: