
Влияние внешних условий на фотосинтез кратко
Обновлено: 02.07.2024
- Для учеников 1-11 классов и дошкольников
- Бесплатные сертификаты учителям и участникам
ПРОЦЕССЫ АССИМИЛЯЦИИ И ДИССИМИЛЯЦИИ В РАСТЕНИЯХ
ФОТОСИНТЕЗ: СУЩНОСТЬ И ЗНАЧЕНИЕ.
МАТЕРИАЛЬНАЯ БАЗА, ХИМИЗМ И ЭНЕРГЕТИКА ФОТОСИНТЕЗА.
ВЛИЯНИЕ ВНУТРЕННИХ И ВНЕШНИХ ФАКТОРОВ НА ФОТОСИНТЕЗ.
ФОТОСИНТЕЗ И УРОЖАЙ
Сущность и значение фотосинтеза
Подобно другим организмам зеленые растения в качестве источника энергии используют углеводы и другие органические вещества; однако в отличие от большинства организмов зеленые растения — автотрофы (питающиеся самостоятельно). Растения создают свою пищу сами в процессе фотосинтеза. Гетеротрофы (животные, грибы и большинство бактерий) питаются готовым органическим веществом.
Фотосинтез — процесс образования зелеными растениями органических веществ из неорганических (диоксида углерода и воды) при участии световой энергии, поглощенной хлорофиллом. В процессе фотосинтеза за счет расщепления (фотолиза) воды в атмосферу выделяется кислород:
![]()
В ходе этого синтеза лучистая энергия Солнца превращается в химическую энергию органических соединений. Посредством фотосинтеза зеленые растения обеспечивают космическую связь жизни на Земле с Вселенной и определяют экологическое благосостояние нашей планеты. По современным данным, на Земле за счет фотосинтеза ежегодно синтезируется около 100 млрд т сухого органического вещества, которое примерно поровну распределяется между сушей и морями. Из наземной растительности на долю лесов приходится 24,4 млрд т сухой органической массы, на долю пастбищ — 10,4, на пашню — 8,7, на пустыни и полупустыни — 5,4 млрд т. Благодаря фотосинтетической деятельности растений в атмосфере Земли содержание диоксида углерода поддерживается примерно на уровне 0,03%, а кислорода — 21%, Фотосинтез — это не только источник пищевых ресурсов и полезных ископаемых, но и фактор сбалансированности биосферных процессов, включая постоянство содержания О 2 и СО 2 в атмосфере, постоянство содержания гумуса в почве и температурных условий.
Из сказанного вытекает вывод о необходимости сохранения зеленого царства на нашей планете, особенно лесов. Уменьшение площади лесов и другой растительности, разложение гумуса почв и увеличивающееся сжигание человеком ископаемого топлива приводят к нарушению биосферных процессов.
Материальная база фотосинтеза
Лист — главный орган растения, осуществляющий фотосинтез. Он состоит из нескольких слоев активно фотосинтезирующих кле ток (клеток мезофилла), окруженных защитным слоем (эпидер мисом) и снабженных проводящими элементами (жилками), пред назначенными для переноса веществ в двух противоположных направлениях: по ксилеме жилок вода и питательные вещества поступают в лист, а по флоэме ассимиляты и прочие продукты жизнедеятельности удаляются из листа. Устьица, пронизывающие эпидермис, открываются и закрываются под влиянием измене ний тургора замыкающих клеток. В мезофилле листа расположены пластиды — хлоропласты, содержащие зеленые пигменты (хлоро филл), поэтому окраска листьев зеленая.
Благодаря большой поверхности и определенному размещению листьев в пространстве растение может использовать как прямой, так и рассеянный свет. Для эффективного улавливания света боль шое значение имеет архитектоника растения, под которой пони мают пространственное расположение органов. Для теневыносли вых древесных растений характерна листовая мозаика, когда лис тья отдельных ветвей располагаются в одной плоскости, не зате няя друг друга. Фотосинтез связан со специфическими органеллами клеток — хлоропластами. Суммарная поверхность хлоропластов может пре вышать площадь листьев в десятки и сотни раз. В клетках столбча той паренхимы находится 30 — 40 хлоропластов, в губчатой — около 20. Хлоропласты высших растений имеют форму двояковыпуклой линзы (диска), которая наиболее удобна для поглощения солнеч ных лучей (рис. 1). С помощью электронного микроскопа удалось установить ультраструктуру хлоропласта.

Рисунок 1 – Строение хлоропласта:
1 — оболочка хлоропласта; 2 — грана; 3 — крахмальное зерно; 4 — строма; 5 — пластоглобула
Снаружи хлоропласты окружены двойной мембраной, состоя щей главным образом из белков и липидов. Внутреннюю среду пред ставляет сильно оводненный белковый матрикс, или строма, ко торую пронизывают мембраны — ламеллы. Ламеллы, соединенные друг с другом, образуют тилакоиды. Плотно прилегая друг к другу, тилакоиды формируют граны.
Пигменты, участвующие в поглощении энергии света, находятся в мембранах тилакоидов. Именно здесь световая энергия преобразуется в химическую. Ферменты, которые катализируют многочисленные реакции темновой фазы фотосинтеза, а также биосинтез различных веществ, в том числе белков, липоидов, крахмала, находятся в строме.
В тилакоидах хлоропластов содержатся зеленые пигменты — хлорофилл а и хлорофилл b , а также желтые — каротины и ксан тофиллы (каротиноиды). Все пластидные пигменты нерастворимы в воде, но растворяются в органических растворителях. Раствор хлорофилла а в этиловом спирте имеет сине-зеленый цвет, ра створ хлорофилла b — желто-зеленый.
Хлорофилл — сложный эфир дикарбоновой кислоты хлорофиллина с метиловым спиртом (СН 3 ОН) и фитолом (С 2 оНз 9 ОН):

Хлорофиллиновая кислота представляет собой порфириновое ядро, состоящее из четырех пиррольных азотсодержащих колец и атома Mg . Хлорофилл b отличается от хлорофилла а лишь тем, что у второго пиррольного кольца вместо метильной группы СН 3 имеется альдегидная группа О = С—Н, что отражается на поло жении максимумов поглощения света. Содержание хлорофилла а в 2 — 3 раза больше, чем хлорофилла Ь. У теневых листьев хлоро филла b больше, чем у световых. Общее количество хлорофилла составляет около 1 % сухой массы листа. Оно изменяется в связи с возрастом листа и деревьев. В зеленых листьях пигменты желтого и оранжевого цвета маскируются хлорофиллом. С его более ранним разрушением осенью или при неблагоприятных условиях связано пожелтение листьев.
Фотосинтетические пигменты избирательно поглощают свет в области видимой части солнечного спектра (длина волны 400 — 700 нм), эту область называют фотосинтетически активной радиацией (ФАР). Листья поглощают в среднем около 60 — 70 % ФАР, а остальную радиацию отражают и пропускают. Кора ветвей и стволов большинства древесных растений, за исключением белой бересты берез, оранжево-красной пробки сосен, поглощает до 80 — 90% и отражает 10 — 20% энергии падающей радиации. Хлорофиллы наиболее интенсивно поглощают красные и сине-фиолетовые лучи спектра, слабо поглощают оранжевый и желтый цвет. Ультрафиолетовые и зеленые лучи практически не поглощаются, поэтому мы видим листья зелеными. Каротиноиды поглощают синие и фиолетовые лучи. Поглощенная этими пигментами энергия передается основному пигменту — хлорофиллу. Кроме того, каротиноиды защищают молекулы хлорофиллов от необратимого фотоокисления.
Химизм и энергетика фотосинтеза
Фотосинтез – это процесс преобразования света в химическую энергию органических соединений, синтезируемых из СО 2 и Н 2 О.
Фотосинтез протекает в две фазы: световую и темновую .

Процесс фотосинтеза начинается с момента освещения хлоропласта видимым светом. При поглощении молекулой хлорофилла кванта света один из ее электронов переходит в "возбужденное" состояние и поднимается на более высокий энергетический уровень.
Одновременно под действием света происходит фотолиз воды с образованием ионов Н + и ОН - . Возбужденный электрон присоединяется к иону водорода (Н + ), восстанавливая его до атома (Н). Далее атомы водорода соединяются с никотинамидадениндинуклеотидфосфатом (НАДФ) и восстанавливают его до НАДФ • Н 2 .
Ионы гидроксила, оставшись без противоионов Н + , отдают свои электроны и превращаются в свободные радикалы ОН, которые, взаимодействуя друг с другом, образуют воду и свободный кислород: 4ОН → 2Н 2 О+СО 2 . Электроны гидроксильных групп возвращаются в молекулу хлорофилла на место возбужденных. В процессе переходов протоны и электроны накапливаются по разные стороны мембраны грана хлоропласта (протоны на внутренней, а электроны на наружной поверхности) и создают разность потенциалов. Когда разность потенциалов достигает критического уровня, протоны проходят по специальным каналам мембран, в которых находятся ферменты, синтезирующие АТФ. Энергия протонов и электронов используется ферментами для присоединения остатка фосфорной кислоты к АМФ или АДФ.
Таким образом, в световую фазу фотосинтеза, которая протекает в гранах хлоропластов только на свету, происходят следующие процессы:
1) фотолиз воды с выделением кислорода;
2) восстановление НАДФ • Н 2 ;
3) синтез АТФ.
В темновую фазу фотосинтеза накопленная в световую фазу энергия используется для синтеза моносахаридов из диоксида углерода (поступает из воздуха через устьица) и водорода (отсоединяется от НАДФ • Н 2 ) путем сложных ферментативных реакций. В итоге получается:
В дальнейшем могут образовываться ди-, полисахариды и другие органические соединения (аминокислоты, жирные кислоты и др.). Этот процесс не требует прямого участия света, поэтому его называют темновой фазой фотосинтеза. Он протекает в строме хлоропластов как на свету, так и в темноте.
Коэффициент полезного действия фотосинтеза достигает 60 %.
Фотосинтез – это главный процесс, протекающий в биосфере. Энергия Солнца аккумулируется в химических связях органических соединений, которые идут на питание всех гетеротрофов. При этом атмосфера обогащается кислородом и очищается от избытка диоксида углерода.
Из кислорода образовался озоновый экран, защищающий живые организмы от жесткой коротковолновой (до 290 нм) ультрафиолетовой радиации, которая оказывает губительное воздействие на все живое.
Наземные растения ежегодно извлекают из атмосферы 20 млрд т углерода в форме СО 2 (1300 кг/га), а все растительные сообщества, включая морские водоросли, — около 150 млрд т. При этом ежегодно расходуется около 3% диоксида углерода атмосферы и 0,3% С0 2 морской воды.
Влияние внутренних и внешних факторов на фотосинтез
К ведущим факторам внутренней регуляции фотосинтеза относятся скорость оттока ассимилятов из хлоропластов и концентрация хлорофилла в листьях. Интенсивность фотосинтеза листа в значительной степени зависит от запроса на ассимиляты со стороны потребляющих органов (акцепторов). Мощными потребителями являются меристематические ткани, в которых идет активный биосинтез. Растущее дерево активно потребляет ассимиляты на рост ствола, ветвей, корневых систем, на формирование плодов, семян, на биосинтез живицы у хвойных. Любое ускорение оттока ассимилятов усиливает интенсивность фотосинтеза. Неблагоприятные лесорастительные условия задерживают отток ассимилятов из хлоропластов и за счет этого снижают активность фотосинтеза. К внешним факторам, влияющим на фотосинтез, относят свет, концентрацию диоксида углерода, температуру и водоснабжение растений.
Свет. По отношению к свету все растения делят на две группы: светолюбивые и теневыносливые. Они различаются как по анатомическому строению листьев, так и по физиологическим признакам. Светолюбивые травянистые растения не выносят затенения и растут на открытых местах. К ним относятся все сельскохозяйственные культуры, растения лугов, степей, пустынь, солончаков. Среди древесных пород светолюбивы лиственница, сосна, ясень, осина, береза, дуб и др. Они растут на открытых местах или в первом верхнем ярусе леса. Светолюбивые деревья отличаются ажурной кроной, быстрым очищением ствола от сучьев, ранним изреживанием древостоя. Теневыносливые древесные растения, такие как ель, пихта, клен, вяз, липа, рябина и другие, хорошо переносят затенение и встречаются как в верхнем ярусе, так и во втором. Высокой теневыносливостью отличаются многие кустарники (лещина, крушина, бересклет, бузина и др.), а также лесные травы и мхи. Теневыносливые древесные растения отличаются густой и плотной кроной с большой протяженностью по высоте ствола. Для них характерно медленное очищение от сучьев. Листья светолюбивых растений имеют более толстую листовую пластинку, хорошо развитую столбчатую паренхиму, большое количество устьиц и проводящих пучков. Содержание пигментов меньше, чем у теневыносливых. Более высокое содержание пигментов и относительной доли хлорофилла б и каротиноидов у теневыносливых растений обеспечивает эффективный фотосинтез в условиях низкой интенсивности света и рассеянной радиации. Светолюбивые и теневыносливые растения имеют разный ход световых кривых. У теневыносливых световое насыщение наступает при более слабом освещении и, кроме того, световой компенсационный пункт (СКП), т.е. та освещенность, при которой фотосинтез и дыхание уравновешивают друг друга, наступает раньше. Это связано с тем, что теневыносливые растения имеют более высокую интенсивность фотосинтеза при низкой освещенности и невысокую интенсивность дыхания. СКП широко используется при оценке качества и состояния естественного возобновления в лесу, поскольку подрост начинает страдать и отмирать под пологом леса при освещенности, равной СКП или ниже его. Периодичность рубок ухода за лесом также определяется величиной СКП.
Концентрация диоксида углерода. СО 2 — основной субстрат фотосинтеза. Его содержание в атмосфере в значительной степени определяет интенсивность процесса. Концентрация СО 2 в атмосфере составляет 0,03 %. При этой концентрации интенсивность; фотосинтеза составляет лишь 50 % от максимальной величины, которая достигается при содержании 0,3 % СО 2 . Поэтому в условиях закрытого грунта весьма эффективны подкормки растений СО 2 .
Температура. Влияние температуры на фотосинтез зависит от интенсивности освещения. При низкой освещенности фотосинтез практически не зависит от температуры, так как лимитируется световыми реакциями. При высокой освещенности скорость фотосинтеза определяется протеканием темновых реакций и в этом случае влияние температуры проявляется отчетливо. Для большинства растений оптимальная температура составляет 20 — 30 °С. Температурный минимум для хвойных колеблется между -2 и -7 0 С.
Снабжение водой. На интенсивность фотосинтеза благоприятно влияет небольшой водный дефицит (до 5 %) в клетках листьев. Однако при недостаточном водоснабжении интенсивность фотосинтеза заметно снижается. Это связано с закрыванием устьиц, в результате чего замедляется доставка СО 2 в лист, а также с задержкой ферментативных реакций фотосинтеза и оттока образовавшихся продуктов фотосинтеза из листа. В природных условиях недостаточное снабжение листьев водой может быть вызвано как почвенной засухой, так и избытком воды в почве. При затоплении в почве создаются полуанаэробные или анаэробные условия, при которых корни растений не могут поглощать воду в достаточных количествах и в листьях возникает водный дефицит.
Суточный ход фотосинтеза. В естественных условиях при сложном взаимодействии внешних и внутренних факторов суточный ритм фотосинтеза неустойчив и подвержен значительным изменениям. При умеренной дневной температуре и достаточном водоснабжении фотосинтез начинается с восходом солнца, достигает максимума днем, постепенно снижаясь к вечеру, и прекращается с заходом солнца. При повышенной температуре и низкой влажности максимум фотосинтеза сдвигается на ранние часы. Часто при высокой напряженности метеорологических факторов наблюдается значительное снижение фотосинтеза в полуденные часы, связанное с закрыванием устьиц и уменьшением оттока ассимилятов.
Фотосинтез и урожай
Биологическим урожаем называется количество органического вещества, накопленное на единицу площади растительного сообщества. Человек использует далеко не весь биологический урожай. Обычно ту часть чистой продукции, которую человек использует па свои нужды, называют хозяйственным урожаем. Урожай растений является функцией фотосинтеза. Сухая масса растений на 95 % состоит из органических соединений, созданных в процессе фотосинтеза, от которого зависит не только количество, но и качество урожая. Связь между фотосинтезом и урожаем довольно сложна, так как общее количество накопленного за определенный период времени органического вещества определяется не только фотосинтезом, но и противоположно направленным процессом дыхания. При этом следует учитывать и опад различных частей: листьев, ветвей, корней, корневых волосков и других органов, что особенно заметно сказывается на балансе органического вещества у древесных растений. Лесохозяйственная часть урожая — это, прежде всего масса сформировавшейся древесины, ее количество на единице площади и высокие физико-технические свойства. Лесохозяйственный урожай формируется главным образом за счет камбиальной деятельности ствола и выражается в приросте древесины. Специальные расчеты показали, что доля древесины с ухудшением лесорастительных условий и с продвижением с юга на север уменьшается. Для увеличения продуктивности лесов и улучшения их качественного состава в лесном хозяйстве широко используются рубки ухода. Регулируя световой режим в лесу, эти рубки оптимизируют процесс фотосинтеза оставшихся деревьев, причем не только в верхней, но и в нижней части кроны, и тем самым повышают размеры активно фотосинтезирующей поверхности этих деревьев. Наряду с рубками ухода существуют и другие мероприятия, направленные на регулирование фотосинтетической активности древесных растений. К ним относятся такие приемы, как орошение в засушливых областях, мелиорация, внесение минеральных удобрений и др.

Зеленый лист – источник жизни на нашей планете. Если бы не зеленые растения, на Земле не было бы ни животных, ни людей. Так или иначе, растения служат источником пищи для всего животного мира.
Человек использует энергию не только солнечных лучей, падающих на землю сейчас, но и тех, что падали на нее десятки и сотни миллионов лет назад. Ведь и уголь, и нефть, и торф – химически измененные остатки растений и животных, живших в те далекие времена.
В последние десятилетия к проблеме фотосинтеза приковано внимание ведущих специалистов ряда отраслей естествознания, ее различные аспекты всесторонне и глубоко исследуются во многих лабораториях мира. Интерес определяется прежде всего тем, что фотосинтез составляет основу энергообмена всей биосферы.
Интенсивность фотосинтеза зависит от многих факторов. Интенсивность света, необходимая для наибольшей эффективности фотосинтеза, у различных растений различна. У теневыносливых растений максимум активности фотосинтеза достигается примерно при половине полного солнечного освещения, а у светолюбивых растений – почти при полном солнечном освещении.
У многих теневыносливых растений не развивается палисадная (столбчатая) паренхима в листьях, и имеется только губчатая (ландыш, копытень). Кроме того, эти растения имеют более крупные листья и более крупные хлоропласты.
Также на интенсивность фотосинтеза влияет температура окружающей среды. Наибольшая интенсивность фотосинтеза наблюдается при температуре 20–28 °С. При дальнейшем повышении температуры интенсивность фотосинтеза падает, а интенсивность дыхания возрастает. Когда интенсивности фотосинтеза и дыхания совпадают, говорят о компенсационном пункте.
Компенсационный пункт изменяется в зависимости от интенсивности света, повышения и понижения температуры. Например, у холодостойких бурых морских водорослей он соответствует температуре около 10 °С. Температура влияет, в первую очередь, на хлоропласты, у которых в зависимости от температуры изменяется структура, что хорошо видно в электронном микроскопе.
Очень большое значение для фотосинтеза имеет содержание углекислого газа в окружающем растение воздухе. Средняя концентрация углекислоты в воздухе составляет 0,03% (по объему). Понижение содержания углекислого газа неблагоприятно влияет на урожай, а его повышение, например до 0,04% может повысить урожай почти в 2 раза. Более значительное повышение концентрации вредно для многих растений: например, при содержании углекислого газа около 0,1% растения томатов заболевают, у них начинают скручиваться листья. В оранжереях и теплицах можно повысить содержание углекислого газа, выпуская его из специальных баллонов или давая испаряться сухой углекислоте.
Свет разных длин волн также по-разному влияет на интенсивность фотосинтеза. Впервые интенсивность фотосинтеза в различных лучах спектра исследовал физик В. Добени, показавший в 1836г., что скорость фотосинтеза в зеленом листе зависит от характера лучей. Методические погрешности при проведении эксперимента привели его к неправильным выводам. Ученый поместил отрезок побега элодеи в пробирку с водой срезом вверх, освещал пробирку, пропуская солнечный свет через цветные стекла или окрашенные растворы, и учитывал интенсивность фотосинтеза по количеству пузырьков кислорода, отрывающихся с поверхности среза за единицу времени. Добени пришел к выводу, что интенсивность фотосинтеза пропорциональна яркости света, а наиболее яркими лучами в то время считались желтые. Этой же точки зрения придерживался и Джон Дрепер (1811–1882), который изучал интенсивность фотосинтеза в различных лучах спектра, испускаемых спектроскопом.
Роль хлорофилла в процессе фотосинтеза доказал выдающийся российский ботаник и физиолог растений К.А. Тимирязев. Проведя в 1871–1875 гг. серию опытов, он установил, что зеленые растения наиболее интенсивно поглощают лучи красной и синей части солнечного спектра, а не желтые, как это считалось до него. Поглощая красную и синюю часть спектра, хлорофилл отражает зеленые лучи, из-за чего и кажется зеленым.
На основании этих данных немецкий физиолог растений Теодор Вильгельм Энгельман в 1883 г. разработал бактериальный метод изучения ассимиляции углекислого газа растениями.

Он предположил, что если поместить в каплю воды клетку зеленого растения вместе с аэробными бактериями и осветить их разноокрашенными лучами, то бактерии должны концентрироваться у тех участков клетки, в которых сильнее всего разлагается углекислый газ и выделяется кислород. Чтобы проверить это, Энгельман несколько усовершенствовал световой микроскоп, укрепив над зеркальцем призму, которая разлагала солнечный свет на отдельные составляющие спектра. В качестве зеленого растения Энгельман использовал зеленую водоросль спирогиру, крупные клетки которой содержат длинные спиральные хроматофоры.
Поместив в каплю воды на предметном стекле кусочек водоросли, Энгельман внес туда же немного аэробных бактерий, после чего, рассмотрел препарат под микроскопом. Оказалось, что в отсутствии призмы приготовленный препарат освещался ровным белым светом, и бактерии равномерно распределялись вдоль всего участка водоросли. В присутствии призмы отраженный от зеркальца луч света преломлялся, освещая участок водоросли под микроскопом светом с разной длиной волны. Спустя несколько минут, бактерии сконцентрировались на тех участках, которые были освещены красным и синим светом. На основании этого Энгельман сделал вывод о том, что разложение углекислого газа, (а, значит, и выделение кислорода) у зеленых растений наблюдается в дополнительных к основной окраске (т.е. зеленой) лучах – красных и синих.

Данные, полученные на современном оборудовании, полностью подтверждают результаты, полученные Энгельманом более 120 лет назад.
Поглощенная хлорофиллом световая энергия принимает участие в реакциях первого и второго этапов фотосинтеза; реакции третьего этапа являются темновыми, т.е. происходит без участия света. Измерения показали, что процесс восстановления одной молекулы кислорода требует минимум восьми квантов световой энергии. Таким образом, максимальный квантовый выход фотосинтеза, т.е. число молекул кислорода, соответствующее одному кванту поглощенной растением световой энергии, составляет 1/8, или 12,5%.
Р.Эмерсон с сотрудниками определили квантовый выход фотосинтеза при освещении растений монохроматическим светом различной длины волны. При этом установлено, что выход остается постоянным на уровне 12% в большей части видимого спектра, но резко снижается вблизи дальней красной области. Это снижение у зеленых растений начинается при длине волны 680 нм. При длине больше 660 нм свет поглощает только хлорофилл a; хлорофилл b имеет максимум поглощения света при 650 нм, а при 680 нм практически свет не поглощает. При длине волны больше, чем 680 нм, квантовый выход фотосинтеза может быть доведен до максимальной величины 12% при условии, что растение одновременно будет освещаться также светом с длиной волны 650 нм. Иначе говоря, если свет, поглощаемым хлорофиллом а дополняется светом, поглощаемый хлорофиллом b, то квантовый выход фотосинтеза достигает нормальной величины.
Усиление интенсивности фотосинтеза при одновременном освещении растения двумя лучами монохроматического света различной длины волны по сравнению с его интенсивностью, наблюдаемой при раздельном освещении этими же лучами, получило название эффекта Эмерсона. Опыты с различными комбинациями дальнего красного света и света с более короткой длиной волны над зелеными, красными, синезелеными и бурыми водорослями показали, что наибольшее усиление фотосинтеза наблюдается в том случае, если второй луч с более короткой длиной волны поглощается вспомогательнымих пигментами.

У зеленых растений такими вспомогательными пигментами являются каротиноиды и хлорофилл b, у красных водорослей – каротиноиды и фикоэритрин, у синезеленых – каротиноиды и фикоцианин, у бурых водорослей – каротиноиды и фукоксантин.
Дальнейшее изучение процесса фотосинтеза привело к заключению, что вспомогательные пигменты передают от 80 до 100% поглощенной ими световой энергии хлорофиллу а. Таким образом, хлорофилл а аккумулирует световую энергию, поглощаемую растительной клеткой, и затем использует ее в фотохимических реакциях фотосинтеза.
Позже было обнаружено, что хлорофилл а присутствует в живой клетке в виде форм с различными спектрами поглощения и различными фотохимическими функциями. Одна форма хлорофилла а, максимум поглощения у которой соответствует длине волны 700 нм, принадлежит к пигментной системе, получившей название фотосистема I, вторая форма хлорофилла а с максимумом поглощения 680 нм, принадлежит к фотосистеме II.
Итак, в растениях была открыта фотоактивная пигментная система, особенно сильно поглощающая свет в красной области спектра. Она начинает действовать уже при ничтожной освещенности. Кроме того, известна и другая регуляторная система, которая избирательно поглощает и использует для фотосинтеза синий цвет. Эта система работает при достаточно сильном свете.
Установлено также, что фотосинтетический аппарат одних растений в значительной степени использует для фотосинтеза красный свет, других – синий.
Для определения интенсивности фотосинтеза водных растений можно использовать метод подсчета пузырьков кислорода. На свету в листьях происходит процесс фотосинтеза, продуктом которого является кислород, накапливающийся в межклетниках. При срезании стебля избыток газа начинает выделяться с поверхности среза в виде непрерывного тока пузырьков, быстрота образования которых зависит от интенсивности фотосинтеза. Данный метод не отличается большой точностью, но зато прост и дает наглядное представление о зависимости процесса фотосинтеза от внешних условий.
Опыт 1. Зависимость продуктивности фотосинтеза от интенсивности света
Материалы и оборудование: элодея; водные растворы NaHCO3, (NH4)2CO3 или минеральная вода; отстоявшаяся водопроводная вода; стеклянная палочка; нитки; ножницы; электролампа мощностью 200 Вт; часы; термометр.
1. Для опыта отбирали здоровые побеги элодеи длиной около 8 см интенсивного зеленого цвета с неповрежденной верхушкой. Их подрезали под водой, привязывали ниткой к стеклянной палочке и опускали верхушкой вниз в стакан с водой комнатной температуры (температура воды должна оставаться постоянной).
2. Для опыта брали отстоявшуюся водопроводную воду, обогащенную СО2 добавлением NaHCO3 или (NH4)2CO3, или минеральную воду, и выставляли стакан с водным растением на яркий свет. Наблюдали за появлением пузырьков воздуха из среза растения.
3. Когда ток пузырьков становился равномерным, подсчитывали количество пузырьков, выделившихся за 1 мин. Подсчет проводили 3 раза с перерывом в 1 мин, данные записывали в таблицу, определяли средний результат.
4. Стакан с растением удаляли от источника света на 50–60 см и повторяли действия, указанные в п. 3.
5. Результаты опытов сравнивали и делали вывод о различной интенсивности фотосинтеза на ярком и слабом свету.
Результаты опытов представлены в таблице 1.
Вывод: при использованных интенсивностях света интенсивность фотосинтеза увеличивается с ростом интенсивности света, т.е. чем больше света, тем лучше идет фотосинтез.
Таблица 1. Зависимость фотосинтеза от интенсивности света
Фотосинтез зависит от множества факторов, которые определяют его интенсивность и продуктивность. На интенсивность процесса фотосинтеза влияют следующие условия: освещение, температура, содержание СО2 в воздухе, снабжение водой, кислородом, минеральное питание, содержание хлорофилла, возраст ассимилирующего органа, степень открытости устьиц. В естественной обстановке все факторы взаимодействуют друг с другом, т.е. действие одного фактора зависит от напряженности всех остальных.
В ясный солнечный день на 1 дм 2 листовой поверхности за 1 ч падает около 30 168 кДж. Из этого количества поглощается примерно 75%, или 22 626 кДж, 25% падающей энергии проходят через лист и отражаются от него (рис.11).
На фотосинтез расходуется 1,5-2% поглощенной энергии.
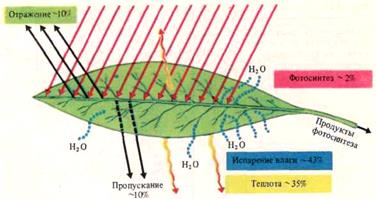
Рис. 11. Примерный энергетический баланс листа растения
Суточные ритмы фотосинтеза можно представить в виде кривой (рис.12):
Рис. 12. Суточный ход интенсивности фотосинтеза
С восходом солнца интенсивность фотосинтеза возрастает вместе с освященностью, достигая максимумов в 9-10 ч. Дальнейший характер процесса определяется степенью оводненности листьев, температурой воздуха, интенсивностью солнечного света, содержанием СО2 в воздухе и т.д.
В нормальных для фотосинтеза условиях процесс протекает неравномерно в течение дня. Утром, к 9-10 ч, наблюдается наиболее интенсивный фотосинтез, затем к 12 часам по мере накопления крахмала и уменьшения количества воды в листьях интенсивность снижается и опять возрастает ко второй половине дня – к 17-18 ч.
Свет. Если расположить факторы, от которых зависит фотосинтез, по их значению, на первое место следует поставить свет. В целом зависимость фотосинтеза от интенсивности освещения может быть выражена логарифмической кривой (рис.13).
Первоначально увеличение интенсивности освещения приводит к пропорциональному усилению фотосинтеза (зона максимального эффекта). При дальнейшем увеличении интенсивности света фотосинтез продолжает расти, но медленнее (зона ослабленного эффекта). И, наконец, интенсивность света растет, а фотосинтез не изменяется (зона отсутствия эффекта).
Наклон кривых, выражающих зависимость интенсивности фотосинтеза от освещенности, различен для разных растений. Есть растения, у которых фотосинтез возрастает вплоть до освещения их прямыми лучами.

Рис.13. Зависимость интенсивности фотосинтеза от освещения у кукурузы
Вместе с тем для многих растений увеличение интенсивности освещения свыше 50% от прямого солнечного света оказывается излишним. Это связано с тем, что конечный выход продуктов фотосинтеза зависит от скорости не столько световых, сколько темновых реакций.
Температура. Общая зависимость фотосинтеза от температуры выражается одновершинной кривой (рис.14). Кривая имеет три основные (кардинальные) температурные точки: минимальную, при которой начинается фотосинтез, оптимальную и максимальную. Нижняя температурная граница фотосинтеза у растений северных широт находится в пределах -15°С (сосна, ель), а у тропических растений – в зоне низких положительных температур – 4-8°С. У растений умеренного пояса в интервале 20-25°С достигается максимальная интенсивность фотосинтеза, а дальнейшее повышение температуры до 40°С приводит к быстрому ингибированию процесса.
Влияние на интенсивность фотосинтеза света, концентрации СО2 и температуры осуществляется в сложном взаимодействии. Особенно тесно взаимосвязаны свет, действующий на скорость фотохимических реакций, и температура, контролирующая скорость энзиматических реакций (рис.15).
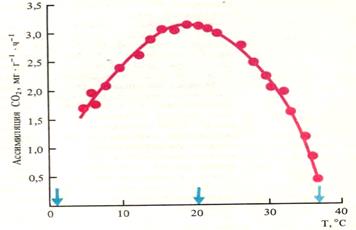
Рис. 14. Зависимость интенсивности фотосинтеза ели от температуры
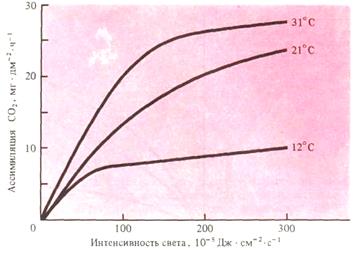 |
Рис.15. Взаимовлияние интенсивности света и температуры на скорость фотосинтеза
СО2. Основным субстратом фотосинтеза является СО2: его содержание определяет интенсивность процесса. Концентрация СО2 в атмосфере составляет 0,03%. Процесс фотосинтеза осуществляется при содержании СО2 не менее 0,008%. Повышение содержания СО2 до 1,5% вызывает прямо пропорциональное возрастание интенсивности процесса фотосинтеза. При повышении содержания СО2 свыше 1,5% фотосинтез продолжает расти, но уже значительно медленнее. При увеличении содержания СО2 до 15-20% процесс фотосинтеза выходит на плато (отсутствие эффекта). При содержании СО2 выше 70% наступает депрессия фотосинтеза (рис. 16).
Рис. 16. Зависимость интенсивности фотосинтеза от концентрации СО2
Взаимовлияние концентрации СО2 и интенсивности света на скорость фотосинтеза показано на рисунке 17.
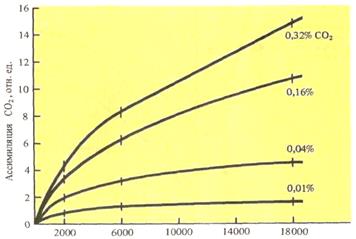 |
Рис.17. Взаимовлияние концентрации СО2 и интенсивности света на скорость фотосинтеза
Водный фактов имеет первоочередное значение для нормального осуществления фотосинтетических функций растений. Значение воды в фотосинтезе обусловливается ее участием в фотохимических и энзиматических реакциях. От степени оводненности тканей растения зависит поглощение энергии солнечной радиации, поступление и ассимиляция углекислого газа, направленность ферментативных реакций, интенсивность транспирации и т.д. Водный дефицит приводит к снижению интенсивности фотосинтеза, которое связано с закрыванием устьиц, что уменьшает диффузию СО2 в лист, к ингибированию циклического и нециклического транспорта электронов и фотофосфорилирования, к снижению активности ферментов, участвующих в темновой фазе фотосинтеза.
Минеральное питание. Для нормального функционирования фотосинтетического аппарата растение должно быть обеспечено всем комплексом макро- и микроэлементов. Два основных процесса питания растительного организма - воздушный и корневой – тесно взаимосвязаны. Многие соединения, функционирующие как переносчики, содержат такие элементы как железо (цитохромы, ферредоксин), медь (пластоцианин). Хлорофилл содержит магний. При недостатке этих элементов интенсивность фотосинтеза понижается.
На интенсивность процесса фотосинтеза влияют и внутренние факторы: содержание хлорофилла, возраст ассимилирующего органа, степень открытости устьиц.
Содержание хлорофилла. Интенсивность фотосинтеза возрастает с увеличением содержания хлорофилла. Содержание хлорофилла и интенсивность освещения можно рассматривать как единый фактор, определяющий поглощение света. При высокой интенсивности света даже сравнительно небольшое количество хлорофилла может поглощать его достаточно, для того чтобы было достигнуто световое насыщение.
Отток ассимилятов. Депрессия фотосинтеза часто связана с нарушением оттока ассимилятов.При нарушении оттока ассимилятов, продукты фотосинтеза (конечные или промежуточные) переполняют клетки и препятствуют их новообразованию, а также они блокируют некоторые ферменты фотосинтетического цикла.
Возраст листа. По мере роста листьев интенсивность фотосинтеза повышается. Наибольшая интенсивность характерна для вполне сформировавшихся листьев. По мере дальнейшего увеличения возраста листьев интенсивность фотосинтеза падает. На интенсивность фотосинтеза оказывает влияние возраст всего растения. У большинства однолетних растений интенсивность фотосинтеза возрастает в процессе онтогенеза и достигает максимума в фазу бутонизации и цветения. После цветения интенсивность фотосинтеза в листьях снижается.
Степень открытости устьиц. В том случае, если содержание воды поддерживается в листе на достаточно высоком уровне, увеличение ширины устьичных щелей, как правило, сопровождается увеличением интенсивности фотосинтеза. При полном закрытии устьиц фотосинтез протекает, но в очень слабой степени.
Влияние внешних и внутренних факторов на интенсивность фотосинтеза можно представить в виде схемы (рис. 18).
Рис.18. Влияние внешних и внутренних факторов на интенсивность фотосинтеза
Фотосинтез зависит от множества факторов, которые определяют его интенсивность и продуктивность. На интенсивность процесса фотосинтеза влияют следующие условия: освещение, температура, содержание СО2 в воздухе, снабжение водой, кислородом, минеральное питание, содержание хлорофилла, возраст ассимилирующего органа, степень открытости устьиц. В естественной обстановке все факторы взаимодействуют друг с другом, т.е. действие одного фактора зависит от напряженности всех остальных.
В ясный солнечный день на 1 дм 2 листовой поверхности за 1 ч падает около 30 168 кДж. Из этого количества поглощается примерно 75%, или 22 626 кДж, 25% падающей энергии проходят через лист и отражаются от него (рис.11).
На фотосинтез расходуется 1,5-2% поглощенной энергии.
Рис. 11. Примерный энергетический баланс листа растения
Суточные ритмы фотосинтеза можно представить в виде кривой (рис.12):
Рис. 12. Суточный ход интенсивности фотосинтеза
С восходом солнца интенсивность фотосинтеза возрастает вместе с освященностью, достигая максимумов в 9-10 ч. Дальнейший характер процесса определяется степенью оводненности листьев, температурой воздуха, интенсивностью солнечного света, содержанием СО2 в воздухе и т.д.
В нормальных для фотосинтеза условиях процесс протекает неравномерно в течение дня. Утром, к 9-10 ч, наблюдается наиболее интенсивный фотосинтез, затем к 12 часам по мере накопления крахмала и уменьшения количества воды в листьях интенсивность снижается и опять возрастает ко второй половине дня – к 17-18 ч.
Свет. Если расположить факторы, от которых зависит фотосинтез, по их значению, на первое место следует поставить свет. В целом зависимость фотосинтеза от интенсивности освещения может быть выражена логарифмической кривой (рис.13).
Первоначально увеличение интенсивности освещения приводит к пропорциональному усилению фотосинтеза (зона максимального эффекта). При дальнейшем увеличении интенсивности света фотосинтез продолжает расти, но медленнее (зона ослабленного эффекта). И, наконец, интенсивность света растет, а фотосинтез не изменяется (зона отсутствия эффекта).
Наклон кривых, выражающих зависимость интенсивности фотосинтеза от освещенности, различен для разных растений. Есть растения, у которых фотосинтез возрастает вплоть до освещения их прямыми лучами.
Рис.13. Зависимость интенсивности фотосинтеза от освещения у кукурузы
Вместе с тем для многих растений увеличение интенсивности освещения свыше 50% от прямого солнечного света оказывается излишним. Это связано с тем, что конечный выход продуктов фотосинтеза зависит от скорости не столько световых, сколько темновых реакций.
Температура. Общая зависимость фотосинтеза от температуры выражается одновершинной кривой (рис.14). Кривая имеет три основные (кардинальные) температурные точки: минимальную, при которой начинается фотосинтез, оптимальную и максимальную. Нижняя температурная граница фотосинтеза у растений северных широт находится в пределах -15°С (сосна, ель), а у тропических растений – в зоне низких положительных температур – 4-8°С. У растений умеренного пояса в интервале 20-25°С достигается максимальная интенсивность фотосинтеза, а дальнейшее повышение температуры до 40°С приводит к быстрому ингибированию процесса.
Влияние на интенсивность фотосинтеза света, концентрации СО2 и температуры осуществляется в сложном взаимодействии. Особенно тесно взаимосвязаны свет, действующий на скорость фотохимических реакций, и температура, контролирующая скорость энзиматических реакций (рис.15).
Рис. 14. Зависимость интенсивности фотосинтеза ели от температуры
| |
Рис.15. Взаимовлияние интенсивности света и температуры на скорость фотосинтеза
СО2. Основным субстратом фотосинтеза является СО2: его содержание определяет интенсивность процесса. Концентрация СО2 в атмосфере составляет 0,03%. Процесс фотосинтеза осуществляется при содержании СО2 не менее 0,008%. Повышение содержания СО2 до 1,5% вызывает прямо пропорциональное возрастание интенсивности процесса фотосинтеза. При повышении содержания СО2 свыше 1,5% фотосинтез продолжает расти, но уже значительно медленнее. При увеличении содержания СО2 до 15-20% процесс фотосинтеза выходит на плато (отсутствие эффекта). При содержании СО2 выше 70% наступает депрессия фотосинтеза (рис. 16).
Рис. 16. Зависимость интенсивности фотосинтеза от концентрации СО2
Взаимовлияние концентрации СО2 и интенсивности света на скорость фотосинтеза показано на рисунке 17.
| |
Рис.17. Взаимовлияние концентрации СО2 и интенсивности света на скорость фотосинтеза
Водный фактов имеет первоочередное значение для нормального осуществления фотосинтетических функций растений. Значение воды в фотосинтезе обусловливается ее участием в фотохимических и энзиматических реакциях. От степени оводненности тканей растения зависит поглощение энергии солнечной радиации, поступление и ассимиляция углекислого газа, направленность ферментативных реакций, интенсивность транспирации и т.д. Водный дефицит приводит к снижению интенсивности фотосинтеза, которое связано с закрыванием устьиц, что уменьшает диффузию СО2 в лист, к ингибированию циклического и нециклического транспорта электронов и фотофосфорилирования, к снижению активности ферментов, участвующих в темновой фазе фотосинтеза.
Минеральное питание. Для нормального функционирования фотосинтетического аппарата растение должно быть обеспечено всем комплексом макро- и микроэлементов. Два основных процесса питания растительного организма - воздушный и корневой – тесно взаимосвязаны. Многие соединения, функционирующие как переносчики, содержат такие элементы как железо (цитохромы, ферредоксин), медь (пластоцианин). Хлорофилл содержит магний. При недостатке этих элементов интенсивность фотосинтеза понижается.
На интенсивность процесса фотосинтеза влияют и внутренние факторы: содержание хлорофилла, возраст ассимилирующего органа, степень открытости устьиц.
Содержание хлорофилла. Интенсивность фотосинтеза возрастает с увеличением содержания хлорофилла. Содержание хлорофилла и интенсивность освещения можно рассматривать как единый фактор, определяющий поглощение света. При высокой интенсивности света даже сравнительно небольшое количество хлорофилла может поглощать его достаточно, для того чтобы было достигнуто световое насыщение.
Отток ассимилятов. Депрессия фотосинтеза часто связана с нарушением оттока ассимилятов.При нарушении оттока ассимилятов, продукты фотосинтеза (конечные или промежуточные) переполняют клетки и препятствуют их новообразованию, а также они блокируют некоторые ферменты фотосинтетического цикла.
Возраст листа. По мере роста листьев интенсивность фотосинтеза повышается. Наибольшая интенсивность характерна для вполне сформировавшихся листьев. По мере дальнейшего увеличения возраста листьев интенсивность фотосинтеза падает. На интенсивность фотосинтеза оказывает влияние возраст всего растения. У большинства однолетних растений интенсивность фотосинтеза возрастает в процессе онтогенеза и достигает максимума в фазу бутонизации и цветения. После цветения интенсивность фотосинтеза в листьях снижается.
Степень открытости устьиц. В том случае, если содержание воды поддерживается в листе на достаточно высоком уровне, увеличение ширины устьичных щелей, как правило, сопровождается увеличением интенсивности фотосинтеза. При полном закрытии устьиц фотосинтез протекает, но в очень слабой степени.
Влияние внешних и внутренних факторов на интенсивность фотосинтеза можно представить в виде схемы (рис. 18).
Рис.18. Влияние внешних и внутренних факторов на интенсивность фотосинтеза

Что нужно повторить для успешного изучения темы? § 23 – учебник для 7 класса; § 54 – учебник для 8 класса.
Факторы, влияющие на фотосинтез. Как вы помните, на физиологические процессы в живых организмах оказывают влияние различные факторы, как внешние, так и внутренние. К внутренним факторам, влияющим на фотосинтез, будут относиться особенности строения хлоропластов, тип хлорофилла, особенности других пигментов и биохимическая конструкция всех ферментов и молекул-переносчиков, задействованных в этом процессе. К внешним факторам будут относиться условия окружающей среды, которые необходимы для фотосинтеза, либо те, что могут отрицательно повлиять на растение, даже не участвуя в фотосинтезе непосредственно.
Лимитирующие факторы фотосинтеза – это те условия, при увеличении (улучшении) которых скорость и эффективность процесса фотосинтеза будут возрастать. Какие же внешние факторы необходимы для осуществления процесса фотосинтеза? Прежде всего, это свет, вода и углекислый газ. Так как почти все биохимические процессы в живой клетке контролируются ферментами, следовательно, для фотосинтеза будет иметь значение и температура окружающей среды. Главным лимитирующим фактором является тот, который в данный момент времени более всего затрудняет фотосинтез. Так, если света совсем мало, лимитирующим фактором становится свет. При улучшении освещения до нормальных значений свет уже перестает ограничивать фотосинтез, и лимитирующим фактором становится какой-то иной. Например, количество углекислого газа в воздухе, увеличение его концентрации вызовет повышение уровня фотосинтеза. Впервые ввел понятие и описал роль лимитирующих факторов английский биохимик Фредерик Блэкман (1866–1947) в 1905 г.
Во времена Блэкмана ученые не знали того, что знаете вы сейчас. Не были еще изучены биохимические механизмы цикла Кальвина и взаимодействие света с хлорофиллом, не было понятия о процессе фотолиза воды. Было известно только то, что углекислый газ, свет и вода исчезают в листе. Вместо них образуются кислород, глюкоза или крахмал (молекулы которого получаются из молекул глюкозы путем полимеризации – однотипного последовательного соединения). Суть работы Блэкмана заключалась в том, что он изменял два условия в жизни растения – количество света и углекислого газа в воздухе. Именно после этих опытов он предположил, что фотосинтез состоит из двух фаз: быстрой – световой и медленной – темновой.
Значения лимитирующих факторов сегодня изучены достаточно хорошо (рис. 9). Так температура окружающей среды, после повышения которой интенсивность фотосинтеза перестает расти, составляет 25°С. В организмах животных эффективность работы ферментов продолжает расти при повышении температуры примерно до 40°С. Но у растений при повышении температуры выше 25°С эффективность фотосинтеза перестает возрастать. При дальнейшем повышении температуры даже снижается. Видимо, происходит закрывание устьиц, и организм растения начинает экономить воду. Зеленым клеткам мякоти листа начинает не хватать углекислого газа, так как через закрытые устьица он не может попасть в растение. Лимитирующим фактором в данном случае становится не столько слишком высокая температура, сколько недостаток СО 2 .
Рис. 9. Факторы окружающей среды (внешние), влияющие на скорость и интенсивность процесса фотосинтеза
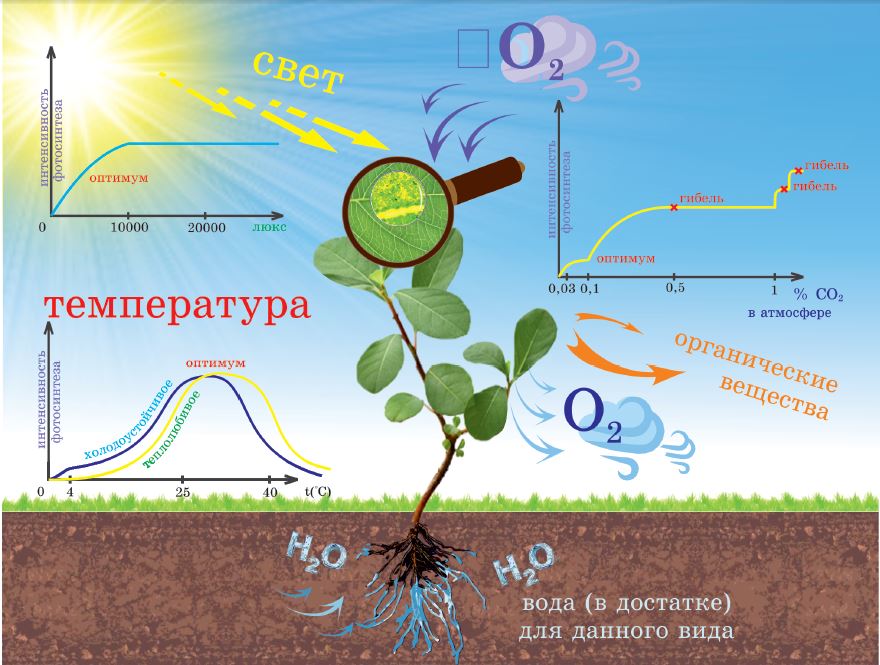
Вода – одно из условий жизнедеятельности организма растений вообще и фотосинтеза в частности. Кроме того, что вода используется в реакциях фотолиза, она влияет и на все другие жизненные процессы, определяет вязкость цитоплазмы, состав клеточного сока, тургорное давление (см. § 14, 15), транспорт веществ по растению, эффективность работы ферментов и т. д. Ученые выявили, что даже если растения испытывают незначительный, временный недостаток влаги, приводящий к увяданию, их урожайность заметно снижается.
Свет – важнейший фактор фотосинтеза. Сколько света нужно растениям, чтобы эффективность фотосинтеза была максимальной? Ученые называют цифру – 10 000 люкс. Для сравнения: освещение в яркий (непасмурный), летний, солнечный день соответствует 100 000 люкс. Следовательно, в естественных природных условиях освещать растение сильнее не имеет смысла, ведь интенсивность фотосинтеза не будет при этом возрастать.
Углекислый газ в современной атмосфере имеет концентрацию 0,03%. Точно установлено, что эффективность фотосинтеза у всех видов растений будет возрастать с повышением концентрации СО 2 от 0,3 до 0,5%. Большинство видов растений отрицательно реагирует на длительное воздействие СО 2 концентрацией выше 0,5%. Так тропические виды, экспериментально содержавшиеся в условиях с концентрацией 0,5% СО 2 , интенсивно развивались, но очень быстро старели и погибали. Дальнейшая эффективность повышения СО 2 под вопросом, но в искусственно созданной атмосфере даже с такой высокой концентрацией СО 2 , как 2,5–5%, мгновенной гибели растений не наступало. Принято считать, что даже если оптимальная концентрация СО 2 зависит от вида растений, среднее ее значение составляет 0,1%.
Лимитирующими факторами могут быть загрязняющие или ядовитые агенты, находящиеся в окружающей среде. Так пыль, оседающая на листья, препятствует проникновению солнечного света, осаждаясь на поверхности листа. Забивая устьица, препятствует газообмену с воздухом. Нарушают функционирование листьев и подавляют фотосинтез различные промышленные загрязнители, такие как диоксид серы, сажа, соединения меди и других металлов. Действие некоторых гербицидов, направленное против сорняков, тоже основано на веществах, подавляющих фотосинтез.
Практическое значение изучения лимитирующих факторов велико, так как эффективность фотосинтеза у культурных растений непосредственно влияет на их урожайность. Чтобы добиться максимальной урожайности, важно знать оптимальные условия фотосинтеза. Ученые совершенствуют методы повышения эффективности синтеза органических веществ, отдельные для каждого сорта культурных растений. Одним из ученых, уделявших большое внимание изучению роли света и хлорофилла в процессе фотосинтеза, был Климент Аркадьевич Тимирязев.
Читайте также: