
Базальное тельце это кратко
Обновлено: 02.07.2024
БАЗАЛЬНОЕ ТЕЛЬЦЕ, внутриклеточное образование, расположенное у основания каждого жгутика или реснички у одноклеточных, а также в клетках тканей многоклеточных организмов и в сперматозоидах. Обычно цилиндрические, дл. ок. 0,5 мкм, диам. 0,1-0,2 мкм. По-видимому, Б. т. центриолярного происхождения (например, в сперматидах удаётся проследить преобразование центриоли в Б. т., а у ряда жгутиковых одно и тоже Б. т. может нести жгутик и, подобно центриоли, принимать участие в образовании веретена деления клетки). Иногда Б. т. наз. блефаропластом.
Смотреть что такое БАЗАЛЬНОЕ ТЕЛЬЦЕ в других словарях:
БАЗАЛЬНОЕ ТЕЛЬЦЕ
внутриклеточное образование, расположенное у основания каждого жгутика или реснички у одноклеточных, а также в клетках тканей многоклеточных ор. смотреть
БАЗАЛЬНОЕ ТЕЛЬЦЕ
БАЗАЛЬНОЕ ТЕЛЬЦЕ кинетосомa (corpusculum basale), внутриклеточная структура эукариот, лежащая в основании ресничек и жгутиков и служащая для них опоро. смотреть
БАЗАЛЬНОЕ ТЕЛЬЦЕ
kinetosome - кинетосома, базальное тельце.Форма центриоли , способна к самовоспроизводству; К. располагаются в основании жгутиков и ресничек. смотреть
БАЗАЛЬНОЕ ТЕЛЬЦЕ
Базальное тельцеорганоид, от к-рого исходят жгутики бактерий и простейших. Располагается в цитоплазме. Имеет вид дисков, состоящих из проксимальных кон. смотреть
БАЗАЛЬНОЕ ТЕЛЬЦЕ
(corpusculum basale, LNH; син. гранула базальная)органелла в виде мелкого тельца в основании ресничек или жгутиков клетки; по ультраструктуре, отношени. смотреть
БАЗАЛЬНОЕ ТЕЛЬЦЕ
базальное тельце (corpusculum basale, LNH; син. гранула базальная) — органелла в виде мелкого тельца в основании ресничек или жгутиков клетки; по ультраструктуре, отношению к красителям, способам репродукции и функциям является гомологом центриоли.
БАЗАЛЬНОЕ ТЕЛЬЦЕ
(corpusculum basale, LNH; син. гранула базальная) органелла в виде мелкого тельца в основании ресничек или жгутиков клетки; по ультраструктуре, отношению к красителям, способам репродукции и функциям является гомологом центриоли. смотреть
БАЗАЛЬНОЕ ТЕЛЬЦЕ
БАЗАЛЬНОЕ ТЕЛЬЦЕ, corpusculum basale (лат. corpusculum, i, n тельце и basalis f) —модифицированный центриоль, расположенный между основанием ресницы и ее корешком.
БАЗАЛЬНОЕ ТЕЛЬЦЕ
(реснички, кинетоцилии) basal corpuscles, corpusculum basale (cilii kinetocilii)
БАЗАЛЬНОЕ ТЕЛЬЦЕ
1) blépharoblaste, blépharoplaste 2) corpuscule basal 3) grain basal 4) granule basal 5) kinétosome 6) mastigosome
У многих клеток животных, вышедших из клеточного цикла, в G0-стадии центриоли принимают участие в образовании аппарата движения – ресничек. Их две группы: кинетоцилии, характерные для специальных эпителиев (ресничные эпителии трахеи, яйцеводов) или свободно плавающих клеток (сперматозоиды, простейшие), и так называемые первичные реснички, встречающиеся во многих клетках, не обладающих способностью к движению.
Клетки, имеющие реснички или жгутики, обладают способностью двигаться, будучи в свободном состоянии, или же перемещать жидкости в случае, если клетки неподвижны. Свободноживущие одноклеточные организмы, снабженные одним или несколькими жгутиками, обычно движутся тем концом вперед, который несет жгутики. У спермиев некоторых животных: жгутик, располагаясь сзади, толкает тело клетки вперед. Скорость движения клеток за счет работы жгутиков может достигать очень большой величины (до 5 мм / мин).
Множественные реснички обеспечивают движение свободноживущих клеток, таких как инфузории или некоторые жгутиконосцы. Реснички эпителиальных клеток многих беспозвоночных и позвоночных животных обеспечивают поток жидкостей вдоль поверхности таких клеток. Число ресничек на клетку может достигать 300 в эпителии трахеи; у инфузории туфельки на клетку приходится 10-14 тыс. рядами расположенных ресничек.
При движении ресничек и жгутиков не происходит уменьшения их длины, поэтому неправильно называть это движение сокращением. Траектория движения ресничек очень разнообразна (рис. 288). В различных клетках это движение может быть маятникообразным, крючкообразным, воронкообразным или волнообразным.
У многоресничных клеток (инфузории, клетки ресничного эпителия) движение ресничек не хаотично, а строго упорядочено. В этом случае реснички расположены рядами. В продольном ряду отдельные реснички начинают движение и проходят отдельные его фазы по очереди, метахронно. В поперечном же ряду все реснички находятся в одной фазе движения (синхронны). Это создает движущую волну по поверхности клетки (рис. )289.
Ресничка представляет собой тонкий цилиндрический вырост цитоплазмы с постоянным диаметром 300 нм. Этот вырост от основания до самой его верхушки покрыт плазматической мембраной. Внутри выроста расположена аксонема, сложная структура, состоящая в основном из микротрубочек. Нижняя, проксимальная часть реснички, базальное тельце, погружена в цитоплазму. Диаметры аксонемы и базального тельца одинаковы (около 200 нм).
На поперечном сечении реснички видна плазматическая мембрана, окружающая аксонему. Аксонема в своем составе имеет девять дублетов микротрубочек, образующих внешнюю стенку цилиндра аксонемы. Дублеты микротрубочек слегка повернуты (около 100) по отношению к радиусу аксонемы. В центре аксонемы располагается пара центральных микротрубочек. В целом систему микротрубочек реснички описывают как (9х2)+2. В дублетах микротрубочек также различают А-микротрубочку, состоящую из 13 субъединиц, и В-микротрубочку, неполную, содержащую 11 субъединиц. А-микротрубочка несет на себе ручки, которые направлены к В-микротрубочке соседнего дуплета. От А-микротрубочки к центру аксонемы отходит радиальная связка, или спица, оканчивающаяся головкой, присоединяющейся к центральной муфте, имеющей диаметр около 70 нм, окружающей две центральные микротрубочки. Последние лежат отдельно друг от друга на расстоянии около 25 нм. Таким образом, в аксонеме располагается 20 продольных микротрубочек, в то время как в базальном тельце их 27 (рис. 291, 292).
Базальное тельце состоит из 9 триплетов микротрубочек (как и центриоль), имеет ручки, втулку и спицы, расположенные в проксимальной (нижней) ее части. На участке базального тельца, примыкающем к плазматической мембране, есть девять придатков, выступов, идущих от каждого триплета микротрубочек к плазматической мембране и связывающих его с клеточной поверхностью.
Базальное тельце и аксонема структурно связаны друг с другом и составляют единое целое: А- и В-микротрубочки триплетов базального тельца продолжаются в А- и В-микротрубочках дуплетов аксонемы. Однако внутренние части аксонемы и базального тельца значительно отличны друг от друга. В зоне перехода базального тела в аксонему наблюдают аморфную поперечную пластинку, которая как бы отделяет эти две части. Центральные микротрубочки аксонемы начинаются от этой пластинки так же, как в этом месте начинается и центральная муфта (капсула) (рис. 290).

В основании ресничек и жгутиков часто встречаются исчерченные корешки, или кинетодесмы, представляющие собой пучки тонких (6 нм) фибрилл, обладающих поперечной исчерченностью (рис. 293). Кинетодесмы простираются от базальных телец вглубь цитоплазмы по направлению к ядру.
При движении ресничек не происходит изменения их длины, они не “сокращаются”, а изгибаются, бьются. Оказалось, что механически отделенные реснички способны к биению в присутствии АТФ. При отделении ресничек базальные тельца остаются в теле клетки. Для механической работы ресничек базальное тело не нужно, а только аксонема участвует в генерации движения. Движение ресничек обеспечивают “ручки”, сидящие на А-микротрубочках.
Установлено, что в состав ручек входят белки динеины. Это большие белковые компоненты, состоящие из 9-12 полипептидных цепей, содержащие 2-3 глобулярные головки, связанные в общий корешок гибкими хвостами (рис. 294). Каждая головка динеина обладает АТФ-азной активностью, которая возрастает примерно в 6 раз при ассоциации с микротрубочками. В состав каждой ручки входит один белковый комплекс, одна молекула динеина. Именно динеин ответственен за это движение, он является мотором или двигателем при биении ресничек. Механизм этого движения следующий.
За счет смещения пар микротрубочек одна относительно другой (рис. 295), аналогично тому, как происходит скольжение миозиновых нитей относительно актиновых. В данном случае наблюдается перемещение головок динеина вдоль микротрубочки от (+)-конца к (-)-концу. При этом соседний дуплет двигается к верхушке реснички. Но так как дуплеты микротрубочек связаны друг с другом и с центральной парой микротрубочек, совместные смещения дуплетов микротрубочек приводят не к удлинению реснички, а к ее изгибу (рис. 296).
Рост ресничек, удлинение микротрубочек их аксонем происходит на вершине реснички. Следовательно, там локализованы (+)-концы микротрубочек.
Образование аксонемы ресничек происходит за счет роста А- и В-микротрубочек центриолей, при этом материнская центриоль подходит к плазматической мембране своим дистальным торцом, связывается с ней своими придатками. В это время начинается рост микротрубочек на (+)- концах А- и В-микротрубочек триплетов. Возникают девять дублетов микротрубочек аксонемы, которые, наращиваясь с (+)-концов на верхушке аксонемы как бы вытягивают плазматическую мембрану, образуя вырост – ресничку. Две центральные микротрубочки возникают в связи с плотным веществом, лежащим на границе бывшей центриоли и выроста плазматической мембраны (рис. 290а).
При образовании многоресничных клеток происходит многочисленная репликация центриолей и образование многочисленных ресничек.
В ресничном эпителии позвоночных множественные базальные тельца возникают вокруг так называемых дейтеросом – аморфных электронноплотных структур размером от 60 до 700 нм, по периферии которых происходит закладка множественных зачатков базальных телец. Вокруг одной дейтеросомы образуются до десятка новых базальных телец. Они затем мигрируют к плазматической мембране и принимают участие в образовании аксонем (рис. 298).
Необходимо отметить, что клетки с множеством ресничек теряют способность к делению и не могут выходить из G0-стадии клеточного цикла. На смену им из эпителиального пласта приходят стволовые недифференцированные клетки, которые могут делиться и давать новые поколения многоресничных клеток.
Вторая категория ресничных клеток – клетки с так называемыми первичными ресничками, не обладающими способностью к движению. Практически все типы клеток, за исключением клеток крови, мышц и кишечного эпителия, в G0-периоде образуют первичные реснички, которые отличаются от настоящих ресничек, или киноцилий, тем, что они не имеют пары центральных микротрубочек и не способны к движению. Они образуются в результате того, что диплосома подходит к плазматической мембране и от материнской центриоли начинается рост аксонемы, но без двух центральных микротрубочек.
У многих клеток животных, вышедших из клеточного цикла, в G0-стадии центриоли принимают участие в образовании аппарата движения – ресничек. Их две группы: кинетоцилии, характерные для специальных эпителиев (ресничные эпителии трахеи, яйцеводов) или свободно плавающих клеток (сперматозоиды, простейшие), и так называемые первичные реснички, встречающиеся во многих клетках, не обладающих способностью к движению.
Клетки, имеющие реснички или жгутики, обладают способностью двигаться, будучи в свободном состоянии, или же перемещать жидкости в случае, если клетки неподвижны. Свободноживущие одноклеточные организмы, снабженные одним или несколькими жгутиками, обычно движутся тем концом вперед, который несет жгутики. У спермиев некоторых животных: жгутик, располагаясь сзади, толкает тело клетки вперед. Скорость движения клеток за счет работы жгутиков может достигать очень большой величины (до 5 мм / мин).
Множественные реснички обеспечивают движение свободноживущих клеток, таких как инфузории или некоторые жгутиконосцы. Реснички эпителиальных клеток многих беспозвоночных и позвоночных животных обеспечивают поток жидкостей вдоль поверхности таких клеток. Число ресничек на клетку может достигать 300 в эпителии трахеи; у инфузории туфельки на клетку приходится 10-14 тыс. рядами расположенных ресничек.
При движении ресничек и жгутиков не происходит уменьшения их длины, поэтому неправильно называть это движение сокращением. Траектория движения ресничек очень разнообразна (рис. 288). В различных клетках это движение может быть маятникообразным, крючкообразным, воронкообразным или волнообразным.
У многоресничных клеток (инфузории, клетки ресничного эпителия) движение ресничек не хаотично, а строго упорядочено. В этом случае реснички расположены рядами. В продольном ряду отдельные реснички начинают движение и проходят отдельные его фазы по очереди, метахронно. В поперечном же ряду все реснички находятся в одной фазе движения (синхронны). Это создает движущую волну по поверхности клетки (рис. )289.
Ресничка представляет собой тонкий цилиндрический вырост цитоплазмы с постоянным диаметром 300 нм. Этот вырост от основания до самой его верхушки покрыт плазматической мембраной. Внутри выроста расположена аксонема, сложная структура, состоящая в основном из микротрубочек. Нижняя, проксимальная часть реснички, базальное тельце, погружена в цитоплазму. Диаметры аксонемы и базального тельца одинаковы (около 200 нм).
На поперечном сечении реснички видна плазматическая мембрана, окружающая аксонему. Аксонема в своем составе имеет девять дублетов микротрубочек, образующих внешнюю стенку цилиндра аксонемы. Дублеты микротрубочек слегка повернуты (около 100) по отношению к радиусу аксонемы. В центре аксонемы располагается пара центральных микротрубочек. В целом систему микротрубочек реснички описывают как (9х2)+2. В дублетах микротрубочек также различают А-микротрубочку, состоящую из 13 субъединиц, и В-микротрубочку, неполную, содержащую 11 субъединиц. А-микротрубочка несет на себе ручки, которые направлены к В-микротрубочке соседнего дуплета. От А-микротрубочки к центру аксонемы отходит радиальная связка, или спица, оканчивающаяся головкой, присоединяющейся к центральной муфте, имеющей диаметр около 70 нм, окружающей две центральные микротрубочки. Последние лежат отдельно друг от друга на расстоянии около 25 нм. Таким образом, в аксонеме располагается 20 продольных микротрубочек, в то время как в базальном тельце их 27 (рис. 291, 292).
Базальное тельце состоит из 9 триплетов микротрубочек (как и центриоль), имеет ручки, втулку и спицы, расположенные в проксимальной (нижней) ее части. На участке базального тельца, примыкающем к плазматической мембране, есть девять придатков, выступов, идущих от каждого триплета микротрубочек к плазматической мембране и связывающих его с клеточной поверхностью.
Базальное тельце и аксонема структурно связаны друг с другом и составляют единое целое: А- и В-микротрубочки триплетов базального тельца продолжаются в А- и В-микротрубочках дуплетов аксонемы. Однако внутренние части аксонемы и базального тельца значительно отличны друг от друга. В зоне перехода базального тела в аксонему наблюдают аморфную поперечную пластинку, которая как бы отделяет эти две части. Центральные микротрубочки аксонемы начинаются от этой пластинки так же, как в этом месте начинается и центральная муфта (капсула) (рис. 290).
В основании ресничек и жгутиков часто встречаются исчерченные корешки, или кинетодесмы, представляющие собой пучки тонких (6 нм) фибрилл, обладающих поперечной исчерченностью (рис. 293). Кинетодесмы простираются от базальных телец вглубь цитоплазмы по направлению к ядру.
При движении ресничек не происходит изменения их длины, они не “сокращаются”, а изгибаются, бьются. Оказалось, что механически отделенные реснички способны к биению в присутствии АТФ. При отделении ресничек базальные тельца остаются в теле клетки. Для механической работы ресничек базальное тело не нужно, а только аксонема участвует в генерации движения. Движение ресничек обеспечивают “ручки”, сидящие на А-микротрубочках.
Установлено, что в состав ручек входят белки динеины. Это большие белковые компоненты, состоящие из 9-12 полипептидных цепей, содержащие 2-3 глобулярные головки, связанные в общий корешок гибкими хвостами (рис. 294). Каждая головка динеина обладает АТФ-азной активностью, которая возрастает примерно в 6 раз при ассоциации с микротрубочками. В состав каждой ручки входит один белковый комплекс, одна молекула динеина. Именно динеин ответственен за это движение, он является мотором или двигателем при биении ресничек. Механизм этого движения следующий.
За счет смещения пар микротрубочек одна относительно другой (рис. 295), аналогично тому, как происходит скольжение миозиновых нитей относительно актиновых. В данном случае наблюдается перемещение головок динеина вдоль микротрубочки от (+)-конца к (-)-концу. При этом соседний дуплет двигается к верхушке реснички. Но так как дуплеты микротрубочек связаны друг с другом и с центральной парой микротрубочек, совместные смещения дуплетов микротрубочек приводят не к удлинению реснички, а к ее изгибу (рис. 296).
Рост ресничек, удлинение микротрубочек их аксонем происходит на вершине реснички. Следовательно, там локализованы (+)-концы микротрубочек.
Образование аксонемы ресничек происходит за счет роста А- и В-микротрубочек центриолей, при этом материнская центриоль подходит к плазматической мембране своим дистальным торцом, связывается с ней своими придатками. В это время начинается рост микротрубочек на (+)- концах А- и В-микротрубочек триплетов. Возникают девять дублетов микротрубочек аксонемы, которые, наращиваясь с (+)-концов на верхушке аксонемы как бы вытягивают плазматическую мембрану, образуя вырост – ресничку. Две центральные микротрубочки возникают в связи с плотным веществом, лежащим на границе бывшей центриоли и выроста плазматической мембраны (рис. 290а).
При образовании многоресничных клеток происходит многочисленная репликация центриолей и образование многочисленных ресничек.
В ресничном эпителии позвоночных множественные базальные тельца возникают вокруг так называемых дейтеросом – аморфных электронноплотных структур размером от 60 до 700 нм, по периферии которых происходит закладка множественных зачатков базальных телец. Вокруг одной дейтеросомы образуются до десятка новых базальных телец. Они затем мигрируют к плазматической мембране и принимают участие в образовании аксонем (рис. 298).
Необходимо отметить, что клетки с множеством ресничек теряют способность к делению и не могут выходить из G0-стадии клеточного цикла. На смену им из эпителиального пласта приходят стволовые недифференцированные клетки, которые могут делиться и давать новые поколения многоресничных клеток.
Вторая категория ресничных клеток – клетки с так называемыми первичными ресничками, не обладающими способностью к движению. Практически все типы клеток, за исключением клеток крови, мышц и кишечного эпителия, в G0-периоде образуют первичные реснички, которые отличаются от настоящих ресничек, или киноцилий, тем, что они не имеют пары центральных микротрубочек и не способны к движению. Они образуются в результате того, что диплосома подходит к плазматической мембране и от материнской центриоли начинается рост аксонемы, но без двух центральных микротрубочек.
БАЗА́ЛЬНОЕ ТЕ́ЛЬЦЕ (кинетосома), внутриклеточная структура эукариот, лежащая в основании ресничек и жгутиков; обеспечивает закрепление ресничек и жгутиков в клетке, а также рост их стержня – аксонемы. Б. т. имеет форму короткого цилиндра, стенки которого состоят из 9 троек параллельно расположенных микротрубочек . Ультраструктура Б. т. сходна с ультраструктурой центриоли . В ресничном эпителии позвоночных, напр., Б. т. образуется путём удвоения центриолей, у многожгутиковых и папоротниковидных – из агрегатов плотного материала неизвестной природы, т. н. дейтеросом.
Различают по меньшей мере три типа таких структур: микротрубочки, микрофиламенты и промежуточные филаменты. Их функции связаны с внутриклеточным движением, со способностью клеток поддерживать свою форму, а также с некоторыми другими видами активности клеток, такими, например, как эндоцитоз и экзоцитоз. Мы рассмотрим здесь только микротрубочки.
Микротрубочки содержатся почти во всех эукариотических клетках. Это полые, очень тонкие неразветвленные трубочки диаметром приблизительно 24 нм; их стенки толщиной около 5 нм построены из спирально упакованных субъединиц белка тубулина.
Рисунок дает представление о том, как выглядят микротрубочки на электронных микрофотографиях. Растут микротрубочки с одного конца путем добавления тубулиновых субъединиц. Рост видимо, может начаться лишь при наличии матрицы; есть основания полагать, что роль таких матриц играют какие-то очень мелкие кольцевые структуры, которые были выделены из клеток и которые, как выяснилось, состоят из тубулиновых субъединиц. В интактных клетках ту же функцию выполняют центриоли, поэтому их иногда называют центрами организации микротрубочек (ЦОМ). Центриоли состоят из коротких микротрубочек.
Микротрубочки принимают участие в различных внутриклеточных процессах; некоторые мы здесь упомянем.
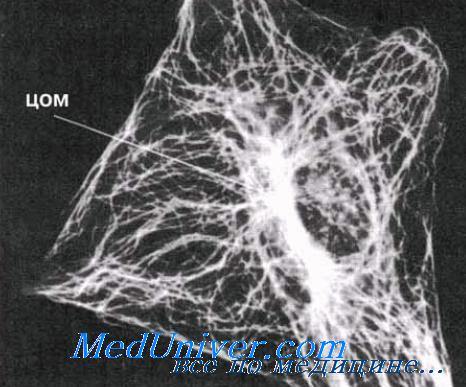
Распределение микротрубочек в клетке. Микротрубочки расходятся от центра организации микротрубочек (ЦОМ), находящегося рядом с ядром. В ЦОМ содержится центриоль. Микротрубочки видны на этой микрофотографии благодаря использованию флуоресцирующих антител, способных специфически соединяться с их белком. Представленная здесь клетка — фибробласт; фибробласты обычно содержатся в соединительной ткани; в них синтезируется коллаген.
Центриоли и деление ядра
Микротрубочки регулируют расхождение хроматид или хромосом. Осуществляется это за счет скольжения микротрубочек. В клетках высших растений центриоли отсутствуют, хотя веретено в них при делении ядра образуется. Возможно, что в этих клетках имеются какие-то очень мелкие центры организации микротрубочек, не выявляемые даже при помощи электронного микроскопа.
Базальные тельца, реснички и жгутики
В ресничках и жгутиках движение осуществляется за счет скольжения микротрубочек. Более подробно эти процессы описаны в нашей статье. Отметим, что жгутики бактерий устроены проще, чем жгутики эукариот, и базальные тельца у них отсутствуют.
Внутриклеточный транспорт
Микротрубочки участвуют также в перемещении различных клеточных органелл, например в перемещении пузырьков Гольджи к формирующейся клеточной пластинке (рис. 5.30). В клетке идет непрерывный транспорт: перемещаются пузырьки Гольджи, направляются к аппарату Гольджи пузырьки, отпочковывающиеся от ЭР, движутся лизосомы, митохондрии и другие органеллы. Все это движение приостанавливается, если повреждена система микротрубочек.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Читайте также: