
Роль нуклеотидов в обмене веществ кратко
Обновлено: 02.07.2024
Нуклеоти́ды — фосфорные эфиры нуклеозидов, нуклеозидфосфаты. Свободные нуклеотиды, в частности АТФ, цАМФ, АДФ, играют важную роль в энергетических и информационных внутриклеточных процессах, а также являются составляющими частями нуклеиновых кислот и многих коферментов.
Нуклеотиды являются сложными эфирами нуклеозидов и фосфорных кислот. Нуклеозиды, в свою очередь, являются N-гликозидами, содержащими гетероциклический фрагмент.В природе наиболее распространены нуклеотиды, являющиеся β-N-гликозидами пуринов или пиримидинов и пентоз — D-рибозы или D-2-дезоксирибозы. В зависимости от структуры пентозы различают рибонуклеотиды и дезоксирибонуклеотиды, которые являются мономерами молекул сложных биологических полимеров (полинуклеотидов) — соответственно РНК или ДНК.
Фосфатный остаток в нуклеотидах обычно образует сложноэфирную связь с 2'-, 3'- или 5'-гидроксильными группами рибонуклеозидов, в случае 2'-дезоксинуклеозидов этерифицируются 3'- или 5'-гидроксильные группы.
Большинство нуклеотидов являются моноэфирами ортофосфорной кислоты, однако известны и диэфиры нуклеотидов, в которых этерифицированы два гидроксильных остатка — например, циклические нуклеотиды циклоаденин- и циклогуанин монофосфаты.
Наряду с нуклеотидами — эфирами ортофосфорной кислоты (монофосфатами) в природе также распространены и моно- и диэфиры пирофосфорной кислоты (дифосфаты,) и моноэфиры триполифосфорной кислоты (трифосфаты).
8. Строение фосфолипидов. Роль фосфолипидов в организме.
Фосфолипи́ды — сложные липиды, сложные эфиры многоатомных спиртов и высших жирных кислот. Содержат остаток фосфорной кислоты и соединенную с ней добавочную группу атомов различной химической природы.
Фосфолипиды входят в состав всех клеточных мембран.
Между плазмой и эритроцитами происходит обмен фосфолипидами, которые играют важнейшую роль, поддерживая в растворимом состоянии неполярные липиды.
Наиболее распространенная группа Фосфолипидов — фосфоглицериды, также к фосфолипидам относятся фосфосфинголипиды и фосфоинозитиды.
Главный липидный компонент клеточных мембран. Они сопутствуют жирам в пище и служат источником фосфорной кислоты, необходимый для жизни человека.
Фосфолипиды являются важной частью клеточных мембран. Они обеспечивают текучие и пластические свойства мембран клеток и клеточных органоидов, в то время как холестерин обеспечивает жёсткость и стабильность мембран. Как фосфолипиды, так и холестерин часто входят в состав липопротеидов клеточных мембран, но имеются в мембранах и в свободном, не связанном с белками состоянии. Соотношение холестерин/фосфолипиды в основном и определяет текучесть либо жёсткость клеточной мембраны.
Фосфолипиды замедляют синтез коллагена и повышают активность коллагеназы (фермента, разрушающего коллаген). Поскольку коллаген определяет замещение эпителиальной ткани соединительной, фосфолипиды оказывают противорубцовый (антифибротический) эффект.
Свидетельство и скидка на обучение каждому участнику
Зарегистрироваться 15–17 марта 2022 г.

Описание презентации по отдельным слайдам:
БИОХИМИЯ НУКЛЕИНОВЫХ КИСЛОТ

БИОХИМИЯ НУКЛЕИНОВЫХ КИСЛОТ
Явление передачи наследственной информации из поколения в поколение связано с нуклеиновыми кислотами.
Впервые нуклеиновые кислоты были выделены швейцарским врачом Ф.Мишером (1868 г.) из ядер клеток и названы нуклеинами (от лат.nucleus – ядро). Ф.Мишер определил, что в состав нуклеина входят атомы углерода, водорода, кислорода, азота и фосфора. В 1889 г. Р.Альтман показал, что нуклеин имеет кислые свойства и предложил назвать эти соединения нуклеиновыми кислотами.
Нуклеиновые кислоты имеют более сложную структуру, чем белки. Mm – несколько десятков или сотен миллионов. Нуклеиновые кислоты – это генетический материал живых клеток.
Нуклеиновые кислоты – это высокомолекулярные соединения, состоящие из большого количества связанных между собой нуклеотидов.
ХИМИЧЕСКИЙ СОСТАВ НУКЛЕИНОВЫХ КИСЛОТ
При полном гидролизе нуклеиновых кислот образуется несколько простых низкомолекулярных органических веществ: азотистые основания, углеводы, молекулы фосфорной кислоты.
АЗОТИСТЫЕ ОСНОВАНИЯ – это производные гетероциклического азотосодержащего соединения ПУРИНА и ПИРИМИДИНА. К пуриновым основаниям относятся аденин (А) и гуанин (Г), к пиримидиновым основаниям относятся – цитозин (Ц), тимин (Т) и урацил (У). Их молекулы различаются наличием определенных функциональных групп: - NH2, -OH, - CH3.
Из пяти азотистых оснований каждая нуклеиновая кислота включает только четыре – два пуриновых и два пиримидиновых.
Углеводный компонент нуклеиновых кислот – моносахариды: рибоза и дезоксирибоза. Фосфорная кислота входит в состав всех нуклеиновых кислот в большом количестве.

ОСНОВНЫЕ СОСТАВЛЯЮЩИЕ НУКЛЕИНОВЫХ КИСЛОТ: АЗОТИСТЫЕ ОСНОВАНИЯ И УГЛЕВОДЫ ИЗ КЛАССА ПЕНТОЗ

НУКЛЕОЗИДЫ И НУКЛЕОТИДЫ
При неполном гидролизе нуклеиновых кислот образуются нуклеозиды и молекулы фосфорной кислоты, а также нуклеотиды.
НУКЛЕОЗИД – это органические вещества, в состав которых входит пентоза и гетероциклическое азотистое основание.
Структурной единицей нуклеиновых кислот является нуклеотид. Нуклеотиды состоят из трех компонентов: пуринового или пиримидинового основания, углевода рибозы или дезоксирибозы, которые соединены между собой и составляют нуклеозид и остатка фосфорной кислоты, присоединенной к гидроксилу пятого атома углерода в молекуле углевода.
Пример нуклеотида : аденозинмонофосфат (АМФ) или адениловая кислота:
Основными нуклеотидмонофосфатами - производными пуриновых оснований являются аденозин-5-монофосфат (АМФ)(аденилат) и гуанозин-5-монофосфат (ГМФ) (гуанилат). Из них образуются нуклеотиддифосфаты и нуклеотидтрифосфаты (АТФ, ГТФ, АДФ, ГДФ)
Основными нуклеотидмонофосфатами – производными пиримидиновых оснований являются цитидин-5-монофосфат (ЦМФ) (цитидилат); уридин-5-монофосфат (УМФ) (уридилат); тимин-5-монофосфат (ТМФ) (тимидилат). Из них обаразуются нуклеотиддифосфаты и нуклеотидтрифосфаты (ЦТФ, УТФ, ТТФ).
Поскольку в состав нуклеотидов входят остатки фосфорной кислоты, они обладают кислотными свойствами и называются кислотами

Нуклеотиды именуют по названию соответствующего нуклеозида с указанием положения и числа фосфорильных групп. По числу фосфорильных групп нуклеотиды делят на моно-, ди- и трифосфаты:

Для цАМФ характерна значительная энергия связи между 3- и 5- углеродными атомами в рибозе. При ее гидролизе выделяется около 59 кДж.моль -1 энергии.
цАМФ – это универсальный внутриклеточный передатчик действия гормонов.
РОЛЬ циклических нуклеотидов:
Участвуют в регуляции процессов транспорта ионов через клеточные мембраны.
Участвуют в процессе распада углеводов и жиров
Регуляторная роль в процессах клеточной дифференцировки.


СТРОЕНИЕ НУКЛЕИНОВЫХ КИСЛОТ
ДНК – имеет большую молекулярную массу - от нескольких миллионов до 2-5 миллиардов, так как состоит из 50 тысяч и более нуклеотидов.
В состав ДНК входят: углевод дезоксирибоза и азотистые основания – АДЕНИН, ГУАНИН, ЦИТОЗИН, ТИМИН. Нуклеотиды соединены между собой 3’, 5’ – фосфодиэфирными связями, образуя полинуклеотидную цепь. Каждый нуклеотид в структуре полинуклеотидной цепи называют нуклеотидным остатком.
ПЕВИЧНАЯ СТРУКТУРА нуклеиновых кислот – это порядок чередования нуклеотидных остатков в полинуклеотидной цепи. Вариабельной частью в нуклеотидных остатках являются азотистые основания

Вторичная и третичная структуры ДНК
В 1953 г. Д.Уотсон и Ф.Крик открыли вторичную структуру ДНК. Методом рентгеноструктурного анализа установили, что ДНК состоит более чем из одной полидезоксирибонуклеотидной цепи и обладает спиральной структурой, имеющей определенные закономерности. Э.Чаргафф определил важные количественные соотношения отдельных азотистых оснований ДНК.
ПРАВИЛО ЧАРГАФФА:
Суммарное содержание пуриновых нуклеотидов равно суммарному содержанию пиримидиновых нуклеотидов.
Содержание тимина равно содержанию аденина.
Содержание гуанина равно содержанию цитозина.
Эти правила говорят о том, что при построении ДНК должно соблюдаться строгое соответствие не пуриновых и пиримидиновых оснований, а конкретно аденина и тимина, гуанина и цитозина
Кислотно-основное титрование ДНК показало, что ее структура стабилизирована водородными связями.
Д.Уотсон и Ф.Крик построили модель вторичной структуры ДНК, получившей название двойной спирали.

ВТОРИЧНАЯ СТРУКТУРА МОЛЕКУЛЫ ДНК
ОСНОВНЫЕ ХАРАКТЕРИСТИКИ ДВОЙНОЙ СПИРАЛИ ДНК:
ДНК представляет собой спираль, закрученную вправо относительно оси.
Две цепи в спирали ДНК антипараллельны, т.е. их 3’, 5’ – межнуклеотидные мостики (связи) направлены в противоположные стороны.
Остовы полинуклеотиднх цепей расположены на внешней стороне двойной спирали – обращены в сторону водной среды.
Пуриновые и пиримидиновые азотистые основания уложены стопкой внутри двойной спирали; основания сближены между собой и расположены перпендикулярно оси симметрии двойной спирали.
Диаметр спирали – 2нм; длина витка – 3,4 нм; расстояние между соседними основаниями – 0,34 нм.
Благодаря такой структуре обеспечивается тесное взаимодействие пар оснований посредством водородных связей. Такие связи образуются между соответствующими – КОМПЛЕМЕНТАРНЫМИ – азотистыми основаниями: аденин комплементарен тимину, гуанин – цитозину. Образуя водородные связи, азотистые основания располагаются в плоскости, перпендикулярной оси ДНК. В стабилизации вторичной структуры ДНК принимают участие катионы металлов и специфические белки.

КОМПЛЕМЕНТАРНОСТЬ АЗОТИСТЫХ ОСНОВАНИЙ НА УЧАСТКЕ ДВОЙНОЙ СПИРАЛИ ДНК

СХЕМА УДВОЕНИЯ (РЕПЛИКАЦИЯ) ДНК

ПОЛИМОРФИЗМ ДНК – в зависимости от условий ДНК может менять свою конформацию. Методом рентгеноструктурного анализа детально изучены А-, В- С- и Т – формы ДНК. Все формы биспиральны, но имеют отличия по ряду параметров.

ТРЕТИЧНАЯ СТРУКТУРА ДНК
В результате дополнительного скручивания в пространстве двойной спирали образуется третичная структура ДНК. В эукариотической клетке ДНК находится в составе хроматина – комплекса ДНК с белками. Хроматин состоит из белков (60%), ДНК (35%) и РНК (5%). Структурной единицей хроматина является нуклеосома. В нуклеосоме ДНК тесно связана с белками гистонами, характерной особенностью которых является наличие в их структуре от 20 до 30% основных аминокислот – лизина, аргинина, гистидина, что определяет их суммарный положительный заряд. Пять классов гистонов: Н1, Н2А, Н2В, Н3, Н4 – отличаются первичной структурой и молекулярной массой. В нуклеосоме гистоны Н2А, Р2В, Н3, Н3 образуют октамер – это ГИСТОНОВЫЙ КОР. ДНК – полианион дважды обвивает гистоновый кор. В промежутках между нуклеосомами расположена соединительная (линкерная) ДНК, с которой связан гистон Н1. Длина соединительных участков ДНК в хроматиновых волокнах человека составляет около 50 нуклеотидных пар. За счет образования нуклеосом достигается плотная упаковка огромной молекулы ДНК в ядре.

БИОЛОГИЧЕСКАЯ РОЛЬ ДНК
Основная функция ДНК – хранение и передача наследственной или генетической информации через посредника (мРНК) к белку.
Все признаки организма проявляются через свойства синтезирующихся в нем белков.
Структура каждого белка закодирована в отдельных участках ДНК
Участок ДНК, выполняющий определенные функции, называют геном. Гены: структурные и регуляторные. Структурные гены несут информацию о структуре различных видов РНК. Если структурный ген содержит информацию о матричной РНК, то в нем заложена информация и о первичной структуре белка. Регуляторные белки участвуют в регуляции различных биохимических процессов, в том числе и в регуляции биосинтеза белка.

Структура и функции РНК
Содержание РНК в клетке в несколько раз превышает содержание ДНК. Клетка содержит несколько видов РНК, отличающихся строением, локализацией и выполняемыми функциями.
Матричные РНК (мРНК)
Транспортные РНК (тРНК)
Рибосомные РНК (рРНК)
Ядерные РНК (яРНК)
Регуляторные РНК (рРНК)
МАТРИЧНЫЕ РНК
Составляют 5% всех РНК клетки. В структуре мРНК можно выделить несколько характерных участков, среди которых есть информативный – несущий информацию о структуре белка и неинформативный, выполняющий специфические функции. К неинформативным участкам относятся КЭП, 5’ – не транслируемая область (5’ – НТО), 3’ – не транслируемая область (3’ – НТО) и полиаденилатная последовательность (поли – (А)).
КЭП (англ. Сар – кепка) – особенность – наличие метилированных (минорных) азотистых оснований. Кэп защищает мРНК от разрушающих ферментов, принимает участие в биосинтезе белка, выполняя сигнальную функцию в присоединении мРНК к рибосоме. Поли –А – это полиаденилатная последовательность, насчитывает от 50 до 400 нуклеотидных остатков и имеет специфические функции: транспортирует мРНК из ядра в цитоплазму, предопределяет время жизни мРНК, защищая ее с 3’-конца от деструктирующих ферментов.
Информационный участок служит матрицей в биосинтезе белка.
У всех мРНК он начинается с тринуклеотидной последовательности АУГ – ИНИЦИИРУЮЩИЙ КОДОН.

Заканчивается одним из трех кодонов – терминирующие (стоп-сигнал) – к ним относятся тринуклеотидные последовательности УАА, УАГ, УНА (это бессмысленные кодоны, т.к. не несут информации ни об одной аминокислоте).
Вторичная структура мРНК формируется за счет спирализации полирибонуклеотидной цепи самой на себя по принципу комплементарности азотистых оснований.

Нуклеотиды являются вездесущими компонентами жизни, участвуя в большинстве биохимических реакций. АТФ является энергетической валютой клетки, производные аденина служат коферментами, циклические нуклеотиды цAMФ и цГМФ играют роль вторичных мессенджеров. Наконец, нуклеотиды — это мономерные звенья РНК и ДНК.
Пути биосинтеза нуклеотидов служат мишенью для противоопухолевых и некоторых антибактериальных препаратов, а нарушения в путях их катаболизма ловят ступни в капкан.
Строение нуклеотидов
Нуклеотиды состоят из трех компонентов: азотистого основания, пентозы и фосфата. Нуклеотид без фосфата называется нуклеозидом. Азотистые основания — это производные пурина и пиримидина (рис. 1).
Рисунок 1. | Главные азотистые основания. Некоторые названия отражают историю открытия этих соединений. Гуанин был выделен из перуанского гуано (птичьего помета), тимин — из тимуса быка, аденин — из поджелудочной железы. Урацил получил свое название от мочевой кислоты, из которой его пытались синтезировать. Б. Структурная формула нуклеотидов. В дезоксирибонуклеотидах OH-группа при 2’-углероде замещена на H.
Биосинтез пуринов
Синтез пуринов de novo сложен и энергозатратен, поэтому в физиологических условиях пул пуринов поддерживается в основном за счет путей их реутилизации (исключение составляют быстроделящиеся клетки). Всасывание нуклеотидов из пищи мало, так как почти все остатки нуклеиновых кислот катаболизируются эпителием тонкой кишки.
Аденин и гуанин образуются из общего предшественника — инозината (IMP). Пуриновое ядро постепенно собирается на остатке 5-Фосфорибозил-1-пирофосфата (PRPP). Все четыре атома азота происходят из аминокислот: два из глутамина, один из аспартата и один из глицина (рис. 2–4).
Рисунок 2. | Синтез пуриновых нуклеотидов de novo: часть 1, образование пуринового кольца инозината. После стадии ③ R обозначает 5-фосфо-D-рибозильную группу. Снизу изображено происхождение атомов пуринового кольца.
Зависимость синтеза пуринов от фолатов на стадиях 3 и 9 означает, что ингибиторы метаболизма фолиевой кислоты являются непрямыми ингибиторами синтеза пуринов, а значит, и ингибиторами деления клетки.
Рисунок 3. | Биосинтез AMP и GMP из IMP.
Регуляция этого пути сложна и мало изучена. Известно, что AMP, GMP и IMP ингибируют его по механизму отрицательной обратной связи. В недавних исследованиях [1] было показано, что шесть ферментов биосинтеза пуринов собираются в пуриносому, являющую собой новый уровень организации ферментов.
Рисунок 4. | Обзорная схема синтеза пуриновых нуклеотидов.
Реутилизация пуринов
Свободные пуриновые основания постоянно высвобождаются в клетке в процессе катаболизма нуклеотидов, и их большая часть повторно используется для синтеза нуклеотидов в более простом пути, нежели синтез de novo.
Один из таких путей настолько прост, что состоит из одной реакции, катализируемой аденозинфосфорибозилтрансферазой: Аденин + PRPP → AMP + PPi
Свободный гуанин и гипоксантин реутилизируются в похожем пути под действием гипоксантин-гуанин-фосфорибозилтрансферазы. Мутация гена вышеназванного фермента вызывает синдром Лёша — Нихена. Это сцепленное с мужским полом заболевание, манифестирующее в раннем возрасте и характеризующееся деформирующим артритом и неврологическими нарушениями. Мальчики с синдромом Лёша — Нихена плохо ориентируются в пространстве, очень враждебны и склонны к саморазрушению вплоть до откусывания себе пальцев.
Биосинтез пиримидинов
Если при синтезе пуриновых нуклеотидов азотистое основание кусочек за кусочком собирается на остатке рибозилфосфата, то при синтезе пиримидинов к рибозе оно присоединяется уже в готовом виде (рис. 5).
Рисунок 5. | Синтез пиримидиновых нуклеотидов. Стадия 1: Синтез карбамоилфосфата. 2: Конденсация аспартата и карбамоилфосфата с образованием N-карбамоиласпартата. 3: Внутримолекулярная конденсация, катализируемая дигидрооротазой с образованием шестичленного гетероцикла (дигидрооротата). 4: Окисление дигидрооротата до оротата. 5: присоединение к оротату рибозо-5-фосфата (PRPP). 6: Декарбоксилирование оротодилата до уридилата. Ферменты 5 и 6 стадий образуют комплекс и называются УМФ-синтазой.
Источник: Campbell M. K., Farell S. O. Biochemistry
Мутации гена UMPS, кодирующего УМФ-синтазу вызывают оротацидурию — редкое (описано лишь 20 случаев) аутосомно-рецессивное заболевание, проявляющееся тяжелой мегалобластной анемией, задержкой развития вследствие дефицита пиримидиновых нуклеотидов и повышенным содержанием оротовой кислоты в моче.
Нуклеозидмонофосфаты превращаются в нуклеозидтрифосфаты под действием нуклеозидмонофосфаткиназ и нуклеозиддифосфаткиназ. Последние необычны тем, что являются неспецифичными ни по основанию, ни по пентозе.
Цитидилатсинтетаза образует цитидилат из UTP.
Прежде чем описать синтез тимидилата, необходимо упомянуть об образовании дезоксирибонуклеотидов, так как его синтез de novo возможен только из дезоксирибонуклеотидного предшественника.
Образование дезоксирибонуклеотидов
Строительные блоки ДНК образуются из соответствующих рибонуклеотидов непосредственным восстановлением 2’-углеродного атома рибозы с образованием 2’-дезоксипроизводного. Реакция катализируется рибонуклеотидредуктазой, ее субстраты — нуклеозиддифосфаты, в ней также участвуют тиоредоксин и тиоредоксинредуктаза.
Рисунок 6. | Восстановление рибонуклеотидов до дезоксирибунклеотидов.
Регуляция рибонуклеотидредуктазы весьма необычна и служит для поддержания баланса четырех нуклеотидов, необходимых для синтеза ДНК. Каждая субъединица имеет 2 типа регуляторных центров: один центр регулирует активность (ATP +, dATP-), второй влияет на субстратную специфичность. Например, если со вторым центром связывается ATP или dATP, фермент начинает отдавать предпочтение восстановлению UDP или CDP, а когда UTP и СTP становится слишком много, возвращается к ADP. ATP выступает главным стимулятором восстановления всех рибонуклеотидов, а dATP в больших концентрациях выключает фермент.
Биосинтез тимидилата
Непосредственный предшественник тимидилата — dUMP. Превращение катализируется тимидилатсинтазой и сопряжено с окислением метилентетрагидрофолата до дигидрофолата (рис. 7).
Рисунок 7. | Синтез тимидилата.
Источник: Campbell M. K., Farell S. O. Biochemistry
Синтез тимидилата является мишенью некоторых противоопухолевых препаратов. Так, в химиотерапии используется ингибитор тимидилатсинтазы — фторурацил (вернее, его метаболит FdUMP), ингибитор дигидрофолатредуктазы — аминоптерин и структурный аналог фолата — метотрексат. Потребность раковых клеток в нуклеотидах гораздо выше, чем у нормальных клеток, поэтому они более чувствительны к ингибиторам биосинтеза тимидилата.
Катаболизм нуклеотидов
Катаболизм пиримидинов
С помощью нуклеотидазы и фосфорилазы от пиримидиновых нуклеотидов отщепляются фосфаты и рибоза, а оставшиеся свободные азотистые основания подвергаются дезаминированию и гидрированию (рис. 8).
Рисунок 8. | Катаболизм пиримидинов. Источник: Campbell M. K., Farell S. O. Biochemistry
Пути распада пиримидинов ведут главным образом к NH4+ и далее к мочевине.
Катаболизм пуринов
Пуриновые нуклеотиды теряют свои фосфаты под действием 5’-нуклеотидазы.
Аденилат превращается в аденозин, который дезаминируется аденозиндезаминазой (ADA) с образованием инозина, который гидролизуется нуклеозидазой до гипоксантина и рибозы. Ксантиноксидаза окисляет гипоксантин до ксантина и затем до мочевой кислоты. (рис. 9)
Рисунок 9. | Катаболизм пуриновых нуклеотидов. Общий метаболит катаболизма пуринов — ксантин. Превращение ксантина в мочевую кислоту катализирует ксантиноксидаза. В правом верхнем углу изображена аденозиндезаминаза (ADA)
У большинства млекопитающих мочевая кислота окисляется до более растворимого соединения — аллантоина, однако у человека, далматинов, приматов и птиц она является конечным экскретируемым продуктом катаболизма пуринов, и здесь время поговорить о подагре.
Заболевания, связанные с катаболизмом пуринов
1. Гиперурикемия и подагра.
Повышенное содержание мочевой кислоты в крови называется гиперурикемией (> 360 мкмоль/л для женщин и > 400 мкмоль/л для мужчин). У этого состояния может быть множество причин: дефект генов, отвечающих за экскрецию уратов почками и кишечником, богатая пуринами и/или фруктозой диета, низкоуглеводная диета, голодание, синдром лизиса опухоли, прием препаратов (диуретиков, аспирина, никотиновой кислоты и др.), синдром Лёша-Нихена, большие дозы алкоголя и другие, но всех их можно поделить на два типа: гиперпродукция уратов и снижение их экскреции. Гиперурикемия далеко не всегда сопровождается подагрой, но подагра гиперурикемией — всегда. Подагра — это метаболическое заболевание, характеризующееся отложением кристаллов уратов натрия преимущественно в плюснефаланговых суставах, поражением почек и приступами острого подагрического артрита.
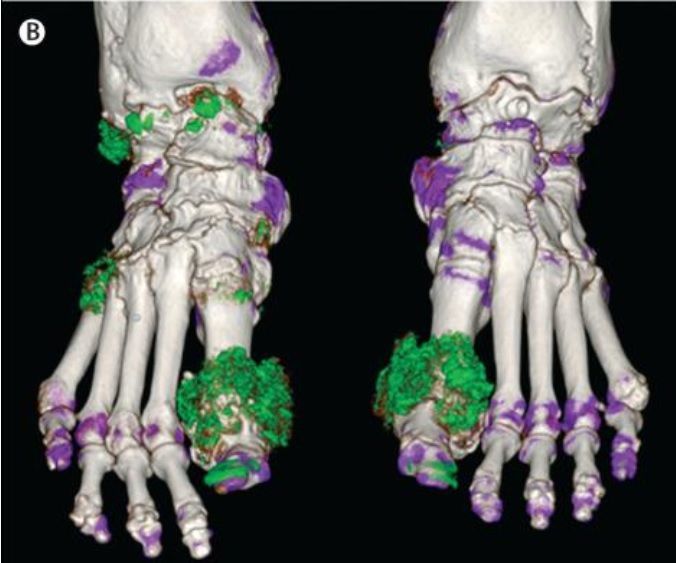
Рисунок 10. | Двухэнергетическая КТ стоп пациента с тофусной подагрой. Депозиты урата показаны зеленым цветом ( на ногтях – артефакт)
Этиология, патогенез и терапия подагры заслуживают отдельной статьи. Здесь я упомяну только о препаратах, влияющих на катаболизм пуриновых нуклеотидов, — ингибиторах ксантиноксидазы (фермента, превращающего ксантин в мочевую кислоту, см. рис. 9). Один из них — аллопуринол, структурный аналог гипоксантина, конкурентный ингибитор ксантиноксидазы. К неконкурентным ингибиторам относится фебуксостат. Оба препарата снижают уровень мочевой кислоты в крови и используются для предотвращения приступов подагры.
2. Дефицит аденозиндезаминазы ведет к одному из типов тяжёлого комбинированного иммунодефицита (SCID, синдром Глянцмана — Риникера, в 30 % случаев вызывается дефицитом ADA), характеризующемуся неспособностью T- и B- лимфоцитов к пролиферации и продукции антител.
Отсутствие аденозиндезаминазы ведет к значительному накоплению dATP, который ингибирует нуклеотидредуктазу, в результате чего возникает дефицит всех дезоксирибонуклеотидов (рис. 11). ADA является Zn-зависимым ферментом, поэтому нарушения в делении лимфоцитов возможны и при дефиците в организме цинка.
Рисунок 11. | Недостаточность аденозиндезаминазы приводит к нарушениям репликации ДНК
Наряду с аминокислотами важнейшей группой азотистых веществ являются нуклеотиды. Их биологическое значение для жизнедеятельности организмов определяется тем, что они используются для построения молекул нуклеиновых кислот - дезоксирибонуклеиновой (ДНК) и рибонук-леиновой (РНК), входят в состав каталитических центров ферментов, участвуют в биоэнергетических процессах и синтезе углеводов, липидов, белков, алкалоидов и других веществ. Некоторые нуклеотиды способны выполнять регуляторные функции.
Главные структурные компоненты нуклеотидов–азотистые основания, пентозы (рибоза или дезоксирибоза) и остаток ортофосфорной кислоты. В зависимости от углеводного компонента различают две группы нук-леотидов: рибонуклеотиды, содержащие остаток рибозы, и дезоксирибо-нуклеотиды, имеющие в своем составе остаток дезоксирибозы. Дезоксирибонуклеотиды используются организмами для синтеза ДНК, а рибонуклетиды входят в состав РНК, ферментов и макроэргических нук-леозидполифосфатов.
Рибоза и дезоксирибоза в составе нуклеотидов находятся в b-D-фура-нозной форме:

Нуклеотиды образуются из двух типов азотистых оснований – произ-водных пиримидина и пурина. Свойства оснований они проявляют в водном растворе при взаимодействии с молекулами воды. Из пиримидиновых осно-ваний наиболее важное значение имеют урацил, тимин и цитозин как основные структурные единицы нуклеотидов, образующих нуклеиновые кислоты. Кроме них, известны и другие основания - 5-метилцитозин, псевдоурацил, 5-оксиметилцитозин и др. 5-Метилцитозин и 5-оксиметилцитозин в небольшом количестве могут
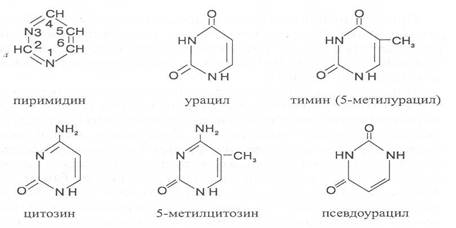
содержаться в составе нуклеотидов ДНК и РНК, псевдоурацил - в транспортной РНК.
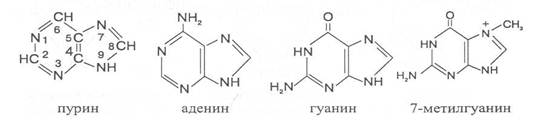
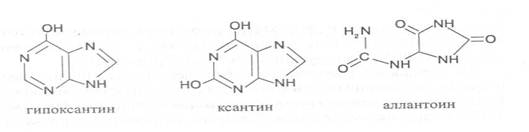
Из пуриновых оснований наибольшее значение имеют аденин и гуанин, так как они используются для синтеза нуклеиновых кислот. В составе нуклеиновых кислот в небольшом количестве обнаружены также и другие основания, которые образуются в результате химической модификации аденина и гуанина: 7-метилгуанин, 2-метиладенин, N-диметилгуанин и др. Важными промежуточными метаболитами являются гипоксантин, ксантин, аллантоин. В некоторых растениях они могут накапливаться в свободном состоянии.
Все азотистые основания интенсивно поглощают ультрафиолетовый свет при длинах волн 200-280нм.
При соединении азотистых оснований с молекулой рибозы или дезоксирибозы образуются соединения, называемые нуклеозидами, так как между пентозой и основанием возникает гликозидная связь. Основания в данном случае можно рассматривать как агликоны по отношению к пентозе.
В нуклеозидах гликозидная связь возникает между первым углеродным атомом пентозы в b-фуранозной форме и азотом пуринового (в девятом положении) или пиримидинового (в первом положении) основания. Азотистые основания аденин, гуанин, цитозин и урацил образуют при со-единении с рибозой нуклеозиды - аденозин, гуанозин, цитидин и уридин,
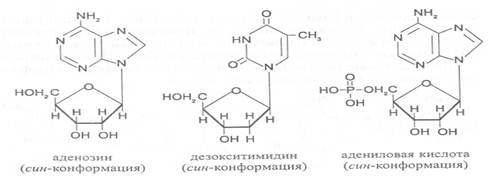
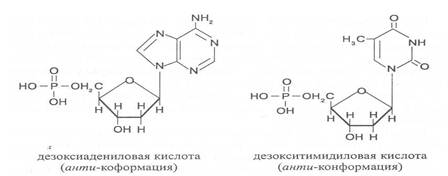
а с дезоксирибозой – дезоксиаденозин, дезоксигуанозин, дезоксицитидин, дезоксиуридин. Тимин, соединяясь с дезоксирибозой, даёт дезоксити-мидин.
Азотистые основания и нуклеозиды могут накапливаться в растениях в значительном количестве при интенсивном распаде нуклеиновых кислот.
Фосфорнокислые эфиры нуклеозидов называют нуклеотидами. В составе нуклеотидов остатки ортофосфорной кислоты могут присоединяться к пятому или третьему атомам углерода рибозы или дезоксирибозы, а у некоторых рибонуклеотидов ещё и ко второму атому углерода рибозы. У свободных нуклеотидов фосфатная группа обычно находится у пятого углеродного атома рибозы или дезоксирибозы. В нейтральной среде остатки ортофосфорной кислоты в молекулах нуклеотидов сильно диссоциированы, вследствие чего могут присоединять катионы, поэтому при химическом выделении нуклеотиды кристаллизуются в виде солей.
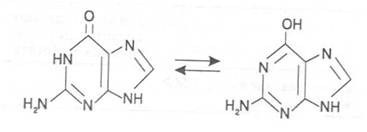
Изучение пространственной структуры азотистых оснований методом рентгеноструктурного анализа показывает, что все они имеют почти плоскую конформацию. У них довольно легко происходит перегруппировка двойных связей, которая сопровождается таутомерными превращениями. Например, гуанин может существовать в виде двух таутомерных форм:
Плоскость гетероциклического ядра основания в структуре нуклеозидов и нуклеотидов может занимать в пространстве два положения по отношению к пентозе, образуя две противоположные конформации - син-конформацию и анти-конформацию. В анти-конформации структура азотистого основания развернута от пентозы, а в син-конформации ориентирована над её плоскостью. В свободном состоянии пиримидиновые нуклеотиды находятся преимущественно в анти-конформации, а пуриновые довольно легко переходят из одной формы в другую.

В связи с тем, что у нуклеотидов сильно выражены кислотные свойства, их называют кислотами с учетом названий азотистых оснований и углеводного компонента. Так, например, рибонуклеотид, имеющий остаток аденина, называют адениловой кислотой, или аденозинмонофосфатом (АМФ). Дезоксирибонуклеотид, образованный из тимина, называют дезокситимидиловой кислотой, или дезокситимидинмонофосфатом (дТМФ). Названия других нуклеотидов представлены в таблице 2.
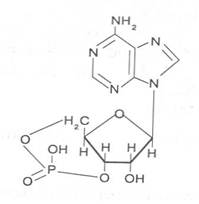
В растениях найдены циклические формы нуклеотидов – адено-зинмонофосфата и гуанозинмонофосфата, которые по-видимому выполняют регуляторные функции. Строение циклического АМФ можно представить слудующей формулой:
2. Названия важнейших нуклеотидов.
| Азотистые основания | Рибонуклеотиды | Сокращён- ное обо- значение | Дезоксирибонук- Леотиды | Сокращён- ное обо- значение |
| Аденин | Адениловая кислота, аде-нозинмонофосфат | АМФ | дезоксиаденило- вая кислота, дезоксиадено- зинмонофосфат | дАМФ |
| Гуанин | Гуаниловая кислота, гуа- нозинмонофосфат | ГМФ | дезоксигуанило- вая кислота, дезоксигуано- зинмонофосфат | ДГМФ |
| Цитозин | Цитидиловая кислота,цити-динмонофосфат | ЦМФ | дезоксицитиди- ловая кислота, дезоксицитидин- монофосфат | ДЦМФ |
| Урацил | Уридиловая кислота, ури- динмонофосфат | УМФ | ||
| Тимин | дезокситимиди- ловая кислота, дезокситимидин- монофосфат | ДТМФ | ||
| Гипоксан- тин | инозиновая кислота, ино- зинмонофосфат | ИМФ |
Путём фосфорилирования нуклеотиды превращаются в более активные формы - нуклезиддифосфаты и нуклеозидтрифосфаты, в виде которых, как мы увидим далее, они участвуют в различных биохимических процессах – синтезе нуклеиновых кислот, сложных углеводов и липидов, коферментов и витаминов, запасании и переносе энергии.
Из адениловой кислоты образуются аденозинди- и аденозинтрифосфат (АДФ и АТФ), из гуаниловой кислоты - гуанозинди- и гуанозинтрифосфат (ГДФ и ГТФ), из цитидиловой кислоты - цитидинди- и цитидинтрифосфат (ЦДФ и ЦТФ), из уридиловой кислоты - уридинди- и уридинтрифосфат (УДФ и УТФ), из инозиновой кислоты – инозинди- и инозинтрифосфат (ИДФ и ИТФ). Из дезоксирибонуклеотидов также синтезируются соответствующие ди- и трифосфаты: дезоксиаденозинди- и дезоксиаденозинтрифосфат (дАДФ и дАТФ), дезоксигуанозинди- и дезоксигуанозинтрифосфат (дГДФ и дГТФ), дезоксицитидинди- и дезоксицитидинтрифосфат (дЦДФ и дЦТФ), де-зокситимидинди- и дезокситимидинтрифосфат (дТДФ и дТТФ). Строение ди- и трифосфатов нуклеозидов можно представить следующими формулами:
Свободные нуклеотиды содержатся во всех растительных клетках. Они возникают или в процессе активного синтеза азотистых веществ, или в результате распада нуклеиновых кислот, который может интенсивно происходить при неблагоприятных условиях произрастания, а также при хранении и переработке растительных продуктов. Так, например, при разваривании клубней картофеля образующиеся в результате распада нуклеиновых кислот свободные нуклеотиды оказывают влияние на формирование вкусовых качеств варёного картофеля.
По интенсивности образования различных фракций нуклеотидов можно судить о направленности биохимических процессов в организме. Если в клетках и тканях организма повышается содержание трифосфатпроизводных нуклеотидов, то это свидетельствует об активизации его жизнедеятельности, тогда как накопление монофосфатов наблюдается при усилении процессов распада веществ, характерных для стареющего организма.
Наряду с аминокислотами важнейшей группой азотистых веществ являются нуклеотиды. Их биологическое значение для жизнедеятельности организмов определяется тем, что они используются для построения молекул нуклеиновых кислот - дезоксирибонуклеиновой (ДНК) и рибонук-леиновой (РНК), входят в состав каталитических центров ферментов, участвуют в биоэнергетических процессах и синтезе углеводов, липидов, белков, алкалоидов и других веществ. Некоторые нуклеотиды способны выполнять регуляторные функции.
Главные структурные компоненты нуклеотидов–азотистые основания, пентозы (рибоза или дезоксирибоза) и остаток ортофосфорной кислоты. В зависимости от углеводного компонента различают две группы нук-леотидов: рибонуклеотиды, содержащие остаток рибозы, и дезоксирибо-нуклеотиды, имеющие в своем составе остаток дезоксирибозы. Дезоксирибонуклеотиды используются организмами для синтеза ДНК, а рибонуклетиды входят в состав РНК, ферментов и макроэргических нук-леозидполифосфатов.
Рибоза и дезоксирибоза в составе нуклеотидов находятся в b-D-фура-нозной форме:
Нуклеотиды образуются из двух типов азотистых оснований – произ-водных пиримидина и пурина. Свойства оснований они проявляют в водном растворе при взаимодействии с молекулами воды. Из пиримидиновых осно-ваний наиболее важное значение имеют урацил, тимин и цитозин как основные структурные единицы нуклеотидов, образующих нуклеиновые кислоты. Кроме них, известны и другие основания - 5-метилцитозин, псевдоурацил, 5-оксиметилцитозин и др. 5-Метилцитозин и 5-оксиметилцитозин в небольшом количестве могут
содержаться в составе нуклеотидов ДНК и РНК, псевдоурацил - в транспортной РНК.
Из пуриновых оснований наибольшее значение имеют аденин и гуанин, так как они используются для синтеза нуклеиновых кислот. В составе нуклеиновых кислот в небольшом количестве обнаружены также и другие основания, которые образуются в результате химической модификации аденина и гуанина: 7-метилгуанин, 2-метиладенин, N-диметилгуанин и др. Важными промежуточными метаболитами являются гипоксантин, ксантин, аллантоин. В некоторых растениях они могут накапливаться в свободном состоянии.
Все азотистые основания интенсивно поглощают ультрафиолетовый свет при длинах волн 200-280нм.
При соединении азотистых оснований с молекулой рибозы или дезоксирибозы образуются соединения, называемые нуклеозидами, так как между пентозой и основанием возникает гликозидная связь. Основания в данном случае можно рассматривать как агликоны по отношению к пентозе.
В нуклеозидах гликозидная связь возникает между первым углеродным атомом пентозы в b-фуранозной форме и азотом пуринового (в девятом положении) или пиримидинового (в первом положении) основания. Азотистые основания аденин, гуанин, цитозин и урацил образуют при со-единении с рибозой нуклеозиды - аденозин, гуанозин, цитидин и уридин,
а с дезоксирибозой – дезоксиаденозин, дезоксигуанозин, дезоксицитидин, дезоксиуридин. Тимин, соединяясь с дезоксирибозой, даёт дезоксити-мидин.
Азотистые основания и нуклеозиды могут накапливаться в растениях в значительном количестве при интенсивном распаде нуклеиновых кислот.
Фосфорнокислые эфиры нуклеозидов называют нуклеотидами. В составе нуклеотидов остатки ортофосфорной кислоты могут присоединяться к пятому или третьему атомам углерода рибозы или дезоксирибозы, а у некоторых рибонуклеотидов ещё и ко второму атому углерода рибозы. У свободных нуклеотидов фосфатная группа обычно находится у пятого углеродного атома рибозы или дезоксирибозы. В нейтральной среде остатки ортофосфорной кислоты в молекулах нуклеотидов сильно диссоциированы, вследствие чего могут присоединять катионы, поэтому при химическом выделении нуклеотиды кристаллизуются в виде солей.
Изучение пространственной структуры азотистых оснований методом рентгеноструктурного анализа показывает, что все они имеют почти плоскую конформацию. У них довольно легко происходит перегруппировка двойных связей, которая сопровождается таутомерными превращениями. Например, гуанин может существовать в виде двух таутомерных форм:
Плоскость гетероциклического ядра основания в структуре нуклеозидов и нуклеотидов может занимать в пространстве два положения по отношению к пентозе, образуя две противоположные конформации - син-конформацию и анти-конформацию. В анти-конформации структура азотистого основания развернута от пентозы, а в син-конформации ориентирована над её плоскостью. В свободном состоянии пиримидиновые нуклеотиды находятся преимущественно в анти-конформации, а пуриновые довольно легко переходят из одной формы в другую.
В связи с тем, что у нуклеотидов сильно выражены кислотные свойства, их называют кислотами с учетом названий азотистых оснований и углеводного компонента. Так, например, рибонуклеотид, имеющий остаток аденина, называют адениловой кислотой, или аденозинмонофосфатом (АМФ). Дезоксирибонуклеотид, образованный из тимина, называют дезокситимидиловой кислотой, или дезокситимидинмонофосфатом (дТМФ). Названия других нуклеотидов представлены в таблице 2.
В растениях найдены циклические формы нуклеотидов – адено-зинмонофосфата и гуанозинмонофосфата, которые по-видимому выполняют регуляторные функции. Строение циклического АМФ можно представить слудующей формулой:
2. Названия важнейших нуклеотидов.
| Азотистые основания | Рибонуклеотиды | Сокращён- ное обо- значение | Дезоксирибонук- Леотиды | Сокращён- ное обо- значение |
| Аденин | Адениловая кислота, аде-нозинмонофосфат | АМФ | дезоксиаденило- вая кислота, дезоксиадено- зинмонофосфат | дАМФ |
| Гуанин | Гуаниловая кислота, гуа- нозинмонофосфат | ГМФ | дезоксигуанило- вая кислота, дезоксигуано- зинмонофосфат | ДГМФ |
| Цитозин | Цитидиловая кислота,цити-динмонофосфат | ЦМФ | дезоксицитиди- ловая кислота, дезоксицитидин- монофосфат | ДЦМФ |
| Урацил | Уридиловая кислота, ури- динмонофосфат | УМФ | ||
| Тимин | дезокситимиди- ловая кислота, дезокситимидин- монофосфат | ДТМФ | ||
| Гипоксан- тин | инозиновая кислота, ино- зинмонофосфат | ИМФ |
Путём фосфорилирования нуклеотиды превращаются в более активные формы - нуклезиддифосфаты и нуклеозидтрифосфаты, в виде которых, как мы увидим далее, они участвуют в различных биохимических процессах – синтезе нуклеиновых кислот, сложных углеводов и липидов, коферментов и витаминов, запасании и переносе энергии.
Из адениловой кислоты образуются аденозинди- и аденозинтрифосфат (АДФ и АТФ), из гуаниловой кислоты - гуанозинди- и гуанозинтрифосфат (ГДФ и ГТФ), из цитидиловой кислоты - цитидинди- и цитидинтрифосфат (ЦДФ и ЦТФ), из уридиловой кислоты - уридинди- и уридинтрифосфат (УДФ и УТФ), из инозиновой кислоты – инозинди- и инозинтрифосфат (ИДФ и ИТФ). Из дезоксирибонуклеотидов также синтезируются соответствующие ди- и трифосфаты: дезоксиаденозинди- и дезоксиаденозинтрифосфат (дАДФ и дАТФ), дезоксигуанозинди- и дезоксигуанозинтрифосфат (дГДФ и дГТФ), дезоксицитидинди- и дезоксицитидинтрифосфат (дЦДФ и дЦТФ), де-зокситимидинди- и дезокситимидинтрифосфат (дТДФ и дТТФ). Строение ди- и трифосфатов нуклеозидов можно представить следующими формулами:
Свободные нуклеотиды содержатся во всех растительных клетках. Они возникают или в процессе активного синтеза азотистых веществ, или в результате распада нуклеиновых кислот, который может интенсивно происходить при неблагоприятных условиях произрастания, а также при хранении и переработке растительных продуктов. Так, например, при разваривании клубней картофеля образующиеся в результате распада нуклеиновых кислот свободные нуклеотиды оказывают влияние на формирование вкусовых качеств варёного картофеля.
По интенсивности образования различных фракций нуклеотидов можно судить о направленности биохимических процессов в организме. Если в клетках и тканях организма повышается содержание трифосфатпроизводных нуклеотидов, то это свидетельствует об активизации его жизнедеятельности, тогда как накопление монофосфатов наблюдается при усилении процессов распада веществ, характерных для стареющего организма.
Нуклеоти́ды — фосфорные эфиры нуклеозидов, нуклеозидфосфаты. Свободные нуклеотиды, в частности АТФ, цАМФ, АДФ, играют важную роль в энергетических и информационных внутриклеточных процессах, а также являются составляющими частями нуклеиновых кислот и многих коферментов.
Содержание
Строение
Нуклеотиды являются сложными эфирами нуклеозидов и фосфорных кислот. Нуклеозиды, в свою очередь, являются N-гликозидами, содержащими гетероциклический фрагмент, связанный через атом азота с C-1 атомом остатка сахара.

В природе наиболее распространены нуклеотиды, являющиеся β-N-гликозидами пуринов или пиримидинов и пентоз — D-рибозы или D-2-дезоксирибозы. В зависимости от структуры пентозы различают рибонуклеотиды и дезоксирибонуклеотиды, которые являются мономерами молекул сложных биологических полимеров (полинуклеотидов) — соответственно РНК или ДНК. [1]
Фосфатный остаток в нуклеотидах обычно образует сложноэфирную связь с 2'-, 3'- или 5'-гидроксильными группами рибонуклеозидов, в случае 2'-дезоксинуклеозидов этерифицируются 3'- или 5'-гидроксильные группы.
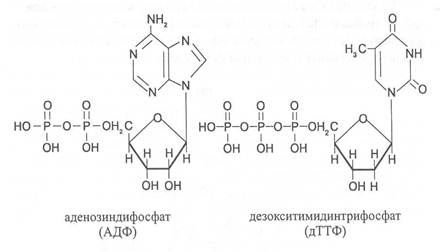
Большинство нуклеотидов являются моноэфирами ортофосфорной кислоты, однако известны и диэфиры нуклеотидов, в которых этерифицированы два гидроксильных остатка — например, циклические нуклеотиды циклоаденин- и циклогуанин монофосфаты (цАМФ и цГМФ). Наряду с нуклеотидами — эфирами ортофосфорной кислоты (монофосфатами) в природе также распространены и моно- и диэфиры пирофосфорной кислоты (дифосфаты, например, аденозиндифосфат) и моноэфиры триполифосфорной кислоты (трифосфаты, например, аденозинтрифосфат).
Номенклатура
| Код | Означает | Комплементарная пара |
|---|---|---|
| A | A | T в ДНК; U в РНК |
| C | C | G |
| G | G | C |
| T или U | T в ДНК; U в РНК | A |
| M | A или C | K |
| R | A или G | Y |
| W | A или T | W |
| S | C или G | S |
| Y | C или T | R |
| K | G или T | M |
| V | A или C или G | B |
| H | A или C или T | D |
| D | A или G или T | H |
| B | C или G или T | V |
| X или N | A или C или G или T (U) | любой |
Соединения, состоящие из двух нуклеотидовых молекул, называются динуклеотидами, из трёх — тринуклеотидами, из небольшого числа — олигонуклеотидами, а из многих — полинуклеотидами, или нуклеиновыми кислотами.
Названия нуклеотидов представляют собой аббревиатуры в виде стандартных трёх- или четырёхбуквенных кодов.
Латинские и русские коды для нуклеиновых оснований:
- A — А: Аденин;
- G — Г: Гуанин;
- C — Ц: Цитозин;
- T — Т: Тимин (5-метилурацил), не встречается в РНК, занимает место урацила в ДНК;
- U — У: Урацил, не встречается в ДНК, занимает место тимина в РНК.
Общепринятые буквенные коды для обозначения нуклеотидных оснований соответствуют номенклатуре, принятой Международным обществом теоретической и прикладной химии (International Union of Pure and Applied Chemistry, сокращённо — англ. IUPAC , рус. ИЮПАК ) и Международным обществом биохимии и молекулярной биологии (International Union of Biochemistry and Molecular Biology, сокращённо — англ. IUBMB ). Если при секвенировании последовательности ДНК или РНК возникает сомнение в точности определения того или иного нуклеотида, помимо пяти основных (A, C, T, G, U), используют другие буквы латинского алфавита в зависимости от того, какие наиболее вероятные нуклеотиды могут находиться в данной позиции последовательности.
Длину секвенированных участков ДНК (гена, сайта, хромосомы) или всего генома указывают в парах нуклеотидов (пн), или парах оснований (англ. base pairs , сокращённо bp), подразумевая под этим элементарную единицу двухцепочечной молекулы нуклеиновой кислоты, сложенную из двух спаренных комплементарных оснований.
3. Выступают в роли коферментов (ФАД, ФМН, НАД + , НАДФ + )
4. Циклические мононуклеотиды являются вторичными посредниками при действии гормонов и других сигналов(цАМФ, цГМФ).
5. Аллостерические регуляторы активности ферментов.
6. Являются мономерами в составе нуклеиновых кислот, связанные 3'-5'- фосфодиэфирными связями.
История
В домолекулярной генетике для обозначения наименьшего элемента в структуре ДНК, который может быть подвержен спонтанной или индуцированной мутации, применялся особый термин рекон. В настоящее время показано, что таким наименьшим элементом является один нуклеотид (или одно азотистое основание в составе нуклеотида), поэтому данный термин более не употребляется. Для определения понятия единица мутации применялся термин мутон. В настоящее время показано, что фенотипически мутация может проявиться даже при замене одного нуклеотида (или азотистого основания в составе нуклеотида), таким образом, термин мутон соответствует одному нуклеотиду.
Читайте также: