
Анафаза это в биологии кратко
Обновлено: 05.07.2024
Анафаза
(от греч. aná — вверх и фаза) , третья стадия, или фаза, митотического деления (митоза) растительной и животной клетки. Во время А. составляющие каждую хромосому хроматиды (или сестринские хромосомы) разъединяются и расходятся к противоположным полюсам клетки. При нормальном митозе движение всех хромосом синхронно; оно осуществляется при взаимодействии двух одновременно протекающих процессов: сокращения нитей, связывающих хромосомы с полюсами; удлинения центральных нитей веретена, связывающих оба полюса.
Метафа́за — фаза митотического деления эукариотических клеток, начало которой знаменует выход хромосом в экваториальную плоскость клетки.
Телофа́за — фаза митотического деления эукариотических клеток, во время которой два набора дочерних хромосом достигают полюсов веретена деления и деконденсируются. Начинается сборка ядерной оболочки вокруг каждого набора хромосом. Разделение цитоплазмы достигается путём сокращения сократительного кольца (цитокинез).
Се́стринские хромати́ды — идентичные хроматиды, образовавшиеся в результате репликации хромосомы и соединенные в области центромеры. Они образуются в S-фазе интерфазы, когда происходит удвоение ДНК, и разделяются во время митоза и второго деления мейоза. В дальнейшем в каждую дочернюю клетку попадает по одной такой хроматиде из пары хроматид данной хромосомы, и каждая из них достраивает себе пару.
Интерфа́за (англ. interphase) — период клеточного цикла, подразделяющийся на G1-, S- и G2-фазы. Во время интерфазы клетка готовится к будущему делению: растёт, удваивает количество цитоплазмы, клеточных белков и органелл. В S-фазе происходит удвоение хромосом и центросом (клеточных центров).
Профа́за (англ. prophase) — самая первая фаза митоза, признаком которой является появление в ядре конденсированных хромосом.
Упоминания в литературе
Анафаза начинается с синхронного расщепления всех хромосом на сестринские хроматиды (в области центромера) и движения дочерних хромосом к противоположным полюсам клетки. Характеризуется удлинением митотического веретена за счет некоторого расхождения полюсов клетки. Завершается скоплением на полюсах клетки двух идентичных наборов хромосом (стадия дочерних звезд).
Вслед за телофазой первого деления следует короткая интерфаза, в которой не происходит синтеза ДНК и клетки приступают к следующему делению, почти сразу после окончания первого. Поэтому на препаратах сперматоциты 2-го порядка почти не встречаются, время их жизни очень мало. Второе деление мейоза по морфологии и последовательности не отличается от митотического деления: парные сестринские хроматиды, связанные в центромерных участках, проходят профазу и метафазу; в анафазе они разъединяются и расходятся по одной в дочерние клетки. Таким образом, при втором мейотическом делении клетка с 2с количеством ДНК и 2n числом хроматид, делясь, дает начало двум клеткам с гаплоидным содержанием ДНК и хромосом. В отношении числа структурных единиц, хроматид, второе деление является редукционным. В результате всего процесса мейоза после двух делений из одной клетки образуется четыре гаплоидных сперматозоида, каждый из которых отличается по своей генетической конституции [76, 79].
Метафаза I напоминает аналогичную стадию митоза, хромосомы устанавливаются в экваториальной плоскости. В анафазе I гомологичные хромосомы отделяются друг от друга и расходятся к полюсам. В телофазе I наборы гомологичных хромосом находятся у полюсов, хотя их число уменьшилось вдвое, но каждая из них состоит уже из двух генетически различных хроматид. Формируются ядерная оболочка и ядрышко, образуются борозда деления, которая углубляется, и две клетки, каждая из которых содержит гаплоидный набор удвоенных хромосом, полностью разделяются.
Центриоли поляризуют процесс деления клетки, чем достигается равномерное расхождение сестринских хромосом (хроматид) в анафазе митоза.
Белок Сdt1 также инактивируется после завершения формирования рге-RС. При этом в клетках низших эукариот Cdt1. периодически экспрессируется, накапливаясь в ядрах в фазе G1, а в фазе G2 его уровень падает. У высших эукариот активность Сdt1 контролируется белком-ингибитором геминином, который отсутствует в клетках в фазе G1, накапливается на протяжении фаз S, G2 и частично – фазы М и исчезает в М-фазе на границе метафазы и анафазы .
Анафаза характеризуется полным обособлением (расхождением) хроматид и образованием двух равноценных диплоидных наборов хромосом, расхождением хромосомных наборов к полюсам митотического веретена и расхождением самих полюсов.
Связанные понятия (продолжение)
Кинетохор — белковая структура на хромосоме, к которой крепятся волокна веретена деления во время деления клетки. Кинетохоры играют важнейшую роль при сегрегации хромосом для последующего разделения родительской клетки на две дочерние.Кинетохоры формируются на центромерах хромосом у эукариотов. Кинетохоры подразделяют на две области — внутреннюю, крепко связанную с центромерной ДНК, и внешнюю, взаимодействующую с микротрубочками веретена деления.
Центромера — участок хромосомы, который связывает сестринские хроматиды, играет важную роль в процессе деления клеточного ядра и участвует в контроле экспрессии генов. Характеризуется специфическими последовательностью нуклеотидов и структурой.
Прометафа́за (англ. prometaphase) — вторая фаза митотического процесса деления клетки. Прометафаза начинается внезапно с быстрого разрушения ядерной оболочки. Прометафаза заканчивается, когда все хромосомы оказываются в экваториальной плоскости веретена деления.
Веретено́ деле́ния — динамичная структура, которая образуется в митозе и мейозе для обеспечения сегрегации хромосом и деления клетки. Типичное веретено является биполярным — между двумя полюсами образуется веретенообразная система микротрубочек. Микротрубочки веретена присоединяются к кинетохорам хроматид в области центромер и обеспечивают движение хромосом по направлению к полюсам.
Цитокинез, цитотомия — деление тела эукариотической клетки. Цитокинез обычно происходит после того, как клетка претерпела деление ядра (кариокинез) в ходе митоза или мейоза. В большинстве случаев цитоплазма и органоиды клетки распределяются между дочерними клетками приблизительно поровну.
Кле́точный цикл — период существования клетки от момента её образования путём деления материнской клетки до собственного деления или гибели.
Центр организации микротрубочек (ЦОМТ, англ. microtubule-organising centre, MTOC) — структура эукариотической клетки, на которой собираются микротрубочки. ЦОМТ имеет две основные функции — сборка жгутиков и ресничек, а также образование нитей веретена деления в ходе митоза и мейоза.
Когези́н — это комплекс белков, который регулирует процесс разделения сестринских хроматид в ходе деления клетки (как мейоза, так и митоза).
Мито́з (др.-греч. μίτος — нить) — непрямое деление клетки, наиболее распространённый способ репродукции эукариотических клеток. Биологическое значение митоза состоит в строго одинаковом распределении хромосом между дочерними ядрами, что обеспечивает образование генетически идентичных дочерних клеток и сохраняет преемственность в ряду клеточных поколений.
Ко́мплекс, стимули́рующий анафа́зу (англ. anaphase-promoting complex, APC), также называемый циклосома, представляет собой крупное белковое соединение, которому отводится решающая роль в активации анафазы митоза. Функционально комплекс стимуляции анафазы представляет собой убиквитинлигазу и катализирует реакции присоединения молекул убиквитина к различным целевым белкам, которые в итоге подвергаются протеолизу.
Конденсины — большие белковые комплексы, которые играют главную роль в расхождении хромосом во время митоза и мейоза.
Хромати́да (греч. chroma — цвет, краска и греч. eidos — вид) — структурный элемент хромосомы, формирующийся в интерфазе ядра клетки в результате удвоения хромосомы.
Гомологи́чные хромосо́мы — пара хромосом приблизительно равной длины, с одинаковым положением центромеры и дающие одинаковую картину при окрашивании. Их гены в соответствующих (идентичных) локусах представляют собой аллельные гены — аллели, то есть кодируют одни и те же белки или РНК. При двуполом размножении одна гомологичная хромосома наследуется организмом от матери, а другая — от отца.
Циклины — семейство белков-активаторов циклин-зависимых протеинкиназ (CDK) (англ. CDK, cyclin-dependent kinases) — ключевых ферментов, участвующих в регуляции клеточного цикла эукариот. Циклины получили своё название в связи с тем, что их внутриклеточная концентрация периодически изменяется по мере прохождения клеток через клеточный цикл, достигая максимума на его определенных стадиях.
Нокдаун гена (англ. Gene knockdown) — методика, позволяющая снизить экспрессию одного или нескольких генов при помощи изменения соответствующей последовательности нуклеотидов, либо при помощи короткого олигонуклеотида, комплементарного соответствующей молекуле мРНК. Метод нокдауна генов относится к методам обратной генетики. В случае, когда изменяется последовательность гена, организм называют нокаутным по данному гену. В случае использования коротких олигонуклеотидов, комплементарных соответствующим.
Деле́ние кле́тки — процесс образования из родительской клетки двух и более дочерних клеток. Обычно деление клетки - это часть большего клеточного цикла.
Нуклеосома — это структурная часть хромосомы, образованная совместной упаковкой нити ДНК с гистоновыми белками H2А, H2B, H3 и H4. Последовательность нуклеосом, соединенная гистоновым белком H1, формирует нуклеофиламент, или иначе нуклеосомную нить.
Дупликация (лат. duplicatio — удвоение) — разновидность хромосомных перестроек, при которой участок хромосомы оказывается удвоенным. Может произойти в результате неравного кроссинговера, ошибки при гомологичной рекомбинации, ретротранспозиции.
Я́дерные спе́клы, или спе́клы, или B-снурпосо́мы, или компа́ртменты фа́кторов спла́йсинга, или доме́ны SC-35, или кла́стеры интерхромати́новых гра́нул (англ. nuclear speckles) — ядерные тельца, регулирующие сплайсинг. В ядерных спеклах находятся факторы сплайсинга и малые ядерные рибонуклеопротеины. Изменения в белковом составе и функционировании ядерных спекл приводят к изменениям в альтернативном сплайсинге пре-мРНК, поэтому считается, что ключевой функцией ядерных спекл является регуляция доступности.
Подавление экспрессии генов (сайленсинг генов от англ. gene silencing, или в частности, выключение гена) — это общий термин, описывающий эпигенетический процесс регуляции генов. При этом последовательность нуклеотидов не изменяется, а лишь прекращается экспрессия соответствующего гена. Для выключения генов в лабораторных условиях применяют метод нокдауна генов.
Ядерные поры, или ядерные поровые комплексы, — крупные белковые комплексы, пронизывающие ядерную мембрану и осуществляющие транспорт макромолекул между цитоплазмой и ядром клетки. Переход молекул из ядра в цитоплазму и в обратном направлении называется ядерно-цитоплазматическим транспортом.
Те́льце гисто́новых ло́кусов (англ. Histone Locus Body) — ядерное тельце, представляющее собой скопление факторов транскрипции генов, кодирующих гистоны, и созревания гистоновых пре-мРНК. К числу таких факторов относят белок NPAT, необходимый для транскрипции генов гистонов, а белок FLASH и малый ядерный рибонуклеопротеин U7 необходим для процессинга гистоновых пре-мРНК. Тельца гистоновых локусов обнаружены в клетках млекопитающих и дрозофилы. В клетках амфибий аналогичные ядерные тельца носят название.
Гисто́ны — обширный класс ядерных белков, выполняющих две основные функции: они участвуют в упаковке нитей ДНК в ядре и в эпигенетической регуляции таких ядерных процессов, как транскрипция, репликация и репарация. Существует пять различных типов гистонов H1/Н5, H2A, H2B, H3, H4. Гистоны H2A, H2B, H3, H4, называемые кóровыми гистонами (от англ. core — сердцевина), формируют нуклеосому, представляющую собой белковую глобулу, вокруг которой накручена нить ДНК. Гистон H1/H5, называемый линкерным гистоном.
Цис-регуляторные элементы (или цис-элементы) — участки ДНК или РНК, регулирующие экспрессию генов, находящихся на той же молекуле (обычно хромосоме).
Тубули́н — белок, из которого построены микротрубочки. В них, а также в цитоплазме клеток он находится в форме димера из одной молекулы α-тубулина и одной — β-тубулина. В составе такого димера к каждой молекуле тубулина присоединена молекула ГТФ. У каждой из этих субъединиц выделяют три домена. Форма γ-тубулина принимает участие в нуклеации микротрубочек, то есть образовании затравки, с которой начинается рост. Тубулин способен связывать в растворе молекулы ГТФ. Рост микротрубочек осуществляется.
То́чка рестри́кции (англ. restriction point, R point) — точка в G1-фазе клеточного цикла животных, в которой клетка принимает окончательное решение двигаться дальше по клеточному циклу. Точка рестрикции делит G1-фазу на два периода. В ходе первого из них (до R) для продвижения клетки по циклу ей необходима постоянная стимуляция митогенными сигналами (в том числе факторами роста), а также интенсивный синтез белка. После прохождения R подобная стимуляция уже не требуется.
Сплайсосо́ма — структура, состоящая из молекул РНК и белков и осуществляющая удаление некодирующих последовательностей (интронов) из предшественников мРНК. Этот процесс называется сплайсингом (от англ. splicing — сращивание).
Транскрипт — молекула РНК, образующаяся в результате транскрипции (экспрессии соответствующего гена или участка ДНК).
Гомеодомен — это структурный домен белков, связывающих ДНК или РНК, широко распространенный среди факторов транскрипции. Домен состоит из 60 остатков аминокислот, и образует структуру спираль-поворот-спираль, в которой альфа-спирали связаны короткими петлевыми участками. Две спирали на N-конце являются антипараллельными, и длиннее спирали на C-конце, которая перпендикулярна осям N-концевым петлям. Непосредственно С-концевая спираль взаимодействует с ДНК. Укладка доменов белков по типу гомеодомена.
Белковая субъединица в структурной биологии — полипептид, который вместе с другими компонентами собирается в мультимерный или олигомерный белковый комплекс. Многие природные ферменты и другие белки состоят из нескольких белковых субъединиц.
Процессинг РНК (посттранскрипционные модификации РНК) — совокупность процессов в клетках эукариот, которые приводят к превращению первичного транскрипта в зрелую РНК.
Эндонуклеазы — белки из группы нуклеаз, расщепляющие фосфодиэфирные связи в середине полинуклеотидной цепи. Эндонуклеазы рестрикции, или рестриктазы, расщепляют ДНК в определенных местах (так называемых сайтах рестрикции), они подразделяются на три типа (I, II и III) на основании механизма действия. Эти белки часто используют в генной инженерии для создания рекомбинантных ДНК, которые вводят затем в бактериальные, растительные или животные клетки.
Точка начала репликации (англ. origin of replication) — это фрагмент молекулы нуклеиновой кислоты, с которого начинается её репликация. Структура точки начала репликации (нуклеотидная последовательность) отличается у разных видов, но у всех организмов это АТ-богатая и потому легкоплавкая последовательность. Точка начала репликации и прилегающие к ней фрагменты нуклеиновой кислоты, не отделённые сайтами терминации, составляют единицу репликации — репликон. Репликация ДНК может начинаться от точки.
ГТФазы (англ. GTPases) — большое семейство ферментов гидролаз, которые связывают и гидролизуют гуанозинтрифосфат (GTP). Связывание и гидролиз GTP осуществляются в консервативном G-домене, сходном для всех ГТФаз.
Клеточный центр, или центросома (от др.-греч. σῶμα — тело) — немембранный органоид в клетках эукариот, состоит из двух центриолей и перицентриолярного материала. Является главным центром организации микротрубочек (ЦОМТ) эукариотической клетки, играет важнейшую роль в клеточном делении, участвуя в формировании веретена деления. Из центросомы образуются реснички и жгутики. Центросомы характерны для клеток животных, их нет у высших растений, у высших грибов, у некоторых простейших. Ряд наследственных.
Транслока́ция — тип хромосомных мутаций, при которых происходит перенос участка хромосомы на негомологичную хромосому. Отдельно выделяют реципрокные транслокации, при которых происходит взаимный обмен участками между хромосомами, и Робертсоновские транслокации, или центрические слияния, при которых происходит слияние акроцентрических хромосом с полной или частичной утратой материала коротких плеч.
Цитоскеле́т прокарио́т — совокупное название для всех структурных филаментов прокариот. В прошлом считалось, что у прокариот цитоскелета нет, однако с начала 1990-х стали накапливаться данные о наличии у прокариот разнообразных филаментов. У прокариот не только имеются аналоги ключевых белков цитоскелета эукариот, но и белки, не имеющие аналогов у эукариот. Элементы цитоскелета играют важные роли в делении клеток, защите, поддержании формы и определении полярности у различных прокариот.
Я́дерная мембра́на, или ядерная оболо́чка, или кариоле́мма, или кариоте́ка, или нуклеоле́мма — двойной липидный бислой, мембрана, окружающая ядро в эукариотических клетках.
Экзонуклеазы — белки из группы нуклеаз, отщепляющие концевые мононуклеотиды от полинуклеотидной цепи путём гидролиза фосфодиэфирных связей между нуклеотидами.
Нуклео́ид (англ. Nucleoid) — неправильной формы зона в цитоплазме прокариотической клетки, в которой находится геномная ДНК и ассоциированные с ней белки. На долю ДНК приходится около 60 % массы нуклеоида; помимо ДНК, нуклеоид содержит РНК и белки. Белки нуклеоида, которые обеспечивают пространственную организацию геномной ДНК, называют нуклеоидными белками или нуклеоид-ассоциированными белками; они не имеют ничего общего с гистонами, упаковывающими ДНК у эукариот. В отличие от гистонов, ДНК-связывающие.
Шаперо́н (англ. chaperones) — класс белка, главная функция которого состоит в восстановлении правильной нативной третичной или четвертичной структуры белка, а также образование и диссоциация белковых комплексов.
В биохимии, димер — макромолекулярный комплекс, образованный двумя, как правило, не ковалентносвязаными макромолекулами, такими как белки или нуклеиновые кислоты. Белковый димер — это четвертичная структура белка.
Анафаза – стадия во время эукариотическая клетка деление, при котором хромосомы выделяются на противоположных полюсах клетка, Этап перед анафазой, метафазы хромосомы тянутся к метафазной пластинке в середине клетки. Хотя хромосомы были сильно конденсированы в начале деление клеток они продолжают конденсироваться через анафазу. Анафаза начинается после того, как клетка проходит контрольную точку формирования веретена, что позволяет разделить хромосомы или хроматиды. По мере укорочения микротрубочек, которые соединяют хромосомы с центросомами, хромосомы тянутся к центросома пока они не образуют полукруг вокруг него. На следующем этапе деления клеток, телофаза клетка реформирует ядро и готовится к делению.
Контрольная точка формирования шпинделя происходит до начала анафазы. Этот клеточный механизм гарантирует, что все хромосомы связаны с микротрубочками и выровнены на метафазной пластине. Как только этот шаг происходит, клетка выпускает сигнал, который создает анафазо-стимулирующий комплекс или APC, вещество, которое будет действовать, чтобы начать процесс деления гомологичные хромосомы или сестринские хроматиды в зависимости от того клеточный цикл происходит. APC, как видно на графике ниже, разлагает секурин, ингибирующий молекула что останавливает действие сепарации. Как только отделяется, он может воздействовать на cohesins, которые удерживают хроматиды вместе. Cohesins – это макромолекулы, состоящие из нескольких белков. Когда эти белки расщепляются по отдельности, хроматиды распадаются. В первом дивизионе мейоз, гомологичны хромосомы удерживаются вместе cohesins, которые разрушаются во время Анафаза I.

Микротрубочки остаются прикрепленными к кинетохоре после разрыва когезинов. Микротрубочки кинетохоры затем сжимаются к центросомам (не показано), что разделяет хромосомы. Не кинетохорные микротрубочки растут в противоположном направлении, расширяя клетку и далее разделяя хроматиды. Обычно сестринские хроматиды называются сестринскими хромосомами после их разделения, поскольку они содержат одинаковую информацию и будут независимо функционировать в своих новых клетках. После полного разделения хромосом образуется ядерная оболочка и цитоплазма будут разделены на заключительных этапах деления клеток.
Анафаза в Митозе
Митоз это процесс, используемый клетками для точного копирования себя. Через митоз, два новых дочерние клетки создаются из одного родителя, каждый из которых идентичен родителю. Перед митозом хромосомы, содержащие ДНК, реплицируются, а реплицированные сестринские хроматиды остаются прикрепленными. Перед анафазой хромосомы конденсируются, волокна шпинделя образуются из микротрубочек, и хромосомы выравниваются на метафазной пластинке. Сестринские хроматиды начинают отделяться в начале анафазы, когда разделение начинает разрушать когезин, который связывает их вместе. Анафаза заканчивается, когда телофаза и цитокинез начать, как реформы ядерной оболочки и хромосомы начинают раскручиваться. Как только они разболтались и клетки были разделены, они снова могут начать функционировать самостоятельно. Это отмечает конец деления клеток и начало интерфаза.
Анафаза в мейозе
Анафаза I
Мейоз состоит из двух последовательных клеточных делений, между которыми нет реплицированной ДНК. Это означает, что диплоид организмы, содержащие два аллеля для каждого ген, будет уменьшен до гаплоидный организм только с одним аллель на каждом гене. Эти аллели разделяются во время анафазы I. До мейоза ДНК дублируется, снова продуцируя сестринские хроматиды, связанные вместе как единые хромосомы. Эти хромосомы имеют гомологичные пары, которые содержат другие аллели для генов на хромосома, Эти хромосомы также дублируются в сестринские хроматиды. Во время мейоза I гомологичные хромосомы разделены.
Гомологичные хромосомы становятся связанными во время первая фаза мейоза, с помощью похожих молекул cohesin, которые связывают сестринские хроматиды. Как мейотический шпиндель устанавливается во время метафазе I клетка обеспечивает присоединение каждой гомологичной пары к микротрубочкам с каждой стороны клетки. Эти микротрубочки затем притягиваются друг к другу, чтобы переместить гомологичные пары в метафазную пластинку. В течение времени, когда пары связаны, они могут обмениваться генетической информацией в процессе, называемом рекомбинацией. Когда гомологичные пары разделяются во время анафазы I, вариации ДНК становятся предназначенными для разных клеток, обеспечивая изменчивость в Население «s генетика.
Анафаза II
Мейоз завершается вторым делением каждой новой дочерней клетки. Проходят те же этапы, на этот раз более похожим на митоз. Хромосомы, все еще сделанные из сестринских хроматид, выравниваются на метафазной пластинке и должны пройти контрольную точку, чтобы перейти к анафазе II. В начале анафазы II сестринские хроматиды отделяются, когда связывающий их когезин высвобождается. Затем копии хромосомы отделяются до их конечного пункта назначения. Эти новые клетки будут давать гаметы, которые могут оплодотворять гамета противоположного пола, чтобы произвести новое потомство.
Если клеточные контрольные точки, ведущие к анафазе I или анафазе II, терпят неудачу, конечным результатом будет слишком много копий хромосомы (или многих) в конечном гамете. Это состояние, известное как нерасхождения, может привести к фатальным врожденным дефектам и другим симптомам у многих размножающихся половым путем животных. У людей эти состояния включают синдром Дауна, синдром Эдвардса, синдром Клайнфелтера и синдром Тернера. Эти синдромы имеют различные симптомы развития, которые создают фенотипы, которые отличаются от среднего.
- Нерасхождения – Состояние, вызванное неспособностью хромосом отделиться во время любого деления мейоза.
- цитокинез – последняя стадия митоза или мейоза, при которой клеточная мембрана водоразделы.
- Анафазный промоушн комплекс – Система белков, коферментов и других молекул, которые позволяют сепарации разрушать молекулы когезина, что приводит к разделению хромосом.
- Cohesin – Белковые молекулы, которые связывают сестринские хроматиды или гомологичные хромосомы вместе.
викторина
1. Клетка подвергается митозу. Хромосомы разделились и движутся к своим новым ядрам, но ядерные оболочки не изменились. На какой стадии находится клетка?A. профаза B. анафазаC. телофаза
Ответ на вопрос № 1
В верно. Эта клетка находится в анафазе. Телофаза характеризуется реформированием ядерных оболочек и дальнейшим делением органелл и цитозоль, Телофаза завершается, когда цитокинез расщепляет клетку на две части. Линия между анафазой и телофазой является серой, но ядерная оболочка начинает формироваться в начале телофазы. Хромосомы могут все еще перемещаться в их заключительные положения, поскольку это происходит.
2. Анафаза происходит невероятно быстро. Весь процесс составляет всего 1% клеточного цикла. Почему хромосомы не разрываются во время этого насильственного движения?A. Конденсированная ДНК более защищена от вредаB. Длинные сегменты ДНК, называемые теломерами, защищают концы ДНК.C. Оба а и Б
Ответ на вопрос № 2
С верно. ДНК защищена рядом механизмов, только два из которых являются теломерами и способностью конденсироваться. Волокна веретена также защищают ДНК, поскольку они обеспечивают четкий путь в одном направлении для продвижения хромосомы. ДНК также быстро окружается ядерной оболочкой после анафазы, которая защищает ее от остального деления клеток.
3. Дрожжи являются одноклеточным�� эукариотическими организмами. Дрожжи проходят через митоз и в настоящее время находятся в анафазе. Сколько организмов сейчас присутствует и сколько их будет после цитокинеза?A. 2; 4B. 1; 1C. 1; 2
Ответ на вопрос № 3
С верно. Иногда дрожжи могут размножаться половым путем, но это пример того, как они размножаются бесполым путем. Таким образом, после того, как клетка делится, присутствуют два организма, когда только один присутствовал во время анафазы. Это верно для любого одноклеточного организма, который подвергается митозу. Разделение двух клеток является разделительной линией между одним организмом и двумя, поскольку клеточная мембрана физически отделяет содержимое каждой клетки от другой. Это означает, что организмы теоретически независимы, хотя они все еще могут взаимодействовать.
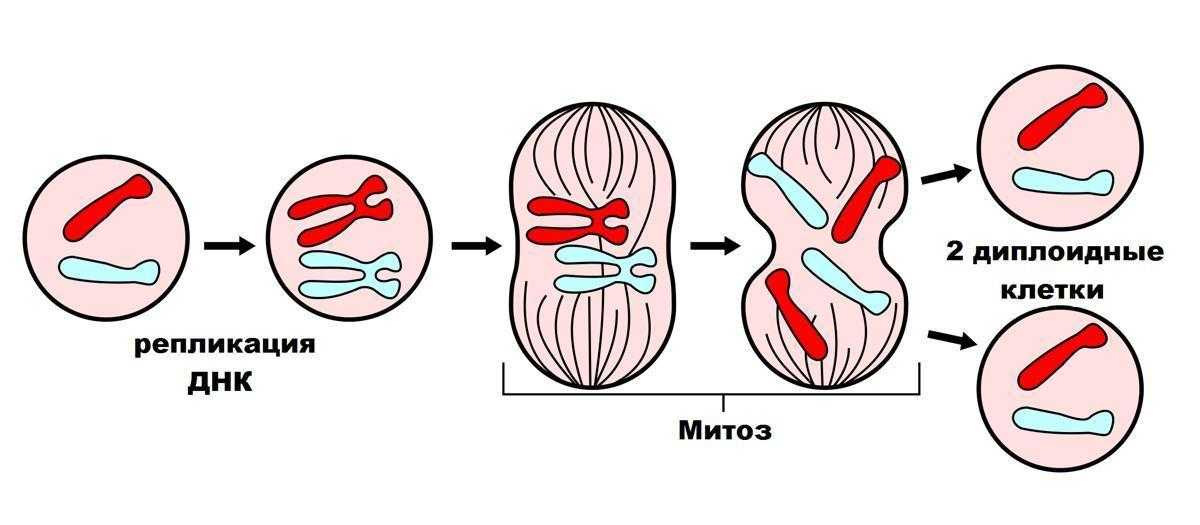
Митоз – процесс деления клеток, при котором хромосомы из ядра материнской клетки равномерно распределены между двумя новыми клетками. После завершения митоза, производятся две дочерние клетки с идентичным генетическим материалом.
Интерфаза
Прежде чем делящаяся клетка попадает в митоз, она подвергается периоду роста, называемому интерфазой. Около 90% времени клетки при нормальном клеточном цикле могут быть потрачены на интерфазу, которая осуществляется в три основные фазы:
- Фаза G1: период до синтеза ДНК. В этой фазе клетка увеличивается в массе, подготавливаясь к делению.
- S-фаза: период, в течение которого происходит синтез ДНК. В большинстве клеток эта стадия происходит за очень короткий промежуток времени.
- Фаза G2: клетка продолжает синтез дополнительных белков увеличиваться в размерах.
В последней части интерфазы, клетка все еще имеет нуклеолы. Ядро ограничено ядерной оболочкой, а хромосомы дублируются, но находятся в форме хроматина. В клетках животных две пары центриолей, образованных из репликации одной пары, расположены за пределами ядра.
После фазы G2 наступает митоз, который в свою очередь состоит из нескольких стадий и завершается цитокинезом (делением клетки).
Фазы митоза:
Препрофаза (в клетках растений)
Препрофаза является дополнительной фазой во время митоза в клетках растений, которая не встречается у других эукариот, таких как животные или грибы. Она предшествует профазе и характеризуется двумя различными событиями.
Изменения, которые происходят в препрофазе:
- Образование полосы препрофазы – плотного микротрубочного кольца под плазматической мембраной.
- Начало зарождения микротрубочек в ядерной оболочке.
Профаза
В профазе хроматин конденсируется в дискретные хромосомы. Ядерная оболочка ломается, а веретено деления образуются на противоположных полюсах клетки. Профаза (по сравнению с интерфазой) является первым истинным шагом митотического процесса.
Изменения, которые происходят в профазе:
- Хроматиновые волокна превращаются в хромосомы, имеющие по две хроматиды, соединенные в центромер. Волокна деления, состоящие из микротрубочек и белков, образуется в цитоплазме.
- В клетках животных волокна деления первоначально появляется как структуры, называемые астерами, которые окружают каждую пару центриолей.
- Две пары центриолей (сформированных из репликации одной пары в интерфазе) отходят друг от друга к противоположным полюсам клетки из-за удлинения микротрубочек, образующихся между ними.
Прометафаза

Прометафаза – фаза митоза после профазы и предшествующая метафазе в эукариотических соматических клетках. Некоторые источники относят процессы протекающие в прометафазе к поздней профазе и начальной стадии метафазы.
Изменения, которые происходят в прометафазе:
Метафаза

В метафазе полностью развиваются волокна деления, а хромосомы выравниваются на метафазной (экваториальной) пластине (плоскость, которая одинаково удалена от двух полюсов).
Изменения, которые происходят в метафазе:
- Ядерная мембрана полностью исчезает.
- В клетках животных две пары центриолей расходятся в противоположных направлениях к полюсам клетки.
- Полярные волокна (микротрубочки, составляющие волокна веретена) продолжают распространяться от полюсов к центру. Хромосомы перемещаются случайным образом, пока не присоединяют (при помощи своих кинетохор) к полярным волокнам с обеих сторон центромеров.
- Хромосомы выравниваются на метафазной пластине под прямым углом к полюсам веретена.
- Хромосомы удерживаются на метафазной пластине равными силами полярных волокон, которые нажимают на их центромеры.
Анафаза
В анафазе парные хромосомы (сестринские хроматиды) отделяются и начинают двигаться к противоположным концам (полюсам) клетки. Волокна веретена, не связанные с хроматидами, вытягиваются и удлиняют клетку. В конце анафазы каждый полюс содержит полную компиляцию хромосом.
Изменения, которые происходят в анафазе:
Телофаза
В телофазе хромосомы достигают ядер новых дочерних клеток.
Изменения, которые происходят в телофазе:
- Полярные волокна продолжают удлиняться.
- Ядра начинают формироваться на противоположных полюсах.
- Ядерные оболочки новых ядер образовываются из остатков ядерной оболочки материнской клетки и кусочков эндомембранной системы.
- Появляются ядрышка.
- Разматываются хроматиновые волокна хромосом.
- После этих изменений телофаза и митоз в основном завершены, а генетическое содержание одной клетки поделено на две части.
Цитокинез
Цитокинез – это разделение цитоплазмы клетки. Он начинается до конца митоза в анафазе и заканчивается вскоре после телофазы. В конце цитокинеза образуются две генетически идентичные дочерние клетки.
Дочерние клетки
В конце митоза и цитокинеза хромосомы распределены поровну между двумя дочерними клетками. Эти клетки являются идентичными диплоидными клетками, причем каждая из которых содержит полный набор хромосом.
Клетки, продуцируемые через митоз, отличаются от клеток, продуцируемых через мейоз. В мейозе образуются четыре дочерние клетки. Эти клетки представляют собой гаплоидные клетки, содержащие половину числа хромосом от исходной клетки. Половые клетки подвергаются мейозу. При делении половых клеток во время оплодотворения, гаплоидные клетки становятся диплоидной клеткой.
Читайте также: