
Биоэлектрические явления в клетке кратко
Обновлено: 05.07.2024
Нейроны, мышечная и железистая клетки относятся к возбудимым тканям и обладают следующими общими свойствами: раздражимостью, возбудимостью, проводимостью и лабильностью.
Раздражимость -способность отвечать на действие раздражающих факторов изменением структурных и функциональных свойств клетки.
Раздражимостью обладают все ткани животных и растительных организмов. Раздражителями являются физические, химические или энергетические факторы внутренней среды организма или действующие на организм из внешней среды. После действия раздражителя изменяются свойства мембраны (электрический потенциал, проницаемость, активность переносчиков, свойства ионных каналов), метаболизм и другие внутриклеточные процессы. По силе раздражители подразделяются на пороговые. под- и надпороговые. Пороговые - минимальные по силе, способные вызвать ответную реакцию. Раздражение клеток соединительной ткани может сопровождаться трансформацией, пролиферацией, размножением, хемотаксисом и фагоцитозом.
Возбудимость – способностью возбудимой ткани осуществить специфический ответ на действие раздражителя. Заключается в изменении уровня потенциала мембраны (наиболее часто деполяризация и генерация потенциала действия) и в специфических функциональных проявлениях, свойственных данной ткани - сокращение мышцы, проведение возбуждения по нерву, выделение секрета железистой клеткой. Возбудимость оценивается порогом - минимальным по силе раздражителем, вызывающим видимую ответную реакцию. Более сильные по величине раздражители - надпороговые, более слабые - подпороговые.
Проводимость - способность локальное изменение свойств мембраны, возникшее в области действия раздражителя, распространить по протяжённости мембраны, вплоть до охвата возбуждением всей мембраны клетки.
Лабильность - способность ткани ответить на определенное количество стимулов в единицу времени. Является мерой функционального диапазона ткани, мерой функциональной подвижности, позволяет количественно измерить и сравнить функциональные возможности тканей и их изменение при каких-то воздействиях. Например, лабильность нейрона выше лабильности мышцы, лабильность утомлённой мышцы ниже ее лабильности до выполнения работы.
Рис. 1. Структура биологической мембраны.
Биоэлектрические явления в тканях.
Исследования электрических явлений в биологических системах начаты итальянским физиком Гальвани в 18 веке, который на нервно-мышечном препарате лапки лягушки продемонстрировал факт существования "животного" электричества. Основополагающие данные получены Ходжкиным, Хаксли и Катцем в 40-50 годы текущего столетия благодаря применению внутриклеточного микроэлектрода.
Рис. 2. Ионный канал.
Структурами клетки, которые обеспечивают электрогенез являются: мембрана клетки, изолирующая её от электрических потенциалов соседних клеток; ионные каналы. Абсолютная величина поляризации определяется неравенством в концентрации ионов в клетке и во внеклеточном пространстве; сохранение постоянного потенциала мембраны и его восстановление обеспечивается работой электрогенного натрий/калиевого насоса с непрерывным расходованием АТФ.
Ионные каналы - особые образования в мембране клетки, представляющие собой олигомерные (состоящие из нескольких субъединиц) белки. Центральным образованием канала является молекула белка, которая пронизывает мембрану таким образом, что в ее гидрофильном центре формируется канал-пора, через которую в клетку способны проникать соединения, диаметр которых не превышает диаметра поры (обычно это ионы). В ионном канале выделяют несколько участков:
1) активационные и инактивационные ворота - особые участки белка, которые, изменяя свою конфигурацию, переводят канал из открытого состояния в закрытое;
2) ионный фильтр - место связывания с ионами, которые пропускает данный канал, при этом канал характеризуется селективностью (способность пропускать только один вид ионов);
3) рецепторы - участки белка, которыми канал связывается с различными регуляторными молекулами;
4) участок модификации - особая часть белка, которая чаще всего подвергается реакции фосфорилирования-дефосфорилирования, что изменяет пропускную способность канала.
Вокруг главной субъединицы канала располагается система из нескольких субъединиц, которые формируют участки для взаимодействия с мембранными регуляторными белками, различными медиаторами, а также фармакологически активными веществами.
Классификация ионных каналов по их функциям:
1) по количеству ионов, для которых канал проницаем, каналы делят на селективные (проницаемы только для одного вида ионов) и неселективные (проницаемы для нескольких видов ионов);
2) по характеру ионов, которые они пропускают на Na + , Ca ++ , Cl - , K + -каналы;
3) по способу регуляции делятся на потенциалзависимые и потенциалнезависимые. Потенциалзависимые каналы реагируют на изменение потенциала мембраны клетки, и при достижении потенциалом определенной величины, канал переходит в активное состояние, начиная пропускать ионы по их градиенту концентрации. Так, натриевые и быстрые кальциевые каналы являются потенциалзависимыми, их активация происходит при снижении мембранного потенциала до 50-60 мВ, при этом ток ионов Na + и Ca ++ в клетку вызывает падение потенциала и генерацию ПД. Калиевые потенциалзависимые каналы активируются при развитии ПД и, обеспечивая ток ионов К + из клетки, вызывают реполяризацию мембраны.
Потенциалнезависимые каналы (хемоуправляемые) реагируют не на изменение мембранного потенциала, а на взаимодействие рецепторов, с которыми они взаимосвязаны, и их лигандов. Так, Cl - -каналы связаны с ГАМК-рецепторами и при взаимодействии этих рецепторов с g-аминомасляной кислотой они активируются и обеспечивают ток ионов хлора в клетку, вызывая ее гиперполяризацию и снижение возбудимости.
1.3. Потенциал покоя и действия. Мембранно-ионная теория происхождения потенциала покоя и действия. Местное и распространяющееся возбуждение.
Мембрана любой живой клетки поляризована, внутренняя поверхность элетроотрицательна по отношению к наружной. Мембранный потенциал равен - (минус) 70 - (90) мв. При возбуждении происходит снижение величины исходного потенциала покоя с перезарядкой мембраны. Формирование и сохранение потенциала покоя обусловлено непрерывным движением ионов по ионным каналам мембраны, постоянно существующей разностью концентраций катионов по обе стороны мембраны, непрерывной работой натрий-калиевого насоса. За счет постоянного удаления из клетки иона натрия и активного переноса в клетку иона калия сохраняется разность концентраций ионов и поляризация мембраны. Концентрация иона калия в клетке превышает внеклеточную концентрацию в 30 - 40 раз, внеклеточная концентрация натрия примерно на порядок выше внутриклеточной. Электроотрицательность внутренней поверхности мембраны обусловлена наличием в клетке избытка анионов органических соединений, абсолютная величина потенциала покоя (мембранный потенциал, трансмембранный потенциал, равновесный калиевый потенциал) обусловлена главным образом соотношением внутри- и внеклеточной концентраций ионов калия и удовлетворительно описывается уравнением Нернста:

(1)
Современная теория учитывает так же:
1) разницу концентраций ионов натрия, хлора, кальция;
2) проницаемость (Р) мембраны для каждого иона в текущий момент времени.
Наличие потенциала покоя позволяет клетке практически мгновенно после действия раздражителя перейти из состояния функционального покоя в состояние возбуждения.
Рис 3. Потенциал действия.
Возникновение потенциала действия (деполяризация)
Потенциал действия (ПД) развивается при наличии исходной поляризации мембраны (потенциала покоя) благодаря изменению проницаемости ионных каналов (натриевых и калиевых). После действия раздражителя потенциал покоя уменьшается, активация каналов повышает их проницаемость для ионов натрия, который входит в клетку и обеспечивает процесс деполяризации. Поступление в клетку иона натрия уменьшает электроотрицательность внутренней поверхности мембраны, что способствует активации новых ионных натриевых каналов и дальнейшему поступлению в клетку иона натрия. Действуют силы:
а) электростатическое притяжение внутриклеточных анионных группировок;
б) концентрационный градиент ионов натрия, направленный внутрь клетки.
Пик потенциала действия обусловлен равновесием поступления в клетку ионов натрия и равным их удалением под влиянием сил отталкивания одноимённо заряженных ионов.
Реполяризация
После инактивации (закрытия) натриевых каналов поступление в клетку ионов натрия становится минимальным. Выход из клетки ионов калия восстанавливает электроотрицательность внутренней поверхности мембраны. В последующем натрий/калиевый насос мембраны удаляет из клетки поступивший при деполяризации натрий и восстанавливает исходную концентрацию калия, который вышел из клетки при реполяризации.
Образцы сочинений-рассуждений по русскому языку: Я думаю, что счастье – это чувство и состояние полного.
Каждая живая клетка подобно гальваническому элементу является генератором электрического тока. Вся ее жизнедеятельность связана с электрическими явлениями, которые в настоящее время благодаря усовершенствованию электрофизиологических методов исследования поддаются довольно точному изучению. Существенную помощь в исследовании ряда электрических явлений в живых клетках оказали микроэлектроды, представляющие собой изготовленные из тугоплавкого стекла толстостенные капилляры диаметром 1 мм с оттянутыми концами (диаметром от 0,5 до 0,1 мкм) и заполненные 2,5 М раствором КС1. Такие электроды можно вводить внутрь клетки, не причиняя ей никакого вреда.
Если два таких микроэлектрода, соединенные с чувствительным измерительным прибором, приложить к любым двум точкам наружной поверхности клеточной мембраны, то стрелка прибора не отклонится: это значит, что между разными точками наружной поверхности клеточной мембраны никакой разности потенциалов не существует. Но если один из микроэлектродов ввести внутрь клетки, стрелка отклонится: между наружной и внутренней поверхностями клеточной мембраны разность потенциалов существует (ее принято называть потенциалом покоя). Легко убедиться, что эта разность появляется в тот момент, когда микроэлектрод проходит через поверхность клетки и касается протоплазмы. При дальнейшем продвижении электрода через протоплазму разность потенциалов не изменяется; она исчезает только после того как электрод выходит из клетки через противоположную поверхность.
У разных клеток разный потенциал покоя. Он обычно колеблется в пределах от 50 до 100 милливольт (мВ). Наружная поверхность мембраны заряжена положительно, внутренняя — отрицательно. Мембранный потенциал существует только до тех пор, пока клетка жива. С ее гибелью исчезает и разность потенциалов между поверхностями мембраны.
Если на какой-либо участок мембраны нанести раздражение определенной силы, клетка из состояния покоя перейдет в деятельное состояние, причем заряды мембраны в месте раздражения изменяют свой знак. Подвергшийся раздражению участок наружной поверхности мембраны становится отрицательно заряженным по отношению к соответствующему участку внутренней поверхности (эта разность потенциалов получила название потенциала действия). Такое изменение знака зарядов при переходе в деятельное состояние определяется во всех клетках, которые способны под влиянием раздражения соответствующей интенсивности перейти в активное состояние — возбуждение. Изменение электрического состояния в месте раздражения длится недолго, в среднем около 1 миллисекунды (мсек), после чего быстро восстанавливается исходное состояние (потенциал покоя). Возникнув в одном месте, возбуждение быстро распространяется вдоль всей клетки, причем на каждом участке по ходу распространения возбуждения происходят описанные изменения электрических зарядов и последующее их восстановление.
Соответствующими измерениями показано, что потенциал действия имеет не только обратное направление, но и на 30—50 мВ больше потенциала покоя. Это значит, что при переходе клетки в активное состояние вначале происходит полная деполяризация мембраны, т. е. исчезновение потенциала покоя вследствие разряда поверхностей мембраны, и последующая ее перезарядка.
Таким образом, переход клетки из одного состояния в другое, как правило, сопровождается вполне определенными электрическими явлениями. Нельзя, например, вызвать возбуждение клетки без соответствующего изменения знаков заряда на поверхностях мембраны. И, наоборот, если с помощью внешнего источника тока соответствующим образом изменить знаки зарядов на клеточной мембране (положительный электрод ввести внутрь клетки, а отрицательный приложить к наружной ее поверхности) и вызвать хотя бы частичную ее деполяризацию (на 15—20 мВ),— в клетке возникнет возбуждение, которое начнет распространяться.
Что же лежит в основе наблюдаемых в возбудимых клетках электрических явлений? Общеизвестно, что почти все физические и химические процессы в природе сопровождаются определенными электрическими явлениями. В живых же клетках физические и химические процессы протекают непрерывно. Основную роль в возникновении электрических зарядов в клетке играют, по общепринятому мнению, ионные процессы (подобно тому как это наблюдается в гальванических элементах). Источником ионов в клетке служат соли. Как показали многочисленные исследования, клетки содержат 80% воды и 1,5% солей, главным образом калия и кальция. Одной из отличительных черт живой клетки является значительная разница в составе и концентрации ионов, в содержимом клетки и окружающей ее среде. Для большинства клеток характерна высокая внутренняя концентрация калия и низкая концентрация натрия. Содержание калия в клетке составляет 0,4%, в то время как в окружающей клетку среде калия всего содержится 0,13%. Натрия же в клетке 0,08%, между тем как в окружающей среде — 0,85%. Это своеобразие химического состава клетки и окружающей среды свойственно почти всем живым организмам. После же их гибели разница в концентрации ионов калия и натрия внутри и вне клетки выравнивается (по законам диффузии).
Военно-медицинское образование
Военно-медицинское образование — система обучения и воспитания различных категорий медицинского состава Вооруженных Сил (врачей,
Яхимов
Яхимов — бальнеологический курорт Чехии. Славится замечательными радиоактивными источниками, являющимися самыми сильными в бывшей
Янске лазне
Ценным продуктом детского питания являются куриные яйца. Аминокислотный состав белков яйца является близким к
Возбуждение представляет собой сложную совокупность физических, химических и физико-химических процессов, в результате которых происходит быстрое и кратковременное изменение электрического потенциала мембраны.
Позднее Дюбуа-Реймоном было установлено, что поврежденный участок мышцы имеет отрицательный заряд, а неповрежденный участок – положительный. При набрасывании нерва между поврежденным и неповрежденным участками мышцы возникает ток, который раздражает нерв и вызывает сокращение мышцы. Этот ток был назван током покоя, или током повреждения. Так было показано, что наружная поверхность мышечных клеток заряжена положительно по отношению к внутреннему содержимому.
Первая теория возникновения и поддержания мембранного потенциала была разработана Ю. Бернштейном (1902). Исходя из того, что мембрана клеток обладает высокой проницаемостью для ионов калия и малой проницаемостью для других ионов, он показал, что величину мембранного потенциала можно определить, используя формулу Нернста.
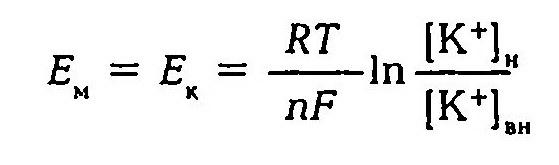
В 1949–1952 гг. А. Ходжкин, Э. Хаксли, Б. Катц создали со-временную мембранно-ионную теорию, согласно которой мембранный потенциал обусловлен не только концентрацией ионов калия, но и натрия и хлора, а также неодинаковой проницаемостью для этих ионов мембраны клетки. Цитоплазма нервных и мышечных клеток содержит в 30 -50 раз больше ионов калия, в 8–10 раз меньше ионов натрия и в 50 раз меньше ионов хлора, чем внеклеточная жидкость. Проницаемость мембраны для ионов обусловлена ионными каналами, макромолекулами белка, пронизывающими липидный слой. Одни каналы открыты постоянно, другие (потенциалозависимые) открываются и закрываются в ответ на изменения МП. Потенциалозависимые каналы подразделяются на натриевые, калиевые, кальциевые и хлорные. В состоянии физиологического покоя мембрана нервных клеток в 25 раз более проницаема для ионов калия, чем для ионов натрия.
Таким образом, согласно обновленной мембранной теории асимметричное распределение ионов по обе стороны мембраны и связанное с этим создание и поддержание мембранного потенциала обусловлено как избирательной проницаемостью мембраны для различных ионов, так и их концентрацией по обе стороны от мембраны, а более точно величину мембранного потенциала можно рассчитать по формуле.
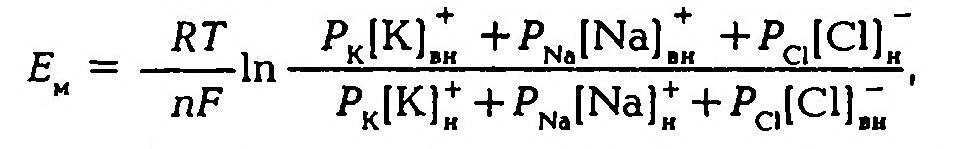
Поляризация мембраны в покое объясняется наличием открытых калиевых каналов и трансмембранным градиентом концентраций калия, что приводит к выходу части внутриклеточного калия в окружающую клетку среду, т. е. к появлению положительного заряда на наружной поверхности мембраны. Органические анионы – крупномолекулярные соединения, для которых мембрана клетки непроницаема, создают на внутренней поверхности мембраны отрицательный заряд. Поэтому чем больше разница концентраций калия по обе стороны от мембраны, тем больше его выходит и тем выше значения МП. Переход ионов калия и натрия через мембрану по их концентрационному градиенту в конечном итоге должен был бы привести к выравниванию концентрации этих ионов внутри клетки и в окружающей ее среде. Но в живых клетках этого не происходит, так как в клеточной мембране имеются натрий-калиевые насосы, которые обеспечивают выведение из клетки ионов натрия и введение в нее ионов калия, работая с затратой энергии. Они принимают и прямое участие в создании МП, так как за единицу времени ионов натрия выводится из клетки больше, чем вводится калия (в соотношении 3:2), что обеспечивает постоянный ток положительных ионов из клетки. То что выведение натрия зависит от наличия метаболической энергии, доказывается тем, что под действием динитрофенола, который блокирует метаболические процессы, выход натрия снижается примерно в 100 раз. Таким образом, возникновение и поддержание мембранного потенциала обусловлено избирательной проницаемостью мембраны клетки и работой натрий-калиевого насоса.
ЛЕКЦИЯ № 2. Физиологические свойства и особенности функционирования возбудимых тканей
ЛЕКЦИЯ № 2. Физиологические свойства и особенности функционирования возбудимых тканей 1. Физиологическая характеристика возбудимых тканей Основным свойством любой ткани является раздражимость, т. е. способность ткани изменять свои физиологические свойства и
1. Физиологическая характеристика возбудимых тканей
1. Физиологическая характеристика возбудимых тканей Основным свойством любой ткани является раздражимость, т. е. способность ткани изменять свои физиологические свойства и проявлять функциональные отправления в ответ на действие раздражителей.Раздражители – это
2. Законы раздражения возбудимых тканей
2. Законы раздражения возбудимых тканей Законы устанавливают зависимость ответной реакции ткани от параметров раздражителя. Эта зависимость характерна для высоко организованных тканей. Существуют три закона раздражения возбудимых тканей:1) закон силы
3. Понятие о состоянии покоя и активности возбудимых тканей
3. Понятие о состоянии покоя и активности возбудимых тканей О состоянии покоя в возбудимых тканях говорят в том случае, когда на ткань не действует раздражитель из внешней или внутренней среды. При этом наблюдается относительно постоянный уровень метаболизма, нет
2. Механизмы проведения возбуждения по нервному волокну. Законы проведения возбуждения по нервному волокну
2. Механизмы проведения возбуждения по нервному волокну. Законы проведения возбуждения по нервному волокну Механизм проведения возбуждения по нервным волокнам зависит от их типа. Существуют два типа нервных волокон: миелиновые и безмиелиновые.Процессы метаболизма в
2. Основные характеристики и законы возбудимых тканей
2. Основные характеристики и законы возбудимых тканей Основным свойством любой ткани является раздражимость, т. е. способность ткани изменять свои физиологические свойства и проявлять функциональные отправления в ответ на действие раздражителей.Раздражители – это
3. Понятие о состоянии покоя О и активности возбудимых тканей
3. Понятие о состоянии покоя О и активности возбудимых тканей О состоянии покоя в возбудимых тканях говорят в том случае, когда на ткань не действует раздражитель из внешней или внутренней среды. При этом наблюдается относительно постоянный уровень метаболизма.Основные
2 глава Патологические изменения в тканях
2 глава Патологические изменения в тканях При заболеваниях различных органов и систем в коже, соединительной ткани, миофасциальных структурах, надкостнице образуются патологические изменения , которые выявляются при наружном осмотре, поверхностной и глубокой
2 глава Патологические изменения в тканях
2 глава Патологические изменения в тканях При заболеваниях различных органов и систем в коже, соединительной ткани, миофасциальных структурах, надкостнице образуются патологические изменения, которые выявляются при наружном осмотре, поверхностной и глубокой
Глава 2. Физиология возбудимых тканей
Глава 2. Физиология возбудимых тканей Способность адаптироваться к постоянно изменяющимся условиям внешней среды является одним из основных признаков живых систем. В основе приспособительных реакций организма лежит раздражимость – способность реагировать на
Законы раздражения возбудимых тканей
Газообмен в легких и тканях
Газообмен в легких и тканях В легких происходит газообмен между поступающим в альвеолы воздухом и протекающей по капиллярам кровью. Интенсивному газообмену между воздухом альвеол и кровью способствует малая толщина описанного аэрогематического барьера. Альвеолярный
Инородное тело в тканях
Инородное тело в тканях Рыбья кость, иголки, осколки костей —
Абсцесс (скопление гноя в тканях)
Абсцесс (скопление гноя в тканях) • Взять по 20 г порошка клубней топинамбура, семян льна и травы донника желтого, по 10 г листьев мальвы, топинамбура и алтея; 2 ст. л. смеси залить 300 г кремниевой воды, варить 5 мин, настоять, укутав, 1 час, процедить, добавить 1 ст. л. меда.
Абсцесс, скопление гноя в тканях
Абсцесс, скопление гноя в тканях ? Взять по 20 г семян льна и травы донника, по 10 г листьев мальвы и алтея; 2 ст. л. смеси залить 300 мл воды, всыпать 1 ст. л. пищевой соды, варить 5 мин. Для компрессов.? Взять по 1 ст. л. луковиц лилии чисто-белой, корневищ купены душистой и корневищ
Абсцесс, скопление гноя в тканях
Абсцесс, скопление гноя в тканях — Взять по 20 г семян льна и травы донника лекарственного, по 10 г листьев мальвы и алтея; 2 ст. ложки смеси залить 300 г воды, варить 5 мин., охладить, процедить, добавить 3 ст. ложки яблочного уксуса. Для
В 1949-1952 гг. Ходжкин, Хаксли и Катц модифицировали существовавшие ранее теории и экспериментально обосновали основные положения мембранно-ионной теории .
Согласно этой теории возникновение МП (мембранного потенциала) обусловлено:
- Разной концентрацией ионов калия, натрия, кальция и хлора внутри клетки и во внеклеточной среде.
- Разной проницаемостью мембраны клеток для этих ионов в покое и при возбуждении.
Состояние покоя — это такое состояние, когда отсутствует раздражающее действие на организм. Это состояние относительное, т.к. абсолютный покой не совместим с жизнью.
Деятельное состояние — это изменение состояния организма, возникающее под действием факторов внешней среды.
Раздражимость — это способность организма отвечать на действие раздражающих факторов изменением его структурных и функциональных свойств.
Возбудимость
Возбудимость — способность ткани отвечать на раздражение специализированной реакцией — возбуждением. Возбудимость является одной из форм раздражимости.
Возбудимыми тканями называются такие ткани, клетки которых способны в ответ на раздражение генерировать специфическую реакцию — возбуждение .
Возбуждение — это специализированная ответная реакция живого объекта на действие раздражителя, проявляющаяся в изменениях его определенных параметров.
К возбудимым тканям относятся:
- нервная,
- мышечная,
- железистая.
Общие признаки возбуждения (присущи всем возбудимым тканям):
- Изменение уровня обменных процессов в тканях;
- Выделение различных видов энергии — тепловой, электрической.
Специфические признаки возбуждения (характерны для определенного вида тканей):
- Мышечная ткань — сокращение,
- Железистая — выделение секрета,
- Нервная — генерация и проведение нервного импульса.
Физиологические свойства возбудимых тканей
Основными свойствами всех возбудимых тканей являются:
- возбудимость,
- проводимость,
- лабильность.
Лабильность (функциональная подвижность) — это способность ткани реагировать на различные воздействия с определенной скоростью, то есть способность воспроизводить определенную частоту раздражения.
Мерой лабильности является наибольшее число ответных реакций, которыми возбудимая ткань способна реагировать за единицу времени в соответствии с частотой приложенных к ней раздражений.
Проводимость — это способность ткани и клетки передавать возбуждение.
Раздражение — это процесс воздействия на живую ткань агентов внешней по отношению к этой ткани среды.
Раздражитель — это причина, способная вызвать ответную реакцию.
Классификация раздражителей
- физические,
- химические,
- физико-химические,
- биологические.
По биологическому значению раздражители делятся на 2 группы :
- Адекватные — раздражители, к воздействию которых ткани в процессе эволюции приспособлены в наибольшей степени.
- Неадекватные — раздражители, к воздействию которых возбудимые ткани специально не приспособлены.
Состав плазматической мембраны
Липиды очень плотно упакованы в мембране, между ними нет практически никаких расстояний, поэтому мембрана плохо пропускает воду, практически непроницаема для ионов и других крупных молекул.
Белковые молекулы могут быть погружены в слой липидов с внеклеточной или с цитоплазматической стороны, либо могут целиком пронизывать мембрану.
Если белки прикреплены к поверхности мембраны, их называют периферическими. С внутренней стороны это будут белки-ферменты, а с наружной — белки-рецепторы.
Если белки пронизывают всю толщу мембраны клеток, то их называют интегральными или трансмембранными.
Такие белки образуют структуры, обеспечивающие движение ионов через мембрану.
Если белки образуют стенки поры, сквозь которую путем простой диффузии проходят ионы, то это ионные каналы.
Если трансмембранные белки перекачивают ионы против концентрационного и электрического градиентов, то это ионные насосы.
Все каналы, имеющиеся в живых тканях, можно разделить на 2 типа:
- первый тип — каналы покоя, которые спонтанно открываются и закрываются без всяких внешних воздействий;
- второй тип — gate-каналы (воротные каналы) — в покое они закрыты и открываются под действием раздражителей.
- неспецифические (каналы утечки, всегда открыты),
- специфические (селективные), обладающие способностью пропускать только определенные ионы при изменении заряда на мембране или действии химических веществ.
Транспорт веществ
Транспорт частиц через каналы является жизненно важным для клеток процессом.
Обычно транспорт веществ делят на пассивный (без затраты энергии), т.е. транспорт веществ по концентрационному, осмотическому и электрохимическому градиентам и активный (с затратой энергии).
Различают первично- и вторично-активный транспорт
Первично-активный транспорт ионов обеспечивается специальными ионными насосами, осуществляется с затратой энергии АТФ, против концентрационного градиента, т.е. перенос веществ происходит из меньшей концентрации через мембрану в большую концентрацию.
- Это вид транспорта для переноса веществ (глюкозы, аминокислот и др.) через мембрану также против градиента, но без затраты энергии .
- Эти вещества проходят через мембрану с помощью специальных переносчиков (напр. ионов Na), на транспорт которых и затрачивается энергия, а эти вещества движутся как бы попутно.
Мембранный потенциал или потенциал покоя
В состоянии покоя между наружной и внутренней поверхностью мембраны возникает разность зарядов или потенциалов, которая в дальнейшем была названа мембранным потенциалом (МП) или потенциалом покоя (ПП) .
Положительные заряды концентрируются на наружной поверхности мембраны, а отрицательные заряды — на внутренней поверхности.
Мембранный потенциал измеряется в отрицательных значениях, т.к. внутренняя поверхность мембраны заряжена отрицательно. Его величина колеблется от -60 до -90 мВ у разных клеток.
Методы измерения мембранного потенциала
В зависимости от места приложения электродов :
- внеклеточный с помощью макроэлектродов,
- внутриклеточный с помощью микроэлектродов.
1. Исследование ПП с помощью макроэлектродов осуществляют, прикладывая один из них к неповрежденному , а другой — к поврежденному участку ткани.
2. Микроэлетродный метод
Микроэлектрод представляет собой микропипетку диаметром 0,5 — 1 мкм, наполненную концентрированным солевым раствором (KCl). Внутри него может также находится неполяризующийся электрод — из серебра, золота или платины.
Второй электрод помещается во внеклеточную жидкость.
Оба электрода присоединяются к усилителю и осциллографу для регистрации потенциала. В момент прокалывания мембраны, осциллограф регистрирует появление отрицательного потенциала , соответствующего ПП.
1-ая причина — ионная асимметрия:
Таким образом, для этих ионов направление концентрационного градиента различно!
- Для K из клетки (из большей концентрации к меньшей);
- Для Na, Ca и Cl в клетку.
2-ая причина, обуславливающая поляризацию мембраны:
- различная проницаемость мембраны для различных ионов. В состоянии покоя мембрана в 25 раз более проницаема для ионов K, чем для Na, т.к. количество калиевых каналов на единицу площади мембраны намного больше, чем натриевых.
- Поскольку концентрация ионов K в цитоплазме намного выше, чем снаружи клетки, они начинают двигаться через канал и выходят из клетки.
- Ионы K несут положительные заряды, поэтому мембрана снаружи заряжается положительно.
Отрицательный заряд на внутренней поверхности мембраны обусловлен наличием органических анионов — крупномолекулярных соединений, которые заряжены отрицательно, и для которых мембрана непроницаема (глютамат, аспартат, органические фосфаты, сульфаты и др.)
Таким образов, на мембране формируется калиево-равновесный потенциал, т.к. уравновешиваются силы диффузии (выход K из клетки по градиенту концентрации) и электростатического взаимодействия (отталкивание выходящих ионов K положительным зарядом на наружной поверхности мембраны).
Калий является основным ионом, обеспечивающим формирование МП (ПП), что подтверждается формулой Нернста . По ней можно, зная концентрацию онов калия внутри и снаружи клетки, подсчитать величину ПП.

Натрий-калиевый насос или помпа
Для непрерывного активного переноса ионов Na и ионов K через мембрану клетки против концентрационного градиента существует Na-Л обменный насос. При каждом цикле он переносит 3 иона Na наружу и 2 иона K внутрь клетки.
Таким образом, МП представляет собой разность потенциалов между наружной и внутренней поверхностью мембраны клетки. Он является результатом разделения зарядов относительно клеточной мембраны , которое возникает за счет движения заряженных ионов (натрия и, преимущественно, калия) по концентрационным градиентам через ионные каналы, открытые в покое и за счет работы Na-K насоса .
Читайте также: