
Роль заднего мозга в регуляции фазных движений и мышечного тонуса физиология кратко
Обновлено: 07.07.2024
1. Роль отделов ЦНС в регуляции мышечного тонуса и фазных движений
Иван Михайлович Сеченов
Всё бесконечное разнообразие внешних
проявлений мозговой деятельности
сводится окончательно к одному лишь
явлению – мышечному движению.
3. Общая характеристика движения
Основным компонентом поведения
человека является двигательная
активность
Виды движения:
1. активные: непроизвольные,
произвольные, автоматизированные
2. пассивные (без сокращения мышц)
3. локомоции (как совокупность
согласованных движений – перемещение
в пространстве)
4. Компоненты двигательного акта
Моторный: тонический и
фазический
Вегетативный: безусловно- и
условнорефлекторный
Сенсорный (двигательный
анализатор)
Нервная регуляция
соответствия позы и
движения, их правильного
сопряжения – одна из
важнейших функций
двигательных центров
6. … и все это – строго координированная деятельность нервной системы и скелетных мышц.
8. Уровни управления движениями
Участки коры больших полушарий
специализированы для выполнения
различных функций
Программирование и управление
движениями осуществляет моторная
кора и премоторная кора
10. ПОЛЯ БРОДМАНА
О функциях лобных
долей
Фейнис Гейдж - рабочий железной дороги штата
Вермонт в 1886 году в результате взрыва
получил тяжелую травму головы.
14. Двигательный гомункулюс
Введение
Задний мозг развивается из заднемозгового пузыря, который является производным ромбовидного пузыря. Он состоит из Варолиева моста, который несет в своем составе полость заднего мозга – часть ромбовидной ямки и мозжечка,который представляет собой дорсальный вырост заднего мозга.
Рис. 1. Расположение мозжечка на стволе мозга (часть ткани мозжечка убрана):
1 — верхняя ножка мозжечка; 2 — средняя ножка мозжечка; 3 — лист (долька мозжечка); 4 — левое полушарие мозжечка; 5 — спинной мозг; 6 — продолговатый мозг; 7 — нижняя ножка мозжечка; 8 — мост; 9 — средний мозг
Основные функции заднего мозга проводниковая ирефлекторная.
Через задний мозг проходят нисходящие пути (кортикоспинальный и экстрапирамидный), восходящие – ретикуло– и вестибулоспинальный, отвечающие за перераспределение мышечного тонуса и поддержание позы тела.
1.Роль заднего мозга в регуляции фазных движений.
До этого отдела ЦНС относятся продолговатый мозг и мост головного мозга. Основные функции заднего мозга в регуляции двигательных функций: 1) анализ сенсорной информации, поступающей в ядра заднего мозга от рецепторов кожи и мышц головы и эфферентная иннервация скелетных мышц головы;
2) анализ сенсорной информации, поступающей от вестибулярных рецепторов, проприорецепторов мышц шеи, туловища, конечностей, и рефлекторная регуляция тонуса мышц тела, которая обеспечивает определенную позу (тонические рефлексы позы);
3) проводниковаяфункция - прохождение нервных импульсов, идущих в вышележащие отделы ЦНС и от них до скелетных мышц.
Сенсорная информация поступает от рецепторов кожи лица, слизистых оболочек носа, зубов, надкостница костей черепа, проприорецепторов глазодвигательных мышц, вкусовых рецепторов, рецепторов вестибулярного аппарата и слуха, глотки, рецепторов многих внутренних органов.
Эфферентная иннервация обеспечиваетфункцию жевательных, глазодвигательных, мимических мышц, мышц языка, глотки, гортани, шеи. Эти функции обеспечиваются нервами, еще сохраняют способность к сегментарной иннервации, но она выражена менее отчетливо, чем в спинном мозге.
Очень большую роль играет задний мозг в поддержании позы антигравитации (обеспечивается тонусом мышц) и перераспределении тонуса при изменении положения головы. Жаба, вкоторой перерезан мозг между продолговатым и спинным (так называемая спинальная животное), не может поддерживать тонус скелетных мышц на таком уровне,
которого было бы достаточно для создания ПОЗЫ антигравитации. Такое животное лежит распластанный на горизонтальной поверхности.
Жаба, в которой перерезан мозг выше заднего, имеет позу, которая позволяет преодолеть силу гравитации Земли. Для создания такой позыважное значение имеет тонус мышц-екстензорив. Этот тонус имеет рефлекторную природу, ибо исчезает после перерезания задних корешков нервов, входящие в спинной или продолговатый мозг. Центрами регуляции есть три двигательные ядра заднего мозга: 1) вестибулярное ядро, 2) ядра ретикулярной формации варолиевого моста, 3) ядра ретикулярной формации продолговатого мозга.
С участием этих центров реализуютсятонические рефлексы позы, среди которых различают вестибулярные статические и шейные.
Вестибулярные статические рефлексы начинаются с рецепторов преддверия - сферического и эллиптического мешочков. Они были подробно исследованы голландским физиологом R. Magnus (1924).
Существует несколько видов вестибулярных статических рефлексов:
1) при подъеме головы повышается тонус мышц-екстензорив переднихконечностей и снижается тонус этих мышц задних конечностей;
2) при наклоне головы вниз наблюдается противоположный эффект;
3) при наклоне головы в сторону повышается тонус мышц-екстензорив с той стороны, куда наклонена голова.
Значение этих рефлексов заключается в том, что происходит перераспределение тонуса мышц при изменении положения головы, и это обеспечивает.
Контроль движений различными отделами нервной системы
Интеграция разных отделов общей системы регуляции движений. Теперь попробуем объединить, насколько это возможно, все, что мы знаем об общем контроле движений. Для этого сначала проведем краткий обзор разных уровней регуляции.
а) Уровень спинного мозга. В спинном мозге заложены программы локальных движений для всех мышечных областей тела, например рефлексы отдергивания, помогающие убрать любую часть тела от источника боли. На уровне спинного мозга осуществляются также сложные ритмические движения, например движения конечностей вперед-назад, характерные для ходьбы с одновременными реципрокными движениями на противоположной стороне тела или реципрокными отношениями между задними и передними конечностями у четвероногих животных.
Все эти программы спинного мозга могут приводиться в действие вышерасположенными уровнями регуляции двигательной активности или затормаживаться, когда высшие уровни принимают контроль на себя.
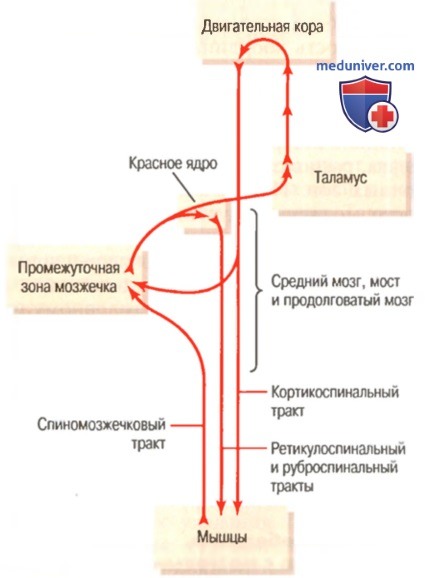
Участие коры больших полушарий и мозжечка, особенно его промежуточной зоны, в регуляции произвольных движений
б) Уровень ромбовидного мозга. Ромбовидный мозг обеспечивает две важные функции в общей регуляции двигательной активности организма:
(1) поддержание тонуса осевых мышц тела для обеспечения стояния;
(2) постоянное изменение степени тонического напряжения различных мышц в ответ на информацию от вестибулярного аппарата для поддержания равновесия тела.
1. Сопряженные функции мозжечка. Мозжечок функционирует вместе со всеми уровнями двигательного контроля. Он тесно связан со спинным мозгом, особенно для усиления рефлекса на растяжение, поэтому когда сокращающаяся мышца сталкивается с неожиданно тяжелой нагрузкой, длительный сигнал рефлекса на растяжение, передаваемый в мозжечок и обратно к спинному мозгу, значительно усиливает эффект сопротивления нагрузке основного рефлекса на растяжение.
На уровне ствола мозга функция мозжечка обеспечивает плавность и непрерывность постуральных движений тела (без патологических колебаний), особенно быстрых движений, необходимых для поддержания равновесия.
На уровне коры большого мозга мозжечок действует в связи с корой, обеспечивая много вспомогательных двигательных функций, особенно придавая дополнительную двигательную силу для быстрого включения мышечного сокращения в начале движения. Ближе к окончанию каждого движения мозжечок включает мышцы-антагонисты в точно определенный момент и с соответствующей силой, чтобы остановить движение в запланированной точке. Более того, есть достоверные физиологические данные о том, что все аспекты этого функционирования мозжечка по принципу включение-выключение могут совершенствоваться в процессе жизненного опыта.
Мозжечок функционирует совместно с корой больших полушарий еще на одном уровне моторного контроля: он помогает программировать заранее мышечные сокращения, которые требуются для плавного перехода от текущего быстрого движения в одном направлении к следующему быстрому движению в другом направлении, причем все это осуществляется в течение доли секунды. Нервный контур для этого проходит от коры большого мозга к большим латеральным зонам полушарий мозжечка и затем назад — к коре большого мозга.
Мозжечок функционирует, когда мышечные движения должны осуществляться быстро. Без мозжечка медленные и сознательные движения могут еще происходить, но кортикоспинальной системе трудно доводить до конца быстрые, меняющиеся движения, предназначенные для достижения особой цели, или гладко переходить от одного быстрого движения к следующему.
Связь контура базальных ганглиев с кортикоспиномозжечковой системой для регуляции двигательной активности Контур скорлупы системы базальных ганглиев для подсознательного выполнения программ приобретенных движений
2. Сопряженные функции базальных ганглиев. Базальные ганглии участвуют в регуляции движений совершенно иным путем, чем мозжечок. Их наиболее важными функциями являются: (1) помощь коре в выполнении подсознательных, но приобретенных двигательных программу (2) содействие планированию множественных параллельных и последовательных программ движения, которые разум должен собрать вместе для достижения намеченной цели.
К двигательным программам, требующим участия базальных ганглиев, относят, например, программы для написания различных букв, бросания мяча и печатания на пишущей машинке. Базальные ганглии также нужны для модификации этих программ, чтобы писать буквы мелко или очень крупно, т.е. регулировать амплитуду движений при выполнении двигательной программы.
На еще более высоком уровне регуляции существует другой совместный контур коры большого мозга и базальных ганглиев. Он начинается с мыслительных процессов в коре и обеспечивает общую последовательность действий в ответ на каждую новую ситуацию, например планирование немедленной двигательной реакции в ответ на удар в лицо со стороны противника или на неожиданное нежное объятие.
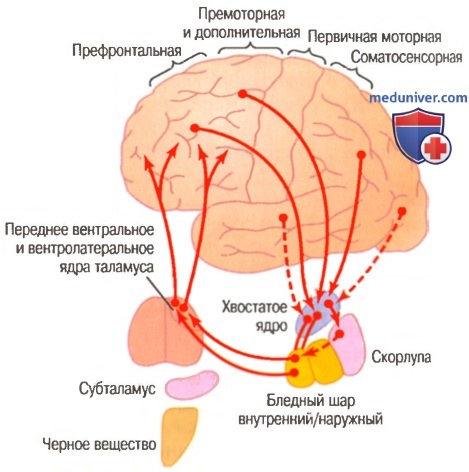
Контур хвостатого ядра системы базальных ганглиев для когнитивного планирования последовательных и параллельных двигательных программ для достижения специфических осознанных целей
г) Что побуждает нас действовать? Что заставляет нас переходить от состояния покоя к активным действиям? Мы только начинаем изучать мотивационные системы мозга. По существу, мозг имеет более старое ядро, расположенное ниже, впереди и латеральнее таламуса, включающее гипоталамус, миндалевидное тело, гиппокамп, область перегородки впереди гипоталамуса и таламуса и даже старые регионы самого таламуса и коры большого мозга. Действуя совместно, эти отделы инициируют большинство моторных и других функциональных активностей мозга. Совокупность этих структур называют лимбической системой мозга. Мы подробно обсудим эту систему в отдельной статье на сайте (просим вас пользоваться формой поиска выше).
Видео физиология движений (тонус, поза, произвольные движения) - профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021

Рецепторным элементом миотатического рефлекса является инкапсулированное мышечное веретено. Каждая мышца содержит большое количество этих рецепторов. Мышечное веретено состоит из интрафузальных мышечных волокон (в сравнении с обычными экстрафузальными мышечными волокнами они значительно тоньше) и ядерной сумки, оплетенной спиралевидной сетью тонких нервных волокон, представляющих собой первичные чувствительные окончания. На некоторых интрафузальных волокнах имеются еще и вторичные, гроздевидные чувствительные окончания. При растяжении интрафузальных волокон первичные чувствительные окончания усиливают исходящую из них импульсацию, которая через быстро проводящие афферентные волокна типа Iа проводятся к альфа-большим мотонейронам спинного мозга. Оттуда через также быстропроводящие альфа-1-эфферентные волокна импульс идет к экстрафузальным белым мышечным волокнам, которые обеспечивают быстрое (фазическое) сокращение мышцы. От вторичных чувствительных окончаний, реагирующих на тонус мышцы, афферентная импульсация проводится по тонким волокнам II типа через систему вставочных нейронов к альфа-малым мотонейронам, которые иннервируют тонические экстрафузальные мышечные волокна (красные мышечные волокна), обеспечивающие поддержание тонуса и позы.
В регуляции мышечного тонуса принимают участие и тормозные механизмы, которые реализуются с помощью рецепторов Гольджи, расположенных в сухожилиях мыщц, и вставочных клеток Реншоу, находящихся в передних рогах спинного мозга. Сухожильные рецепторы Гольджи при растяжении или значительном напряжении мышцы посылают афферентные импульсы, которые проводятся по быстропроводящим волокнам 1b типа в спинной мозг и оказывают тормозящее воздействие на мотонейроны передних рогов. Вставочные нейроны Реншоу активизируются через коллатерали при возбуждении альфа-мотонейронов и действуют по принципу отрицательной обратной связи, тормозя активность последних. Таким образом, нейрогенные механизмы регуляции мышечного тонуса многообразны и сложны.
Функция поддержания мышечного тонуса обеспечивается по принципу обратной связи на различных уровнях регуляции организма Периферическая регуляция осуществляется с участием гамма-петли, в состав которой входят супраспинальные моторные пути, вставочные нейроны, нисходящая ретикулярная система, альфа- и гамма-нейроны.
Существует два типа гамма-волокон в передних рогах спинного мозга. Гамма-1-волокна обеспечивают поддержание динамического мышечного тонуса, т.е. тонуса, необходимого для реализации процесса движения. Гамма-2-волокна регулируют статическую иннервацию мышц, т.е. осанку, позу человека. Центральная регуляция функций гамма-петли осуществляется ретикулярной формацией через ретикулоспинальные пути. Основная роль в поддержании и изменении мышечного тонуса отводится функциональному состоянию сегментарной дуги рефлекса растяжения (миотатического, или проприоцептивного рефлекса). Рассмотрим его подробнее.
Рецепторным элементом его является инкапсулированное мышечное веретено. Каждая мышца содержит большое количество этих рецепторов. Мышечное веретено состоит из интрафузальных мышечных волокон (тонких) и ядерной сумки, оплетенной спиралевидной сетью тонких нервных волокон, представляющих собой первичные чувствительные окончания (анулоспинальная нить). На некоторых интрафузальных волокнах имеются также и вторичные, гроздевидные чувствительные окончания. При растяжении интрафузальных мышечных волокон первичные чувствительные окончания усиливают исходящую из них импульсацию, которая через быстропроводящие гамма-1-волокна проводится к альфа-большим мотонейронам спинного мозга. Оттуда, через также быстропроводящие альфа-1-эфферентные волокна, импульс идет к экстрафузальным белым мышечным волокнам, которые обеспечивают быстрое (фазическое) сокращение мышцы. От вторичных чувствительных окончаний, реагирующих на тонус мышцы, афферентная импульсация проводится по тонким гамма-2-волокнам через систему вставочных нейронов к альфа-малым мотонейронам, которые иннервируют тонические экстрафузальные мышечные волокна (красные), обеспечивающие поддержание тонуса и позы.
Интрафузальные волокна иннервируются гамма-нейронами передних рогов спинного мозга. Возбуждение гамма-нейронов, передаваясь по гамма-волокнам к мышечному веретену, сопровождается сокращением полярных отделов интрафузальных волокон и растяжением их экваториальной части, при этом изменяется исходная чувствительность рецепторов к растяжению (происходит снижение порога возбудимости рецепторов растяжения, и усиливается тоническое напряжение мышцы).
Гамма-нейроны находятся под влиянием центральных (супрасегментарных) воздействий, передающихся по волокнам, которые идут от мотонейронов оральных отделов головного мозга в составе пирамидного, ретикулоспинального, вестибулоспинального трактов.
При этом если роль пирамидной системы заключается преимущественно в регуляции фазических (т.е. быстрых, целенаправленных) компонентов произвольных движений, то экстрапирамидная система обеспечивает их плавность, т.е. преимущественно регулирует тоническую иннервацию мышечного аппарата. Так, по мнению J. Noth (1991), спастичность развивается после супраспинального или спинального поражения нисходящих двигательных систем при обязательном вовлечении в процесс кортикоспинального тракта [2] .
В регуляции мышечного тонуса принимают участие и тормозные механизмы, без которых невозможно реципрокное взаимодействие мышц-антагонистов, а значит, невозможно и совершение целенаправленных движений. Они реализуются с помощью рецепторов Гольджи, расположенных в сухожилиях мышц, и вставочных клеток Реншоу, находящихся в передних рогах спинного мозга. Сухожильные рецепторы Гольджи при растяжении или значительном напряжении мышцы посылают афферентные импульсы по быстропроводящим волокнам 1б-типа в спинной мозг и оказывают тормозящее воздействие на мотонейроны передних рогов. Вставочные клетки Реншоу активизируются через коллатерали при возбуждении альфа-мотонейронов, и действуют по принципу отрицательной обратной связи, способствуя торможению их активности. Таким образом, нейрогенные механизмы регуляции мышечного тонуса многообразны и сложны.
При поражении пирамидного пути растормаживается гамма-петля, и любое раздражение путем растяжения мышцы приводит к постоянному патологическому повышению мышечного тонуса. При этом поражение центрального мотонейрона приводит к снижению тормозных влияний на мотонейроны в целом, что повышает их возбудимость, а так же на вставочные нейроны спинного мозга, что способствует увеличению числа импульсов, достигающих альфа-мотонейронов в ответ на растяжение мышцы [4].
В качестве других причин спастичности можно указать структурные изменения на уровне сегментарного аппарата спинного мозга, возникающие вследствие поражения центрального мотонейрона: укорочение дендритов альфа-мотонейронов и коллатеральный спрауттинг (разрастание) афферентных волокон, входящих в состав задних корешков.
Возникают так же и вторичные изменения в мышцах, сухожилиях и суставах. Поэтому страдают механико-эластические характеристики мышечной и соединительной ткани, которые определяют мышечный тонус, что еще больше усиливает двигательные расстройства.
Следствием этого является спастичность мышц, гиперрефлексия, появление патологических рефлексов, а также первоочередная утрата наиболее тонких произвольных движений [5].
Наиболее значимым компонентом мышечного спазма является боль. Болевая импульсация активирует альфа- и гамма-мотонейроны передних рогов, что усиливает спастическое сокращение мышцы, иннервируемой данным сегментом спинного мозга. В то же время, мышечный спазм, возникающий при сенсомоторном рефлексе, усиливает стимуляцию ноцицепторов мышцы. Так, по механизму отрицательной обратной связи формируется замкнутый порочный круг: спазм – боль – спазм – боль [3].
Интерес представляют также данные о центральных холинергических механизмах регуляции мышечного тонуса. Показано, что клетки Реншоу активируются ацетилхолином как через коллатерали мотонейрона, так и через ретикулоспинальную систему.
M.Schieppati и соавт., (1989) установили, что фармакологическая активация центральных холинергических систем значительно снижает возбудимость альфа-мотонейронов путем повышения активности клеток Реншоу.
В последние годы исследователи регуляции мышечного тонуса придают огромное значение роли нисходящих адренергических супраспинальных путей, начинающихся в области голубого пятна. Анатомически эти образования тесно связаны со спинальными структурами, особенно с передними рогами спинного мозга. Норадреналин, высвобождаемый с терминалей бульбоспинальных волокон, активизирует адренорецепторы, располагающиеся во вставочных нейронах, первичных афферентных терминалях и мотонейронах и воздействует одновременно на альфа- и бета-адренорецепторы в спинном мозге (D.Jones et al., 1982). К ядерным образованиям ретикулярной формации ствола подходят многочисленные аксоны болевой чувствительности. На основе информации, поступающей в ретикулярную формацию ствола головного мозга, выстраиваются соматические и висцеральные рефлексы. От ядерных образований ретикулярной формации формируются связи с таламусом, гипоталамусом, базальными ядрами и лимбической системой, которые обеспечивают реализацию нейроэндокринных и аффективных проявлений боли, что особенно важно при хронических болевых синдромах [2].
В итоге формирующийся порочный круг включает в себя мышечный спазм, боль, локальную ишемию, дегенеративные изменения, которые самоподдерживают друг друга, усиливая первопричину патологических изменений.
Следует учитывать, что чем больше компонентов этого порочного круга становятся мишенями при лечении, тем выше вероятность его успеха. Поэтому современными требованиями к миорелаксирующей терапии являются: мощность миорелаксирующего действия, его селективность, наличие противосудорожного и антиклонического эффектов, мощность анальгетического действия, а так же безопасность и наличие широкого терапевтического диапазона доз препарата.
Согласно современным представлениям, большинство миорелаксантов воздействуют на трансмиттеры или нейромодуляторы ЦНС. Воздействие может включать супрессию возбуждающих медиаторов (аспартат и глутамат) и/или усиление тормозных процессов (ГАМК, глицин). Далее
Читайте также:
- Сотрудничество родителей и педагогов в школе
- Дзержинский 6 сад московской детский области со скольки берут в сад
- Важная черта общения и поведения необходимая для успешного обучения в школе 14 букв
- Григорьянц а г методика обучения географии во вспомогательной школе
- Тематическое планирование индивидуального обучения 4 класс школа россии