
Биологическая фиксация молекулярного азота кратко
Обновлено: 02.07.2024
Некоторые бактерии способны фиксировать атмосферный (молекулярный) азот, т. е. переводить его в связанное состояние. Они восстанавливают азот до аммиака; часть его используется самими микроорганизмами, а часть выделяется в окружающую среду.
Одни азотфиксирующие (азотусваивающие) бактерии живут свободно в почве и воде; другие – в симбиотическом сожительстве с растениями, преимущественно бобовыми. Бактерии поселяются в бородавчатых вздутиях – клубеньках корней этих растений. Отсюда произошло и название этих бактерий – клубеньковые. Энергию, необходимую для фиксации бактерии получают в процессе окисления безазотистых органических соединений, которые они берут из клеток корней растений.
Величина и форма клубеньковых бактерий значительно изменяются в зависимости от их возраста и условий жизни. Молодые клетки – мелкие подвижные палочки – не образуют спор. По мере развития клетки теряют жгутики, становятся искривленными, утолщенными или ветвистыми; эти формы клубеньковых бактерий называются бактероидами.
Среди свободно живущих азотфиксирующих бактерий наибольшее значение имеет аэробная бактерия Azotobacter chroocaccum, имеющая форму слегка приплюснутых кокков, часто объединенных попарно; клетки имеют слизистую капсулу.
Из анаэробных свободно живущих азотусваивающих бактерии следует отметить бактерию, открытую С. Н. Виноградским (1893 г.), – Clostndium pasteurianum. Это подвижные спорообразующие палочки, способные сбраживать углеводы по типу маслянокислого брожения, которое и служит бактериям источником энергии для связывания молекулярного азота.
Азотфиксирующие бактерии имеют важное значение для сельского хозяйства. За счет их деятельности постоянно пополняются азотистые запасы почвы, что способствует ее плодородию.
В практике сельского хозяйства препараты из азотфиксирующих бактерий используются в качестве бактериального удобрения: азотобактерин – из культур азотобактера, нитрагин – из культур клубеньковых бактерий.
Свободноживущие азотфиксирующие микроорганизмы:
Азотобактер (Azotobacter). В 1901 году Бейеринк выделил из почвы аэробную неспорообразующую грамотрицательную бактерию, фиксирующую молекулярный азот, и назвал ее Azotobacter chroococcum (в родовом названии отражена способность бактерии фиксировать азот, в видовом – способность синтезировать коричневый пигмент – chroo и образовывать кокковидные клетки – coccum). Азотобактер – типичный представитель свободноживущих микроорганизмов. Свободноживущие – это все те микроорганизмы, которые живут в почве независимо от того, развивается вблизи растение или нет.
Бейеринкия (Beijerinckia) Впервые аэробные бактерии рода Beijerinckia были выделены из кислых почв рисовых полей в Индии (в 1939 г.). Г. Деркс (1950), обнаружив эту бактерию в почве Ботанического сада в Богоре (Ява), предложил назвать ее именем М. Бейеринка – одного из первых исследователей фиксаторов азота.
Клостридиум (Clostridium) Первый анаэробный микроорганизм, усваивающий молекулярный азот, был выделен и описан С. Н. Виноградским в 1893 г. Он оказался спорообразующей бактерией, которой было дано наименование Clostridium pasteurianum (родовое название происходит от латинского слова clostrum – веретено; видовое – pasteurianum – дано в честь Луи Пастера).
Сейчас известно свыше 80 видов и разновидностей бактерий, несколько видов актиномицетов, дрожжей, дрожжеподобных организмов и плесневых грибов, способных фиксировать азот. Они населяют почву, дно морей и пресных водоемов.
Остановимся на характеристике лишь основных представителей бактерий.
Способность фиксировать азот присуща ряду представителей семейства Pseudomonadaceae. Они довольно широко распространены в природе. К наиболее интересным представителям этого семейства относится несколько видов: Azotomonas insolita, фиксирующий до 12 мг азота на 1 г использованного сахара и встречающийся нередко не только в почве, но и как эпифит на растениях; Azotomonas fluorescens, выделенный впервые из компостов Н. А. Красильниковым (1945), продуктивность азотфиксации которого не меньше, чем у первого вида; Pseudomonas azotocolligans, населяющий кислые и щелочные почвы Америки; Pseudomonas azotogensis, широко распространенный в почвах Канады и легко выделяющийся из парниковых почв, и, наконец, Pseudomonas methanitrificens, который встречается в почвах, имеющих выход источников природных газов. Последняя бактерия особенно интересна тем, что она использует метан и другие газообразные углеводороды в качестве единственного источника углерода, за счет энергии разложения которых она усваивает азот атмосферы.
Представители семейства Spirillaceae, фиксирующие молекулярный азот, распространены преимущественно в рыбоводных прудах, озерах, морской воде, морских отложениях. Возможно, они играют немаловажную роль в фиксации азота в водоемах.
Если вам нужна помощь в написании работы, то рекомендуем обратиться к профессионалам. Более 70 000 авторов готовы помочь вам прямо сейчас. Бесплатные корректировки и доработки. Узнайте стоимость своей работы.
Не только азотобактеру, как члену семейства Azotobacteriaceae, свойственна азотфиксирующая функция. В почвах Индии встречаются еще два представителя этого семейства — Derxia gummosa и Derxia indica — активные, хотя и медленнодействующие, азотфиксаторы. В азотном балансе почв они, как и бактерии Agrobacterium radiobacter из семейства Rhizobiaceae, по-видимому, не играют сколько-нибудь значительной роли. Молекулярному азоту они предпочитают связанные источники азота, такие, как мочевина, пептон, аминокислоты и минеральные соединения.
Среди спорообразующих грамположительных бактерий семейства Bacillaceae азотфиксирующая способность выявлена у факультативных анаэробов Bacillus polymyxa, аэробов Bacillus megaterium и Thermobacillus azotofigens. Последняя бактерия, выделенная из удобренной навозом дерново-карбонатной почвы Эстонской ССР, оказалась термофильной с оптимумом роста 45-50°С и максимумом 60-65°С. При температурах ниже 20°С она не развивается.
Распространение функции азотфиксации в ряде семейств фотосинтезирующих бактерий (семейства Thiorhodaceae, Athiorhodaceae, Chlorobacteriaceae, Hyphomicrobiaceae) не случайно, так как, по-видимому, они являются представителями одной из древнейших групп азотфиксаторов на Земле.
Небольшие количества молекулярного азота способны усваивать почвенные микобактерии. Усвоенный микобактериями азот в сочетании с азотом, ассимилированным другими олигонитрофильными микроорганизмами, сине-зелеными водорослями, лишайниками и мхами, настолько обогащает примитивную почву азотом, что ее могут заселять высшие растения.
Биологическая фиксация азота атмосферы имеет важное значение. Об этом свидетельствуют масштабы процесса — до 200 млн т N/год. Благодаря биологической фиксации азот переходит в формы, которые могут использовать все растительные, а через них и животные организмы.
Организмы, способные к усвоению азота воздуха, можно разделить на группы:
1) симбиотические азотфиксаторы — микроорганизмы, которые усваивают азот атмосферы, только находясь в симбиозе с высшим растением;
2) не симбиотические азотфиксаторы — микроорганизмы, свободно живущие в почве и усваивающие азот воздуха;
3) ассоциативные азотфиксаторы — микроорганизмы, обитающие на поверхности корневой системы злаков, т. е. живущие в ассоциации с высшими растениями.
Важное значение имеют симбиотические азотфиксаторы, живущие в клубеньках корней бобовых растений (клубеньковые бактерии), относящиеся к роду Rhizobium. Связывание азота атмосферы возможно только при симбиотической ассоциации микроорганизмов этого вида и высшего растения в основном из семейства Бобовые. Существует большое количество разновидностей (штаммов) клубеньковых бактерий, каждая из которых приспособлена к заражению одного или нескольких видов бобовых растений. Это отражается в их названиях: Rhizobium lupini — клубеньковые бактерии люпина и Rhizobium trifolii— клубеньковые бактерии клевера и т. д..
Корневые системы бобовых растений обладают специфическими корневыми выделениями. Благодаря этому клубеньковые бактерии скапливаются вокруг корневых волосков, которые при этом скручиваются. Такая способность организмов передвигаться в ответ на узнавание химических продуктов, называется хемотаксисом. В осуществлении контактного взаимодействия микроорганизмов С растением важное значение имеет так называемое лектину-глеводное узнавание растения микроорганизмом. Суть этого в том, что лектин корневых волосков растений прочно связывается с углеводом поверхности бактерий. Бактерии, внедрившиеся в корневой волосок, в виде сплошного тяжа (т. н. инфекционные нити), состоящего из соединенных слизью бесчисленных бактерий, проникают в паренхиму корня. Клетки перицикла начинают усиленно делиться. Возможно, бактерии выделяют гормональные вещества типа ауксина и именно это является причиной разрастания тканей, образуются вздутия — клубеньки. Клетки клубеньков заполняются быстро размножающимися бактериями, но остаются живыми и сохраняют крупные ядра. Бактерии при этом трансформируются сами, увеличиваются в размерах, поэтому их называют бактероиды.Клубеньковые бактерии заражают только полиплоидные клетки корня. Ткань к пубеньков, заполненная бактериями, приобретает розовую окраску, так как поте заражения в клетках бактерий образуется пигмент, сходный с гемоглобином, — леггемоглобин. Этот пигмент связывает кислород воздуха и тем самым предохраняет фермент нитрогеназу от воздействия кислорода. Исследования показали прямую зависимость между содержанием леггемоглобина и скоростью фиксации азота. При отсутствии леггемоглобина азот не усваивается. Информация об образовании леггемоглобина содержится в ДНК ядра клетки высшего растения. Синтезируется клетками растения-хозяина. Однако он образуется после их заражения. Гены растений, кодирующие образование клубеньков, носят название nod-GENE (нодулин-гены). Показано, что скопление бактерий вокруг корня вызывает выделение веществ (возможно олигосахаров), которые активируют т. н. нодулин-белок, индуцирующий транскрипцию нодулин-генов. Взаимоотношения между высшими растениями и клубеньковыми бактериями обычно характеризуют как симбиоз. Однако на первых этапах заражения бактерии питаются целиком за счет высшего растения, т. е. практически паразитируют на нем. В этот период рост зараженных растений даже несколько тормозится.
В дальнейшем азотфиксирующая способность бактерий увеличивается, и они начинают снабжать азотистыми веществами растение-хозяина, вместе с тем бактерии получают от высшего растения углеводы (симбиоз). По мере дальнейшего развития наступает этап, когда высшее растение паразитирует на клетках бактерий, потребляя все образующиеся там азотистые соединения. В этот период часто наблюдается растворение (лизис) бактериальных клеток.
Благодаря деятельности клубеньковых бактерий часть азотистых соединений из корней бобовых растений диффундирует в почву, обогащая ее азотом. Посев бобовых растений ведет к повышению почвенного плодородия. Гектар бобовых растений в симбиозе с бактериями может перевести в связанное состояние от 100 до 400 кг азота за год. Значение этого трудно переоценить, если учесть, что азотные удобрения наиболее дорогостоящи, а в почве соединения азота содержатся в небольших количествах. Существуют и другие виды высших растений, у которых наблюдается симбиоз с микроорганизмами. Так, маленький водный папоротник азолла (Azolla) находится в симбиотических отношениях с азотфиксирующими цианобактериями. Азолла способна фиксировать до 0,5 кг азота на га в сутки. Некоторые деревья и кустарники (например, ольха, облепиха, лох) имеют в качестве симбионтов бактерии из рода актиномицеты. Большое значение имеют свободноживущие бактерии — азотфиксаторы. В 1893 г. русским микробиологом С.Н. Виноградским была выделена анаэробная азотфиксирующая бактерия Clostridium pasteurianum. В 1901 г. голландский ученый М. Бейеринк выделил две аэробные азотфиксирующие бактерии — Azotobacter chroococum, Azotobacter agile. Сейчас известен ряд видов Azotobacter. Свободноживущие азотфиксаторы могут быть факультативными аэробными или факультативными анаэробными. Для того чтобы эти микроорганизмы осуществляли процесс фиксации азота, необходимо присутствие молибдена, железа и кальция. Особенно важно присутствие молибдена. Свободно живущие азотфиксаторы Ассоциативные азотфиксаторы были обнаружены в 70—80-х годах XX в. в лаборатории Д. Доберейнер в Бразилии (1976). Число их видов велико, как велико разнообразие ассоциативных взаимоотношений растений с микроорганизмами. Такие отношения характерны для ризосферных микроорганизмов, т. е. живущих на поверхности корневой системы растений. Часто микробиологи не делают различия между ассоциативными и свободноживущими азотфиксаторами. Последовательность взаимоотношений с растением-хозяином ассоциативных азотфиксаторов имеет определенное сходство с симбиотическими организмами: хемотаксическое узнавание, лектин-углеводное узнавание и этап установления прочных связей. Отсутствует только этап образования клубеньков. Эффективность азотфиксации ассоциативной микрофлорой меньше по сравнению с симбиотической, но ассоциативные азотфиксаторы продуцируют гормоны роста растений и обладают другими свойствами, положительно влияющими на рост и развитие растений (защита от фитопатогенов, разрушение токсических веществ). Наиболее изучены из этой группы микроорганизмы из рода азоспирилл (Azospirillum). Они колонизируют корни злаков и в связи с этим представляет интерес технология их выращивания. Азоспириллы легко инфицируют корневую систему злаков и других растений. Поможем написать любую работу на аналогичную тему Фиксация молекулярного азота. Свободноживущие и симбиотические азотофиксирующие микроорганизмы. Фиксация молекулярного азота. Свободноживущие и симбиотические азотофиксирующие микроорганизмы. Как известно, атомы в молекуле азота (N2) соединены тремя ковалентными связями, энергия диссоциации которых равна 9,4 • 105 дж/молъ. Наибольшей энергией — 5,3-105 дж/молъ — обладает первая связь, вторая — 2,5-105 дж/молъ, третья — 1,6 • 105 джмолъ. Эти связи очень прочны, и молекулы азота поэтому химически инертны. Именно инертность азота обусловливает необходимость его активации, так же как и повышения химической активности вступающего с ним в реакцию соединения или элемента. Синтез аммиака из атмосферного азота химическим путем, осуществляемый методом Габе-ра — Боша, происходит при температуре 400— 500 °С и высоком давлении (200—1000 атм). Клетки микроорганизмов проводят процесс азотфиксации в обычных условиях. Наиболее вероятно, что разрыв связей в молекуле азота происходит не сразу, а последовательно, в результате действия ферментных систем. В. Е.Шилов и Г.И.Лихтенштейн (1971) предложили интересный механизм активации азота ферментом нитрогеназой (рис. 179). Молекулярный азот через щель размером 4—5 А (это соответствует длине молекулы азота) поступает внутрь фермента и здесь активируется. В его активации большая роль принадлежит молибдене- и железосодержащим центрам, по цепям атомов которых происходит эстафетная передача активирующих азот электронов от восстановителя. Реакции активации способствует близость группировок серы. В результате активации и хемосорбции азота нитрогеназой высвобождается энергия и происходит разрыв двух связей в молекуле азота. На это расходуется 7,8*105 дж/моль. Третья связь разрывается при внедрении в молекулу активированного азота: водорода, активированного, в свою очередь, ферментами дегидрогена-зной и гидрогеназной систем. Получающийся аммиак или другие восстановленные продукты азота реагируют с кетокислотами, образуя аминокислоты. В симбиотических азотфиксирующих системах фиксация азота осуществляется в результате сложного взаимодействия микроорганизма и высшего растения. Несомненно, механизм процесса азотфиксации в клубеньках имеет своеобразные особенности, обусловленные совместным функционированием двух совершенно различных по своим свойствам партнеров, присутствием леггемоглобина, принимающего участие в процессе фиксации, и наличием бактероидов. Процесс фиксации в симбиотической системе иллюстрируется схемой (рис. 180), где видно, что растение и бактерии функционируют в единстве и что электрон-транспортной цепью между ними является леггемоглобин. Высшее растение по схеме является источником углеродсодержащих соединений. Их трансформация обеспечивает получение энергетического материала для процессов активации и восстановления N2. Активированный азот — конечный акцептор электронов. Продукты неполного окисления углеродсодержащих соединений служат акцепторами NHs и в клубеньках образуют аминокислоты, которые становятся доступными высшему растению. Растения выполняют роль накопителя углеродсодержащих соединений (продуктов фотосинтеза) и поставщика энергии. Клубеньковые бактерии в стадии бактероидов проявляют способность с помощью нитрогеназы переносить активированный водород к азоту. Путь от Na до NHs рассматривается как восстановительный процесс. В последнее время получены доказательства локализации процесса азотфиксации в бактероидах с помощью измерения азотфиксирую-щей активности у фракции бактероидов, которую отделяли центрифугированием от других фракций клубенька. Как известно, атомы в молекуле азота (N2) соединены тремя ковалентными связями, энергия диссоциации которых равна 9,4 • 105 дж/молъ. Наибольшей энергией — 5,3-105 дж/молъ — обладает первая связь, вторая — 2,5-105 дж/молъ, третья — 1,6 • 105 джмолъ. Эти связи очень прочны, и молекулы азота поэтому химически инертны. Именно инертность азота обусловливает необходимость его активации, так же как и повышения химической активности вступающего с ним в реакцию соединения или элемента. Синтез аммиака из атмосферного азота химическим путем, осуществляемый методом Габе-ра — Боша, происходит при температуре 400— 500 °С и высоком давлении (200—1000 атм). Клетки микроорганизмов проводят процесс азотфиксации в обычных условиях. Наиболее вероятно, что разрыв связей в молекуле азота происходит не сразу, а последовательно, в результате действия ферментных систем. В. Е.Шилов и Г.И.Лихтенштейн (1971) предложили интересный механизм активации азота ферментом нитрогеназой (рис. 179). Молекулярный азот через щель размером 4—5 А (это соответствует длине молекулы азота) поступает внутрь фермента и здесь активируется. В его активации большая роль принадлежит молибдене- и железосодержащим центрам, по цепям атомов которых происходит эстафетная передача активирующих азот электронов от восстановителя. Реакции активации способствует близость группировок серы. В результате активации и хемосорбции азота нитрогеназой высвобождается энергия и происходит разрыв двух связей в молекуле азота. На это расходуется 7,8*105 дж/моль. Третья связь разрывается при внедрении в молекулу активированного азота: водорода, активированного, в свою очередь, ферментами дегидрогена-зной и гидрогеназной систем. Получающийся аммиак или другие восстановленные продукты азота реагируют с кетокислотами, образуя аминокислоты. В симбиотических азотфиксирующих системах фиксация азота осуществляется в результате сложного взаимодействия микроорганизма и высшего растения. Несомненно, механизм процесса азотфиксации в клубеньках имеет своеобразные особенности, обусловленные совместным функционированием двух совершенно различных по своим свойствам партнеров, присутствием леггемоглобина, принимающего участие в процессе фиксации, и наличием бактероидов. Процесс фиксации в симбиотической системе иллюстрируется схемой (рис. 180), где видно, что растение и бактерии функционируют в единстве и что электрон-транспортной цепью между ними является леггемоглобин. Высшее растение по схеме является источником углеродсодержащих соединений. Их трансформация обеспечивает получение энергетического материала для процессов активации и восстановления N2. Активированный азот — конечный акцептор электронов. Продукты неполного окисления углеродсодержащих соединений служат акцепторами NHs и в клубеньках образуют аминокислоты, которые становятся доступными высшему растению. Растения выполняют роль накопителя углеродсодержащих соединений (продуктов фотосинтеза) и поставщика энергии. Клубеньковые бактерии в стадии бактероидов проявляют способность с помощью нитрогеназы переносить активированный водород к азоту. Путь от Na до NHs рассматривается как восстановительный процесс. В последнее время получены доказательства локализации процесса азотфиксации в бактероидах с помощью измерения азотфиксирую-щей активности у фракции бактероидов, которую отделяли центрифугированием от других фракций клубенька. Фиксация азота это процесс, посредством которого молекулярные азот в воздуха превращается в аммиак ( NH Фиксация азота необходима для жизни, поскольку фиксированные неорганические соединения азота необходимы для биосинтез всех азотсодержащих органические соединения, Такие как аминокислоты и белки, нуклеозидтрифосфаты и нуклеиновые кислоты. В рамках азотный цикл, это важно для сельское хозяйство и изготовление удобрение. Это также косвенно связано с производством всех химических соединений азота, включая некоторые взрывчатые вещества, фармацевтические препараты и красители. Фиксация азота осуществляется естественным путем в почва к микроорганизмы названный диазотрофы которые включают бактерии Такие как Азотобактер и археи. Некоторые азотфиксирующие бактерии имеют симбиотический отношения с группами растений, особенно бобовые. [2] Более слабые несимбиотические отношения между диазотрофами и растениями часто называют ассоциативными, что проявляется в азотфиксации на рис корни. Фиксация азота происходит между некоторыми термиты и грибы. [3] Это происходит в воздухе естественным путем за счет НЕТИкс производство молния. [4] [5] Все биологические реакции, связанные с азотфиксацией, катализируются ферментами, называемыми нитрогеназы. [6] Эти ферменты содержат утюг, часто со вторым металлом, обычно молибден но иногда ванадий. Азот можно зафиксировать молния который преобразует газообразный азот ( N Немецкий агроном открыл биологическую азотфиксацию. Германн Хеллригель [10] и голландский микробиолог Мартинус Бейеринк. [11] Биологическая фиксация азота (BNF) происходит, когда атмосферный азот превращается в аммиак посредством нитрогеназа фермент. [1] Общая реакция на BNF: Этот процесс связан с гидролиз 16 эквивалентов АТФ и сопровождается сообразованием одного эквивалента ЧАС Нитрогеназы быстро разлагаются кислородом. По этой причине многие бактерии прекращают производство фермента в присутствии кислорода. Многие азотфиксирующие организмы существуют только в анаэробный условия, дыхание для снижения уровня кислорода или связывание кислорода с белок Такие как леггемоглобин. [1] Диазотрофы широко распространены внутри домена Бактерии включая цианобактерии (например, очень важные Триходесмий и Cyanothece), а также зеленые серные бактерии, Азотобактерии, ризобия и Frankia. Некоторые облигатно анаэробные бактерии фиксируют азот, в том числе многие (но не все) Clostridium виды Немного археи также фиксируют азот, в том числе несколько метаногенный таксоны, которые вносят существенный вклад в фиксацию азота в почвах с дефицитом кислорода. [16] Цианобактерии населяют почти все освещенные среды на Земле и играют ключевую роль в углероде и азотный цикл из биосфера. Как правило, цианобактерии могут использовать различные неорганические и органические источники комбинированного азота, такие как нитрат, нитрит, аммоний, мочевина, или несколько аминокислоты. Некоторые штаммы цианобактерий также способны к диазотрофному росту - способность, которая могла присутствовать у их последнего общего предка в Архейский эон. [17] Фиксация азота цианобактериями в коралловые рифы может фиксировать вдвое больше азота, чем на суше - около 660 кг /ха/год. Колониальные морские цианобактерии Триходесмий считается, что он фиксирует азот в таком масштабе, что на него приходится почти половина фиксации азота в морских системах во всем мире. [18] Морские поверхностные лишайники и нефотосинтезирующие бактерии, принадлежащие к Proteobacteria и Planctomycetes, в значительной степени фиксируют атмосферный азот. [19] Растения, способствующие азотфиксации, включают растения бобовые семья—Fabaceae- с таксоны Такие как кудзу, клевер, соя, люцерна, люпин, арахис и Ройбуш. В них содержатся симбиотический ризобия бактерии внутри узелки в их корневые системы, производя азотные соединения, которые помогают растению расти и конкурировать с другими растениями. [20] Когда растение умирает, фиксированный азот высвобождается, делая его доступным для других растений; это помогает удобрять почва. [1] [21] У подавляющего большинства бобовых есть эта ассоциация, но некоторые роды (например., Стифнолобий) не. Во многих традиционных методах ведения сельского хозяйства поля повернутый через различные типы культур, которые обычно включают растение, состоящее в основном или полностью из клевер. [ нужна цитата ] Эффективность фиксации в почве зависит от многих факторов, в том числе от бобовые и состояние воздуха и почвы. Например, фиксация азота красным клевером может составлять от 50 до 200 фунтов на акр. [22] Другие семейства азотфиксаторов включают: Способность фиксировать азот присутствует и у других семейств, принадлежащих к заказы Тыквенные, Fagales и Росалес, который вместе с Fabales сформировать кладу евросиды. Способность фиксировать азот у этих семейств не всегда. Например, из 122 Розоцветные родов только четыре фиксируют азот. Fabales были первой линией, ответвленной от этой азотфиксирующей клады; таким образом, способность фиксировать азот может быть плезиоморфный и впоследствии утерян у большинства потомков исходного азотфиксирующего растения; однако может оказаться, что основной генетический и физиологический требования присутствовали в начальном состоянии в самые недавние общие предки всех этих растений, но развились до полноценной функции только у некоторых из них. Несколько азотфиксирующих симбиотических ассоциаций включают: цианобактерии (Такие как Носток): Ропалодия гибба, а диатомовые водоросли водоросль, это эукариот с цианобактериями N Некоторые ученые работают над внедрением генов, отвечающих за фиксацию азота, непосредственно в ДНК растений. Поскольку все известные примеры фиксации азота имеют место у прокариот, передача функциональности эукариотам, таким как растения, является проблемой; одна команда использует дрожжи в качестве своего эукариотического тестового организма. Основная проблема, которую необходимо преодолеть, - это чувствительность продуцируемых ферментов к кислороду, а также потребность в энергии. Рассматривается возможность того, что процесс протекает внутри митокондрий или хлоропластов. [28] Возможность того, что атмосферный азот вступает в реакцию с некоторыми химическими веществами, впервые была обнаружена Desfosses в 1828 г. Он заметил, что смеси щелочной металл оксиды и углерод вступают в реакцию с азотом при высоких температурах. С использованием карбонат бария В качестве исходного материала в 1860-х годах стал доступен первый коммерческий процесс, разработанный Маргаритой и Сурдеваль. Результирующий цианид бария может реагировать с паром с образованием аммиака. В 1898 г. откровенный и Каро развязал процесс и произвел карбид кальция и на последующей стадии реагировал с азотом до цианамид кальция. В Оствальдский процесс для производства азотная кислота был открыт в 1902 году. Франк-Каро и процессы Оствальда преобладали в промышленной фиксации до открытия Процесс Габера в 1909 г. [31] [32] Самый распространенный метод производства аммиака - это Процесс Габера. Производство удобрений в настоящее время является крупнейшим источником фиксированного азота, производимого людьми на суше. экосистема. Аммиак - необходимый прекурсор для удобрения, взрывчатка, и другие продукты. Процесс Габера требует высокого давления (около 200 атм) и высоких температур (не менее 400 ° C), которые являются обычными условиями для промышленного катализа. В этом процессе в качестве источника водорода используется природный газ, а в качестве источника азота - воздух. [33] Было проведено много исследований по открытию катализаторов фиксации азота, часто с целью снижения потребности в энергии. Однако такие исследования до сих пор не смогли приблизиться к эффективности и простоте процесса Габера. Многие соединения реагируют с атмосферным азотом с образованием диазотные комплексы. Первый диазот сложный быть сообщенным было Ru (NH Достижение каталитической химической фиксации азота в условиях окружающей среды является постоянной научной задачей. На примере нитрогеназы продолжается работа в этой области гомогенного катализа с особым упором на гидрирование. [35] Металлический литий горит в атмосфере азота, а затем превращается в нитрид лития. Гидролиз образовавшегося нитрида дает аммиак. В связанном процессе триметилсилилхлорид, литий и азот реагируют в присутствии катализатора с образованием трис (триметилсилил) амин. Затем это можно использовать для реакции с α, δ, ω-трикетоны дать трициклический пирролы. [36] Однако процессы с участием металлического лития не представляют практического интереса, поскольку они не являются каталитическими и восстанавливают Ли + Начиная с 1960-х годов было выявлено несколько гомогенных систем, которые превращают азот в аммиак, иногда каталитически, но часто действуя через плохо определенные механизмы. Оригинальное открытие описано в раннем обзоре: «Вольпин и его сотрудники, используя непротонную кислоту Льюиса, трибромид алюминия, смогли продемонстрировать истинный каталитический эффект титана, обработав диазот смесью тетрахлорида титана, металлического алюминия и трибромида алюминия при 50 ° C. , либо в отсутствие, либо в присутствии растворителя, например бензола. До 200 моль аммиака на моль TiCl Поиск четко определенных промежуточных продуктов привел к характеристике многих диазотные комплексы переходных металлов. Хотя некоторые из этих четко определенных комплексов действуют каталитически, их поведение указывает на вероятные стадии азотфиксации. Первые плодотворные исследования были сосредоточены на (MN 20− В 2003 году было обнаружено, что амидокомплекс молибдена катализирует восстановление N В 2011 году была открыта еще одна система на основе молибдена, но с дифосфором. клещевой лиганд. [44] Фотолитический расщепление азота также учитывается. [45] [46] [47] [48] [49] Фиксация азота на p-блок элемент был опубликован в 2018 году, согласно которому одна молекула диазота связана двумя временными База Льюиса-стабилизированный борилен разновидность. [50] Результирующий дианион впоследствии был окисленный до нейтрального соединения и восстанавливается водой. С помощью катализа и энергии электричества и света, NH По состоянию на 2019 год в исследовании рассматривались альтернативные способы подачи азота в сельское хозяйство. Вместо использования удобрений исследователи рассматривали возможность использования разных видов бактерий и отдельного покрытия семян пробиотики которые способствуют росту азотфиксирующие бактерии. [51] Обзор Азот — один из важнейших лимитирующих факторов жизни на Земле Генеральным спонсором конкурса, согласно нашему краудфандингу, стал предприниматель Константин Синюшин, за что ему огромный человеческий респект! Что вообще такое азот? На Земле азот представлен своим двухатомным соединением в виде газа (N2) и составляет добрых 76% объема нашей атмосферы. Но, как это ни странно, живым организмам азот достается не так уж и просто. В чем же проблема? Она заключается в тройной ковалентной связи между двумя атомами азота, благодаря которой в нормальных земных условиях азот, можно сказать, инертный газ. Но давайте разбираться, как, откуда и зачем этот азот нам сдался. Люди получают азот вместе с пищей, где он содержится в аминокислотах. Однако человек и большинство животных не могут сами синтезировать все необходимые аминокислоты. Часть из них (называемая незаменимыми аминокислотами) должна поступать вместе с пищей. И только автотрофным организмам (таким как растения и многие микроорганизмы) дано синтезировать органику из минеральных веществ. Но и им для этого требуется исходный материал. Но цивилизация не стоит на месте, да и удобрения тоже. Так вот. Азотистые соединения. В природе азот фиксируется био- и абиогенным путями (табл. 1). Биогенный способ фиксации азота осуществляется с помощью бактерий; абиогенный — во время грозы, когда на месте удара молнии температура может достигать 25 000 °С, в результате чего образуются оксиды азота. Далее они взаимодействуют с водой и образуют азотистую/азотную кислоту, которая потом прореагирует с солью и образует нитрат или же нитрит какого-либо металла. Но в современном мире этих двух пассивных способов не хватает. Как уже было сказано выше, азот — лимитирующий элемент всей жизни на Земле. Ежегодно около 2% всей вырабатываемой человечеством электроэнергии тратится на процесс Габера-Боша для создания азотных удобрений [1]! 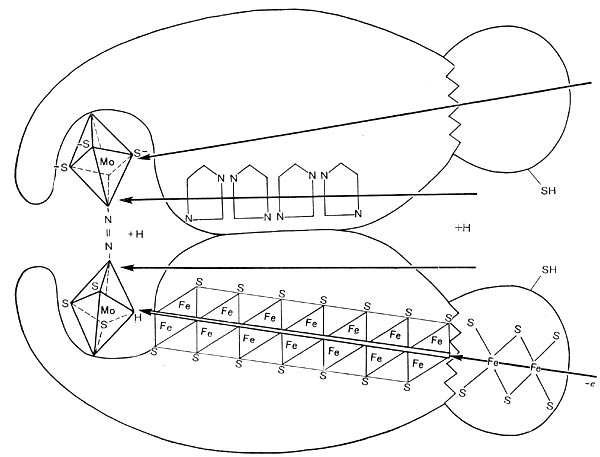
Рис. 179. Схема действия нитрогеназы.
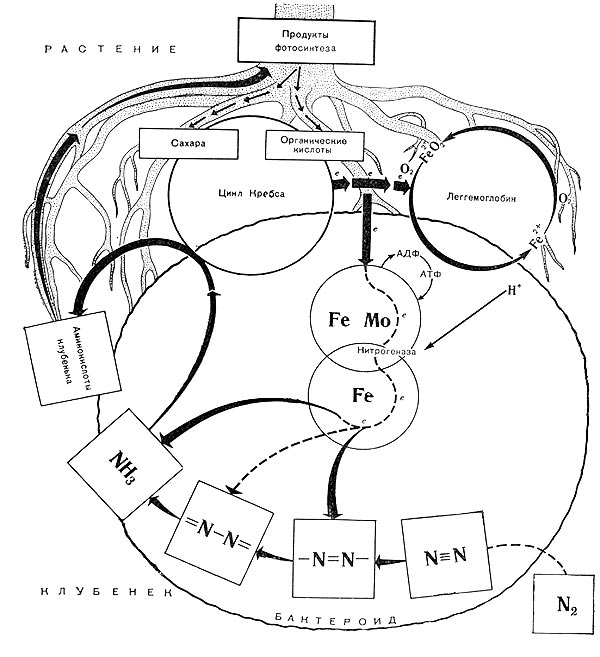
Рис. 80. Схема симбиотической азотфиксации
Рис. 179. Схема действия нитрогеназы.
Рис. 80. Схема симбиотической азотфиксации
3 ) или родственные азотистые соединения в почве. [1] Атмосферный азот молекулярный диазот, относительно нереактивная молекула, которая метаболически бесполезна для всех, кроме нескольких микроорганизмов. Преобразователи биологической азотфиксации N
2 в аммиак, который метаболизируется большинством организмов.Содержание
Фиксация
Небиологический

2 ) и кислородный газ ( О
2 ) присутствует в атмосфере в НЕТ
Икс (оксиды азота). НЕТ
Икс может реагировать с водой с образованием азотистая кислота или же азотная кислота, который просачивается в почву, где делает нитрат, что полезно для растений. Азот в атмосфере очень стабилен и инертен из-за тройная связь между атомами в N
2 молекула. [7] Молния производит достаточно энергии и тепла, чтобы разорвать эту связь [7] позволяя атомам азота реагировать с кислородом, образуя НЕТ
Икс . Эти соединения не могут быть использованы растениями, но когда эта молекула охлаждается, она реагирует с кислородом с образованием НЕТ
2 . [8] Эта молекула, в свою очередь, реагирует с водой с образованием HNO
3 (азотная кислота) или его ион НЕТ −
3 (нитрат), которую могут использовать растения. [9] [7] Биологические
2 . [12] Преобразование N
2 в аммиак происходит при металлический кластер называется FeMoco, сокращение от iron-молибден кофактор. Механизм действует через серию протонирование и стадии восстановления, на которых FeMoco активный сайт гидрогенизирует то N
2 субстрат. [13] В свободной жизни диазотрофыаммиак, образующийся нитрогеназой, ассимилируется в глутамат сквозь глютамин синтетаза/ путь глутаматсинтазы. Микробный гены nif необходимые для азотфиксации, широко распространены в различных средах. [14] [15] Микроорганизмы
Симбиозы корневых клубеньков
Семейство бобовых
Не зернобобовые

Эндосимбиоз у диатомовых водорослей
2 -фиксирующий эндосимбионт органеллы. Сфероидные тела находятся в цитоплазме диатомовых водорослей и неотделимы от своих хозяев. [26] [27] Инженерия эукариотической нитрогеназы
Промышленные процессы
История
Франк-Каро процесс
Процесс Габера

3 )
5 ( N
2 ) 2+ . [34] Снижение содержания азота в окружающей среде
ионный остаток затруднен.
4 был получен после гидролиза.… " [37] 
2) (dppe)2 (M = Mo, W), который протонирует с образованием промежуточных продуктов с лиганд M = N− N
2 . В 1995 г. молибден(III) был открыт амидокомплекс, расщепляющий N
2 дать соответствующие молибден (VI) нитрид. [39] Этот и родственные ему терминальные нитридокомплексы были использованы для получения нитрилы. [40]
2 , хотя и с небольшими оборотами. [38] [41] [42] [43] В этих системах, как и в биологической, водород подводится к субстрату гетеролитически, с помощью протонов и сильного Восстановитель а не с ЧАС
2 .
Фотохимическое и электрохимическое восстановление азота
3 может производиться непосредственно из азота и воды при температуре и давлении окружающей среды. [ нужна цитата ]Исследование

Автор
Редакторы

Сценарий азота в жизни
Таблица 1. Количество азота, фиксируемого различными путями
Тип фиксации N2 фиксировано,
млн. тонн в годАбиотический Азотистые удобрения 50 Окисление в почве 20 Грозы 10 ВСЕГО 80 Биотический Агрокультуры 90 Леса и дикие растения 50 Море 35 ВСЕГО 175 Итого в мире 255
| Свободноживущие | Симбионты растений | ||
|---|---|---|---|
| Аэробы | Анаэробы | Бобовые | Другие растения |
| Азотобактер Клебсиеллы(нек.) Цианобактерии | Клостридии Пурпурные серобактерии Пурпурные несерные бактерии Зелёные серобактерии | Клубеньковые бактерии | Франкии Азоспириллы |
Нитрогеназа, или как это сделано
Фермент, отвечающий за фиксацию азота, называется нитрогеназой и представляет собой комплекс из двух железосодержащих белков: Mo-Fe-белка и Fe-белка. Так как железо в них находится в виде ионов Fe 2+ и Fe 3+ , нитрогеназа крайне чувствительна к кислороду и после взаимодействия с ним навсегда утрачивает свои полезные свойства.
Для биологической фиксации азота клетке необходимы:
- активная нитрогеназа;
- Mg 2+ и постоянное снабжение АТФ;
- сильный восстановитель, т.е. подходящий источник электронов;
- низкое содержание кислорода.
Общая формула восстановления азота до аммиака выглядит следующим образом:
N2 + 8H + + 8e − + 16Mg·АТФ = 2NH3 + H2 + 16Mg·АДФ + 16Фн
Формула формулой, но давайте разбираться, как все происходит в жизни:

- Атмосферный азот попадает в клетку прокариотического организма.
- При условии недостатка аммиака и нитратов в окружающей среде клетка начинает процессы транскрипции и трансляции, в результате которых синтезируется фермент нитрогеназа.
- Азот — субстрат нитрогеназы — обеспечивается двумя электронами, протонами и 4-мя молекулами АТФ, связанными с магнием. Далее с ним происходят следующие превращения:
Хотя механизм фиксации азота нитрогеназой изучен пока недостаточно, есть основания полагать, что белок, содержащий молибден (Mo-Fe-белок), реагирует с потенциальными субстратами нитрогеназы, т.е. N2 и C2H2 (нитрогеназа без разбору восстанавливает все соединения с тройной связью), путем взаимодействия восстанавливаемого субстрата с атомами молибдена белка. Fe-бeлок с меньшей молекулярной массой связывает АТФ в виде мономагниевой соли и становится более мощным восстановителем, принимая электроны от физиологической электрон-донорной системы (рис. 1). Находясь в таком восстановленном состоянии, Fе-белок доводит до наиболее восстановленного состояния Mo-Fe-белок. Схематически процесс фиксации включает восстановление атомов Fe в железосодержащем белке с меньшей молекулярной массой, вследствие чего АТФ гидролизуется с образованием АДФ и фосфата (Фн). Электроны атомов железа в Mo-Fe-белке используется затем для восстановления связанного с азотом субстрата. Должно произойти несколько таких переносов электронов, прежде чем фермент высвободит конечный восстановленный продукт (NН3, если субстратом служит N2). При каждом переносе электрона от Fe-белка на Mo-Fe-бeлок АТФ гидролизуется до АДФ [2], [3]. Общая схема этого процесса восстановления показана на рисунке 1.

Рисунок 1. Донором электрона в реакции служит восстановленный ферредоксин. Электрон с затратой энергии АТФ переносится Fe-белком с донора на Mo-Fe-белок, который, в свою очередь, трижды отдает пару электронов двум атомам азота, восстанавливая связанную в активном центре молекулу N2 до двух NH3 через следующие стадии: азот → диамин → гидразин → аммиак.
Как известно, вечных двигателей не бывает. Так же и с нитрогеназой. При условиях, оптимальных для фиксации азота, только 75% доступных электронов действительно используются для этой цели. Остальные 25% расходуются на восстановление протонов до водорода в АТФ-зависимом процессе, катализируемом нитрогеназой, который, как и процесс восстановления азота, нуждается в Mg 2+ и восстановителе. Многие азотфиксирующие бактерии не могут избежать этой продуцирующей водород побочной реакции, которая и является причиной низкой эффективности фиксации азота этими клетками, ведь на восстановление протонов АТФ тратится впустую, а это до трети энергии, протекающей через нитрогеназу.
Как видно, нитрогеназа — очень невыгодный белок. Но нам известны целые семейства растений, которые принимают азотфиксирующих бактерий в качестве симбионтов. Давайте разберемся, как им удается делать это.
Проникновенные отношения
Заселение клеток корня растения-хозяина происходит после того, как корень натолкнется на симбиотических бактерий, оставшихся в почве с прошлого вегетационного периода. Бактерии подползают к корням, после чего разрушают клеточные стенки растения с помощью фермента пектиназы и размещаются в вакуолях (рис. 2). В результате растительная клетка оказывается переполненной бактериями, которые затем прекращают делиться, увеличиваются в размерах почти в 40 раз и одновременно изменяют свое строение и функцию, превращаясь в бактероиды, богатые нитрогеназой. Одновременно с вторжением бактерий в клетки растения-хозяина начинается активное деление инфицированной клетки и нескольких слоев соседних неинфицированных клеток. Это способствует распространению бактерий и приводит к образованию корневых клубеньков, столь характерных для этой симбиотической ассоциации.

Рисунок 2. Вторжение бактерий в клетки растения-хозяина. а — Бактерии проникли внутрь клеток корня растения. б — Образовавшиеся в корневых клетках бактероиды. в — Деление клеток, инфицированных бактериями-симбионтами. г — Рост клубенька.

Рисунок 3. Схематичное изображение симбиотических связей клубенькового растения.
Чтобы симбиотическая система могла фиксировать азот, она должна удовлетворять трем обязательным требованиям:
- образование корневых клубеньков;
- дифференциация бактерий в бактероиды;
- образование леггемоглобина.
При выполнении этих условий фиксация азота будет продолжаться в течение всего жизненного цикла растения, обычно до формирования семян, когда клубеньки стареют, и фиксация азота прекращается. Некоторые бактероиды утрачивают способность к дальнейшему размножению, однако остальные сохраняют жизнеспособность, и после старения и отмирания растения-хозяина живут в почве до начала нового вегетационного периода [3].
Кислородный искуситель
Фермент нитрогеназа необратимо разрушается при контакте с кислородом, и поэтому микроорганизмы, фиксирующие азот на планете, атмосфера которой содержит аж 20% кислорода, нашли различные способы исключить его из зоны фиксации азота. Такая адаптация не понадобилась лишь облигатным анаэробам, которые решили эту проблему радикальным способом.
Клубеньковые вывернулись с помощью леггемоглобина, который обладает очень высоким сродством к кислороду. Бактерии-симбионты получают этот элемент в концентрации, безопасной для их нитрогеназы: кислород не накапливается в больших количествах, однако его достаточно для обеспечения дыхательного метаболизма бактерий.
- Механизм повышенного уровня дыхания. Азотобактеры удаляют из себя кислород посредством окисления органических веществ. Энергия при этом не запасается.
- Механизм локализации азотфиксации в гетероцистах. Характерен для цианобактерий, способных к фотосинтезу с выделением кислорода. Для защиты нитрогеназы от кислорода они имеют особые, лишенные хлорофилла клетки — гетероцисты (о них будет сказано в последней главе).
- Механизм симбиотической защиты. Характерен для клубеньковых бактерий. В корнях бобовых продуцируется леггемоглобин, выполняющий функцию защиты от избытка кислорода [4].
Синтез жизни
Хорошо, растение добыло себе аммиак, а что дальше? Дальше он включается в реакции синтеза аминокислот. Так из аспарагиновой кислоты синтезируется аспарагин, из глутамата — глутамин (рис. 4). В виде глутамина азот доставляется ко всем клеткам организма, где используется для синтеза азотсодержащих органических соединений [5].

Рисунок 4. Глутамин. а — Реакция превращения глутамата в глутамин. б — Функции глутамина.
Аспарагин также синтезируется с участием аммиака (рис. 5) [6].

Побочные ветви цикла азота

Рисунок 6. Цикл азота в природе.
Нитрификация — это процесс окисления аммиака до азотистой кислоты, а далее ее окисление до азотной. Сначала аммиак присваивается нитрозными бактериями, которые окисляют аммиак в три стадии:
Далее образованную азотистую кислоту перехватывают нитратные бактерии, которые доокисляют ее до азотной кислоты в ходе одной реакции:
Двумя этими процессами в почве (или водоеме) заняты различные бактерии. Но все они — хемолитоавтотрофы, то есть получают энергию при окислении аммиака и используют ее для окисления углекислого газа [7].
Только после того, как почвенные бактерии осуществили перечисленные выше процессы, в цикл могут включиться бактерии-денитрификаторы. В их цитоплазмах проходят следующие превращения:
Микробиологическая денитрификация в почве вызывает потерю зафиксированного в солях азота. Это очень распространенное в природе явление, в ходе которого каждый год в атмосферу Земли возвращается 270–330 млн. тонн азота. Поэтому часто денитрификацию рассматривают как невыгодный человечеству процесс и пытаются его избежать. Сами денитрификаторы получают почти 70% энергии, которую они получили бы при проведении реакций аэробного метаболизма. Отлично устроились.
Но, согласитесь, должна же быть у нитрифицирующих бактерий хоть какая-нибудь конкуренция. И, конечно, природа не преминула ее создать. Такие бактерии идут в обход нитрификации и денитрификации. В их цитоплазмах протекает гениально примитивная реакция:
Эта примитивная, но действенная реакция получила название анаммокс (рис. 7). В масштабах планеты этот процесс дает начало 30–50% азота, образующегося в океанах. Анаммокс представляет большой интерес для биотехнологов, так как промежуточным продуктом этой реакции является гидразин — ядовитое вещество, применяемое как топливо для ракет. Также бактерий, осуществляющих аноммокс, используют для освобождения водоемов от излишков аммиака [8].

Осталось разобрать последнюю ветвь цикла азота. Это гниение. Автотроф, потребляющий нитраты, рано или поздно погибнет. Тут-то в игру включатся различные бактерии-аммонификаторы. Они будут заниматься разложением сначала белков, а затем аминокислот.
Первая стадия разложения белков — гидролиз. Он осуществляется как микробными, так и собственными ферментами клеток погибшего организма, высвобождаемыми из лизосом в результате смерти клеток (аутолиз). Белки распадаются на аминокислоты, которые, в свою очередь, дезаминируются, в результате образуя аммоний и углекислый газ.
Вот так, например, происходит анаэробная аммонификация у некоторых представителей бактериального семейства клостридий:
На этом мы завершили рассмотрение цикла азота, и можно смело перейти к рассмотрению организмов, азот фиксирующих [9].
Бактериальные собственники

Рисунок 8. Гетероциста. а — гетероциста в составе нитчатой водоросли; б — схематическое изображение процессов, протекающих в гетероцисте. В гетероцисте молекулярный азот с помощью нитрогеназы переводится в аммиак, затем при участии глутамин-синтазы аммоний с глутаматом превращаются в глутамин. Поскольку глутамат-синтаза (ГОГАТ) находится, в основном, в вегетативных клетках, образованный глутамин передается туда из гетероцист и посредством ГОГАТ превращается в глутамат. Глутамат поступает из вегетативной клетки обратно в гетероцисту, и цикл замыкается. Из глутамата и глутамина образуются остальные аминокислоты в вегетативных клетках. Связанный азот запасается в гранулах в виде аргинина и аспартата. Усваиваемый атмосферный углерод при фотосинтезе в вегетативных клетках переходит в органическую форму — глюкозу, которая может метаболизироваться до пирувата, а тот в изоцитрат в цикле Кребса. Углевод (глюкоза/сахароза), поступая в гетероцисту, окисляется в пентозофосфатном цикле до углекислоты. При этом образуются протоны и электроны, необходимые для синтеза аммония из молекулярного азота. В гетероцистах обнаружена инвертаза, разлагающая сахарозу на глюкозу и фруктозу. Изоцитрат с помощью изоцитратдегидрогеназы преобразуется в α-кетоглутарат.
Итак, в этой статье хоть и поверхностно, но охвачены все аспекты цикла азота, которые известны науке на сегодняшний день. Может быть, вскоре мы научимся использовать процессы, протекающие в бактериях, для чистой добычи азотных соединений, ведь у них огромный для этого потенциал. Надеюсь, что эта статья заинтересовала вас. Спасибо за прочтение!
Читайте также: