
Процессы способствующие переносу генетической информации кратко
Обновлено: 02.07.2024
Концепция генетического кода имела принципиальное значение для генетики, поскольку постулировала существование в природе определенной системы передачи информации. Пути переноса информации легли в основу представлений, которые Ф. Крик назвал центральной догмой молекулярной биологии. Основной порядок переписывания генетической информации в живой природе стал выражаться формулой:
ДНК → РНК → белок.
Такой поток информации реализуется в подавляющем большинстве живых систем. Единство генетического кода и общность потока генетической информации в природе показали, что эти явления произошли на самом раннем этапе эволюции жизни. Однако оба этих фундаментальных процесса имеют свои исключения. У некоторых вирусов были обнаружены особые варианты переноса информации – от РНК к РНК и от РНК к ДНК.
Огромное теоретическое значение имело открытие обратной транскрипции, вызвавшее широкий резонанс в научных кругах. Это открытие резко меняло устоявшиеся в генетике представления. При обратной транскрипции на РНК-матрице синтезируется двунитиевая структура ДНК (сначала одна нить, затем – другая). Этот процесс катализируется ферментом обратной транскриптазой (или РНК-зависимой-ДНК-полимеразой). Обратная транскриптаза была открыта в 1970 г. при исследовании так называемых ретровирусов. Синтезированная ими молекула ДНК встраивается в хромосому клетки-хозяина и при делении передается дочерним клеткам. В такой форме геном вируса может существовать долгое время, переходя от родителей к потомкам. Скрытая интегрированная форма вируса получила название провирус. При активации провируса происходит воспроизведение РНК ретровируса. Эта РНК может транслироваться, синтезируя вирусные белки (в том числе и обратную транскриптазу), и образовывать новые вирусные частицы, покрытые оболочкой.
Явление обратной транскрипции оказалось отнюдь не экзотикой. В настоящее время ретровирусам отводится определенная роль в эволюционном процессе как переносчикам генетической информации между организмами разных систематических групп. При образовании новых вирусных частиц активированного провируса может захватываться часть генетического материала клетки-хозяина и передаваться другим организмам, минуя межвидовые барьеры. Эволюционная роль ретровирусов будет рассмотрена ниже.
Таким образом, с учетом явления обратной транскрипции, общая схема передачи информации в природе стала представляться следующим образом:
Сенсационным стал результат анализа прионных болезней. Он оказался столь неожиданным, что поставил под сомнение незыблемость центральной догмы биологии.
Белок-прион обозначается как PrPSc. Он гомологичен обычному клеточному белку (255 аминокислотных остатков) – PrPс, находящемуся в клетках нервной системы, некоторых тканей и лимфоцитах. PrPSc и PrPс имеют одинаковую первичную структуру, но различаются по вторичной и третичной структурам. Они кодируются геном, весьма похожим у всех млекопитающих. У человека этот ген расположен на 20-й хромосоме. Он характеризуется выраженным полиморфизмом: в настоящее время у человека обнаружено 15 его вариантов.
Прионы образуются путем посттрансляционной модификации нормального клеточного белка PrPс. Благодаря такой модификации прионы приобретают свойства инфекционности. Молекула PrPс, сталкиваясь с молекулой PrPSc, меняет свою конфигурацию и сама становится PrPSc.
Инфекционность прионов, т. е. воспроизводимость их модифицированной структуры даже в другом организме, является самой большой загадкой. Единичный прион превращает все вновь синтезированные полипептиды клетки с близкой ему первичной структурой в свое подобие. Таким образом, белки-прионы выступают в роли матрицы, вызывая изменение вторичной и третичной структуры(рефолдинг) клеточного белка PrPс.
Возможным объяснением этого явления может служить гипотеза конформационных матриц – наличие в клетке двух категорий матричных процессов: для последовательностей мономеров и для конформации молекул. Такой взгляд заставляет еще раз пересмотреть центральную догму биологии и внести в нее необходимые изменения.
Однако в молекулярной природе прионов еще очень много неясного, поэтому следует остерегаться скоропалительных выводов. Прионы в настоящее время интенсивно изучаются. Причиной столь пристального внимания служит не только большой теоретический интерес, но и неизлечимость на сегодняшний день прионных болезней.
Взаимодействие цепей ДНК может быть нарушено при нагревании, приводящем к денатурации ДНК. Денатурация сопровождается увеличением поглощения при 260 нм (так называемый гиперхромный эффект). Помимо этого метода процесс денатурации можно контролировать по уменьшению вязкости раствора. При медленном остывании (отжиге) раствора ДНК, подвергнутой тепловой денатурации, две комплементарные цепи ДНК вновь соединяются с образованием исходной структуры, т.е. происходит ренатурация. На явлении денатурации и ренатурации основан метод молекулярной гибридизации -– один из главных в генной инженерии.
Гибридизация днк-днк
Если смешать растворы ДНК, выделенных из организмов разных видов, нагреть эту смесь, а затем вновь охладить, то вновь будут возникать двуспиральные структуры. При этом наряду с молекулами ДНК, идентичными исходным, могут образовываться гибридные молекулы, содержащие цепи ДНК от разных видов. Такие гибридные молекулы несовершенны: спирализованные участки в них чередуются с неспирализованными. В неспирализованных участках цепи ДНК некомплементарны друг другу. Несовершенство гибридов ДНК-ДНК выявляется с помощью электронной микроскопии.
Исследование гибридизации ДНК-ДНК позволило сделать следующие важные для биологии выводы:
1) ДНК всех органов и тканей одного и того же организма идентичны.
2) ДНК, выделенные из тканей разных особей одного биологического вида, идентичны. Небольшие различия, имеющие место, методом гибридизации не обнаруживаются.
3)ДНК, полученные от особей разных биологических видов, неидентичны и образуют несовершенные гибридные молекулы. Степень несовершенства гидридов ДНК-ДНК тем больше, чем отдаленнее филогенетическое сродство между видами.
Из результатов изучения ДНК методом гибридизации следует, что первичная структура ДНК характеризуется видовой специфичностью.
Гибридизация днк-рнк
Сходным образом происходит гибридизация ДНК-РНК. В этом случае гибридная молекула содержит одну цепь ДНК и одну – РНК.
Перенос генетической информации
Способность к переносу генетической информации, т.е. передаче наследственных свойств, является уникальным свойством живых организмов. Хранение и передача генетической информации, которую можно рассматривать как биологическую память, возложена природой на нуклеиновые кислоты.
Различают три варианта переноса генетической информации: 1) в пределах одного класса нуклеиновых кислот, т.е. от ДНК к ДНК. Он называется репликацией. 2) между разными классами нуклеиновых кислот, т.е. от ДНК к РНК. Он называется транскрипцией. В ходе транскрипции образуются все типы РНК: мРНК, тРНК и рРНК. 3) в пределах разных классов макромолекул, т.е. от мРНК к белку. Он называется трансляцией. Трансляция может происходить только в одном направлении – от мРНК к белку.
Направление переноса генетической информации от ДНК через РНК к белку называется центральным постулатом молекулярной биологии. Он был сформулирован Ф.Криком.
Все виды передачи генетической информации основаны на матричном механизме. Это означает, что для каждого из них необходима матрица, которая позволяет с большой точностью и экономичностью воспроизводить имеющуюся в клетке генетическую информацию.
Биосинтез ДНК (репликация)
Репликация – это процесс, при котором информация, закодированная в последовательности оснований молекулы родительской ДНК, передается дочерней ДНК.
В 1958 году Меселсон и Сталь в экспериментах с тяжелым азотом (N 15 ) доказали, что репликация ДНК происходит по полуконсервативному механизму, при котором дочерние клетки первого поколения получают одну цепь ДНК от родителей, а вторая цепь является вновь синтезированной.
Для репликации ДНК необходим ряд условий:
1) Реакция идет только в присутствии уже готовой ДНК, выполняющей роль матрицы. Вновь синтезируемые молекулы ДНК имеют первичную структуру, идентичную первичной структуре ДНК-матрицы.
2) Сложный набор белков и ферментов, образующих репликативный комплекс. В него входят ДНК-топоизомеразы, которая является обратимой нуклеазой; ДНК-хеликазы, использующей энергию АТФ для расплетения двойной спирали ДНК; белков, дестабилизирующих спираль (SSB-белков); ДНК-полимераз, катализирующих образование 3’, 5’-фосфодиэфирных связей.
3) Субстратами служат дезоксирибонуклеозидтрифосфаты (дАТФ, дГТФ, дТТФ, дЦТФ). В ходе ДНК-полимеразной реакции от каждого из них отщепляется пирофосфатный остаток. Таким образом, включение каждого мономера в молекулу ДНК требует расхода энергии высокоэнергетических связей.
Итак, в результате действия сложного набора белков образуется репликативная вилка. ДНК-полимераза δ не способна инициировать синтез новых цепей ДНК. Она может лишь удлинять уже имеющуюся нуклеотидную цепь – затравку РНК (праймер). Роль завтравки выполняет РНК, синтезируемая ферментом РНК-полимеразой α. Каждый праймер состоит примерно из 10 нуклеотидов. ДНК-полимераза δ (дельта), активируемая праймером, продолжает синтез новой цепи в направлении от 5’- к 3’-концу. Поэтому на одной из ветвей репликативной вилки новая цепь наращивается непрерывно; она называется лидирующей. На другой же ветви по мере раскручивания ДНК под действием ДНК-полимеразы έ (эпсилон) образуются короткие фрагменты, называемые фрагментами Оказаки. Каждый фрагмент Оказаки состоит примерно из 100 нуклеотидов. Исключение праймеров осуществляется с помощь фермента ДНК-полимеразы β, которая постепенно отрезает от 5’-конца фрагмента по одному рибонуклеотиду и присоединяет дезоксирибонуклеотиды в количестве, равном вырезанному праймеру, заполняя образованную брешь. Затем концы этих фрагментов соединяются между собой под действием ДНК-лигазы.
Молекула ДНК человека имеет очень большие размеры. Репликация такой большой молекулы шла бы в течение примерно 800 часов. Поэтому инициация синтеза ДНК происходит в нескольких точках хромосомы, которые называются точками инициации репликации, или ориджинами репликации.
Центральная догма молекулярной биологии — обобщающее наблюдаемое в природе правило реализации генетической информации: информация передаётся от нуклеиновых кислот к белку, но не в обратном направлении. Правило было сформулировано Френсисом Криком в 1958 году [1] и приведено в соответствие с накопившимися к тому времени данными в 1970 году [2] . Переход генетической информации от ДНК к РНК и от РНК к белку является универсальным для всех без исключения клеточных организмов, лежит в основе биосинтеза макромолекул. Репликации генома соответствует информационный переход ДНК->ДНК. В природе встречаются также переходы РНК->РНК и РНК->ДНК (например у некоторых вирусов), а также изменение конформации белков, передаваемое от молекулы к молекуле.
Содержание
Информация, содержащаяся в биологических последовательностях
Биополимеры — это синтезируемые живыми существами (биологические) полимеры. ДНК, РНК и белки относятся к линейным полимерам, то есть каждый входящий в их состав мономер соединяется с минимум двумя другими мономерами. Последовательность мономеров кодирует информацию, правила передачи которой описываются центральной догмой. Информация передаётся с высокой точностью, детерминистически и один биополимер используется как шаблон для сборки другого полимера с последовательностью, которая полностью определяется последовательностью первого полимера.
Универсальные способы передачи биологической информации
| Общие | Специальные | Неизвестные |
|---|---|---|
| ДНК → ДНК | РНК → ДНК | белок → ДНК |
| ДНК → РНК | РНК → РНК | белок → РНК |
| РНК → белок | ДНК → белок | белок → белок |
В живых организмах встречаются три вида гетерогенных, то есть состоящих из разных мономеров полимера — ДНК, РНК и белок. Передача информации между ними может осуществляться 3 х 3 = 9 способами. Центральная догма разделяет эти 9 типов передачи информации на три группы:
- Общий — встречающиеся у большинства живых организмов;
- Специальный — встречающиеся в виде исключения, у вирусов и у мобильных элементов генома или в условиях биологического эксперимента;
- Неизвестные — не обнаружены.
Репликация ДНК (ДНК → ДНК)
ДНК — основной способ передачи информации между поколениями живых организмов, поэтому точное удвоение (репликация) ДНК очень важна. Репликация осуществляется комплексом белков, которые расплетают хроматин, затем двойную спираль. После этого ДНК полимераза и ассоциированные с ней белки, строят на каждой из двух цепочек идентичную копию.
Транскрипция (ДНК → РНК)
Транскрипция — биологический процесс, в результате которого информация, содержащаяся в участке ДНК, копируется на синтезируемую молекулу информационной РНК. Транскрипцию осуществляют факторы транскрипции и РНК-полимераза. В эукариотической клетке первичный транскрипт (пре-иРНК) часто редактируется. Этот процесс называется сплайсингом.
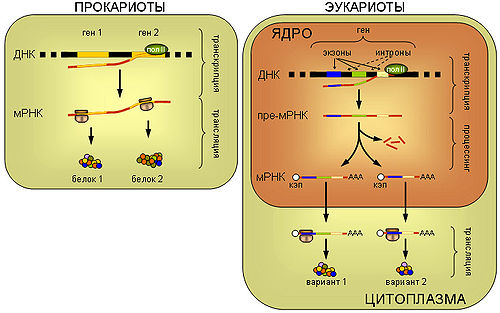
Принципиальная схема реализации генетической информации у про- и эукариот.
ПРОКАРИОТЫ. У прокариот синтез белка рибосомой (трансляция) пространственно не отделен от транскрипции и может происходить еще до завершения синтеза мРНК РНК-полимеразой. Прокариотические мРНК часто полицистронные, то есть содержат несколько независимых генов.
ЭУКАРИОТЫ. мРНК эукариот синтезируется в виде предшественника, пре-мРНК, претерпевающего затем сложное стадийное созревание — процессинг, включающий присоединение кэп-структуры к 5'-концу молекулы, присоединение нескольких десятков остатков аденина к ее 3'-концу (полиаденилирование), выщепление незначащих участков — интронов и соединение друг с другом значащих участков — экзонов (сплайсинг). При этом соединение экзонов одной и той же пре-мРНК может проходить разными способами, приводя к образованию разных зрелых мРНК, и в конечном итоге разных вариантов белка (альтернативный сплайсинг). Только мРНК, успешно прошедшая процессинг, экспортируется из ядра в цитоплазму и вовлекается в трансляцию.
Трансляция (РНК → белок)
Обратная транскрипция (РНК — ДНК)
Обратная транскрипция — перенос информации с РНК на ДНК, процесс, обратный нормальной транскрипции, осуществляемый ферментом обратной транскриптазой . Встречается у ретровирусов, например, ВИЧ и в случае ретротранспозонов.
Репликация РНК (РНК — РНК)
Репликация РНК — копирование цепи РНК на комплемлементарную ей цепь РНК с помощью фермента РНК-зависимой РНК-полимеразы. Вирусы, содержащие одноцепочечную (например, пикорнавирусы, к которым относится вирус ящура) или двуцепочечную РНК реплицируются подобным способом.
Прямая трансляция белка на матрице ДНК (ДНК — белок)
Прямая трансляция была продемонстрирована в клеточных экстрактах кишечной палочки, которые содержали рибосомы, но не иРНК. Такие экстракты синтезировали белки с введённых в систему ДНК, и антибиотик неомицин усиливал этот эффект. [3] [4]
Эпигенетические изменения
Эпигенетические изменения — это изменения в проявлении генов, не обусловленные изменением генетической информации (мутациями). Эпигенетические изменения происходят в результате модификации уровня экспрессии генов, то есть их транскрипции и/или трансляции. Наиболее изученным видом эпигенетической регуляции является метилирование ДНК с помощью белков ДНК-метилтрансфераз, что приводит к временной, зависящей от условий жизни организма инактивации метилированного гена. Однако поскольку первичная структура молекулы при этом ДНК не изменяется, это исключение нельзя считать истинным примером передачи информации от белка к ДНК.
Прионы
Прионы — белки, которые существуют в двух формах. Одна из форм (конформаций) белка является функциональной, обычно растворимой в воде. Вторая форма образует нерастворимые в воде агрегаты, часто в виде молекулярных трубочек-полимеров. Мономер — молекула белка — в этой конформации способен просоединяться к другим сходным молекулам белка, переводя их во вторую, прионоподобную, конформацию. У грибов такие молекулы могут передаваться по наследству. Но, как и в случае метилирования ДНК, первичная структура белка в данном случае остаётся прежней, и переноса же информации на нуклеиновые кислоты не происходит.
My mind was, that a dogma was an idea for which there was no reasonable evidence. You see?!" And Crick gave a roar of delight. "I just didn't know what dogma meant. And I could just as well have called it the 'Central Hypothesis,' or — you know. Which is what I meant to say. Dogma was just a catch phrase
I called this idea the central dogma, for two reasons, I suspect. I had already used the obvious word hypothesis in the sequence hypothesis, and in addition I wanted to suggest that this new assumption was more central and more powerful. . As it turned out, the use of the word dogma caused almost more trouble than it was worth. Many years later Jacques Monod pointed out to me that I did not appear to understand the correct use of the word dogma, which is a belief that cannot be doubted. I did apprehend this in a vague sort of way but since I thought that all religious beliefs were without foundation, I used the word the way I myself thought about it, not as most of the world does, and simply applied it to a grand hypothesis that, however plausible, had little direct experimental support.
Живые организмы обладают уникальной способностью передавать генетическую информацию от поколения к поколению, сохраняя свои наследственные свойства. Материальным носителем воспроизводства наследственной информации является нуклеиновая кислота, имеющая для этого определенное химическое строение и биологические свойства. В большинстве организмов эту функцию выполняет ДНК. Исключение составляют отдельные вирусы, у которых носителем информации является РНК. С участием нуклеиновых кислот происходит образование всех белков, являющихся материальной основой всех жизненных процессов. Каждый живой организм содержит свои специфические белки, которыми он отличается от других организмов. Информация, определяющая особенности структуры белков, закодирована в ДНК и передается в ряду поколений молекулами ДНК.
Процесс переноса генетической информации (одной из форм биологической памяти) является определяющим и очень важным для развития и нормальной жизнедеятельности клеток организма. Его можно упрощенно представить схемой:

Виды переноса генетической информации
Прежде чем раскрыть виды переноса генетической информации следует подчеркнуть, что они основаны на матричном механизме синтеза (воспроизводства) новых молекул. Это означает, что для синтеза новой ДНК или РНК необходимы соответствующие матрицы. Точность копирования обеспечивается правилом комплементарности азотистых оснований, согласно которому происходит спаривание А с Т в ДНК (или с У в РНК) и Г с Ц. Благодаря этому порядок чередования нуклеотидов в каждой новой полинуклеотидной цепи комплементарен матрице. Матричный синтез позволяет очень быстро, экономно и с большой точностью (а это очень важно, поскольку речь идет о наследственных свойствах) воспроизводить имеющуюся в клетке генетическую информацию.
Следует отметить три вида переноса генетической информации, имеющих место на различных уровнях организации живой материи:
1. Репликация (самоудвоение, копирование). Это перенос генетической информации в пределах одного класса нуклеиновых кислот: в основном от ДНК к ДНК или у некоторых вирусов от РНК к РНК. Этот процесс происходит только во время деления клетки (на стадии S-фазы митотического цикла) и размножения вирусов и сопровождается репликацией всей молекулы ДНК или РНК. Молекула ДНК расплетается и на ее одиночных цепях в результате репликации образуются точные копии исходной ДНК, т.е. полученные ДНК похожи друг на друга и на исходную материнскую; следовательно наследственная информация сохраняется.
Таким образом, в результате репликации из одной молекулы образуются две новые абсолютно одинаковые молекулы ДНК: одна из них остается в материнской клетке, а другая переходит в дочернюю.
Возможна также репликация отдельных фрагментов ДНК, которая называется амплификацией.
2. Транскрипция , или переписывание. Это перенос генетической информации между разными классами нуклеиновых кислот: ДНК РНК. В отличие от репликации происходит копирование не всей молекулы ДНК, а только ее отдельных участков (цистронов). В ходе транскрипции образуются разные виды РНК (мРНК, тРНК, рРНК), участвующие в биосинтезе белка. Цистроны ДНК содержат информацию о структуре всех типов РНК и о структуре всех белков данного вида организма.

Различают транскрипцию прямую (от ДНК к РНК) и обратную (от РНК к ДНК). Подробно прямая транскрипция будет рассмотрена ниже. Обратная транскрипция впервые была выявлена у РНК-содержащих онкогенных вирусов, и осуществляется она специальным ферментом – обратной транскриптазой, или ревертазой. Сначала к матрице РНК вируса с помощью этого фермента комплементарно присоединяются дезоксирибонуклеозидтрифосфаты и синтезируется одна цепь ДНК. При этом образуется гибридная объединенная молекула РНК–ДНК. Затем фермент РНКаза Н удаляет рибонуклеотидную цепь из гибридной молекулы, а на одиночной цепи ДНК комплементарно в присутствии фермента ДНК-полимеразы происходит синтез второй цепи ДНК:
Образовавшаяся ДНК (копия вирусной РНК) встраивается в ДНК клетки-хозяина и вызывает опухолевую трансформацию клетки.
Трансляция может быть только прямой. Перенос генетической информации можно представить в виде схемы:

Направление переноса генетической информации от ДНК через РНК к белку называется центральным постулатом молекулярной генетики, согласно которому не может быть переноса информации от белка к РНК, но допускается от РНК к ДНК.
БИОСИНТЕЗ БЕЛКА
Биосинтез белка является центральным вопросом биохимии. Раскрытие его имеет важное теоретическое и практическое значение. Учение о биосинтезе белка тесно связано с такими важнейшими проблемами, как наследственность, изменчивость, приспособляемость, естественный отбор, выведение новых форм растительных и животных организмов, с разработкой методов управления процессами жизнедеятельности организма.
Развитие генной инженерии поднимает науку на качественно новый уровень.
Роль нуклеиновых кислот
В живых организмах синтезируются тысячи различных специфических белков. Они отличаются друг от друга в первую очередь первичной структурой, информация о которой заложена в ДНК. Однако сама ДНК не используется в качестве непосредственной матрицы для синтеза белка.
Информация о структуре, записанная в геноме ДНК, передается к рибосомам с помощью информационной РНК (иРНК), которая служит связующим звеном между генами и системой белкового синтеза. Этот процесс называется транскрипциейили переписыванием.
Таким образом, иРНК является точной копией генетической информации, закодированной в определенном участке ДНК, а именно информации о последовательности аминокислот в белках. У прокариот иРНК образуется сразу же в процессе транскрипции на ДНК, и по мере ее постепенного отделения от матрицы ДНК к ней присоединяются рибосомы и начинается синтез белка. В эукариотических клетках в процессе транскрипции вначале синтезируется предшественник – пре-иРНК, которая затем уже превращается в иРНК. Затем иРНК поступает в цитоплазму к рибосомам и выполняет роль матрицы, поэтому ее называют еще матричной РНК (мРНК).
Другие разновидности РНК (рРНК, тРНК) также синтезируются на молекуле ДНК и являются частью аппарата белкового синтеза.
В норме поток генетической информации в клетке идет в следующем направлении:


Рис.1 Общая принципиальная схема биосинтеза белка.
Генетический код
Генетический код связывает последовательность нуклеотидов в ДНК и последовательность аминокислот в белках. Следовательно, для каждой аминокислоты существует свой кодон (кодовое слово) для пе ревода последовательности нуклеотидов в соответствующую последовательность аминокислот.
Используя 4-буквенный алфавит нуклеотидных оснований ДНК (А, Г, Т, Ц), можно теоретически составить такие варианты:
1. Если аминокислота кодируется одним основанием, то можно получить полипептидную цепь только из 4 разновидностей аминокислот (4 1 = 4).
2. Если предположить, что кодон для каждой аминокислоты содержит два подряд расположенных нуклеотида (дуплет), то возможно 4 2 = 16 сочетаний (такого числа кодонов тоже недостаточно для кодирования 20 аминокислот). В этом случае полипептидная цепь состояла бы только из 16 разновидностей аминокислот.
3. Если взять комбинации по три нуклеотида (триплет), то получается 4 3 = 64 кодона, т.е. с избытком. Триплетная природа генетического кода была подтверждена многочисленными экспериментами. Генетическая информация (порядок расположения нуклеотидов в гене ДНК) передается в процессе биосинтеза белков на мРНК.
Код или кодон – это триплет из трех подряд расположенных пуриновых или пиримидиновых оснований на мРНК, ответственных за присоединение определенной аминокислоты в полипептидной цепи.
Таблица 1
Как следует из таблицы, генетический код оказался вырожденным (избыточным), так как каждой аминокислоте (кроме метионина и триптофана) соответствует больше одного кодона.
Важным свойством генетического кода является его неперекрываемость – каждый из триплетов независим друг от друга.
Код является специфичным – каждой аминокислоте соответствуют только определенные кодоны, которые не могут кодировать другие аминокислоты.
Код – колинеарен, т.е соблюдается соответствие линейной последовательности триплетов мРНК и аминокислот в полипептиде.
Одно из существенных свойств кода – его универсальность . Тринуклеотиды, кодирующие одну и ту же аминокислоту, имеют одинаковый состав и последовательность для всех организмов (бактерий, растений, животных и человека).
Все перечисленные выше свойства генетического кода характерны для всех живых организмов. Благодаря универсальности кода возможна генетическая инженерия. Таким образом, путь информации от ДНК к белку представляется следующим образом:

Рибосомы, их структура и химический состав
В 50-е годы нашего столетия в исследованиях П.Замечника и других авторов было показано, что биосинтез белков происходит в небольших субклеточных образованиях, которые получили назва ние рибосом. Последние были найдены во всех клетках прокариот и эукариот. Если выделить рибосомы путем дифференциального центрифугирования и посмотреть под электронным микроскопом, то они представляются как плотные округлые гранулы сферической формы, состоящие из двух субъединиц: малой и большой. Рибосомы характеризуются коэффициентом или константой седиментации, которая определяется ультрацентрифугированием и обозначается буквой S (единица Сведберга, 1 10 –13 с). По размерам и молекулярной массе все рибосомы делят на три группы. Первую группу образуют относительно мелкие бактериальные рибосомы. Рибосомы прокариот имеют константу седиментации 70 ед. Сведберга и обозначаются 70S; они диссоциируют на две субъединицы с молекулярной массой (М.м.):

Вторую группу образуют крупные рибосомы эукариотических клеток. Они имеют константу седиментации 80S и состоят из двух субъединиц:

Третью группу составляют рибосомы митохондрий и хлоропластов эукариотических клеток. Рибосомы митохондрий относятся к классу 70S, однако они различаются по коэффициенту седиментации у разных групп эукариот. Так, у грибов он составляет 70–74S, у высших животных – 55–60S, у высших растений – около 80S. Рибосомы хлоропластов более однородны по этому признаку, коэффициент их седиментации равен 67–70S.
Обычно рибосомы изображают в виде симметричной фигуры, в которой 30S субчастица лежит на 50S-субъeдинице, имеющей форму, близкую к сфере. На основе рентгеноструктурного анализа и электронно-микроскопических методов было показано, что трехмерная структура частиц рибосом весьма сложна. Малая субчастица изогнута в виде телефонной трубки, а большая напоминает ковш(рис.2) . Две субчастицы соответствуют друг другу, хотя между ними остается щель. Через щель проходит молекула мРНК, вдоль которой в процессе биосинтеза белка перемещается рибосома. Из этой щели появляется и новосинтезированная полипептидная цепь.

Рис.2 Большая (а) и малая (б) субчастицы рибосом (в)
В состав рибосомы входят рРНК, белки, низкомолекулярные соединения: ди- и полиамины, различные соли, ионы двухвалентных металлов Мg 2+ , Са 2+ , Мn 2+ и др.
Рибосомы 80S построены более сложно, они содержат больше белков, и это имеет определенное значение в их функции.
Этапы биосинтеза белка
1. Активация аминокислот, соединение их с тРНК и перенос к рибосомам .
Этот процесс идет в одну стадию, но для удобства и лучшего понимания его разбивают на два этапа.
а) Активирование аминокислот – образование аминоациладенилатов.
Аминокислоты в цитоплазме находятся в неактивном состоянии. Они активируются по карбоксильной группе за счет энергии АТФ, в присутствии солей Mg 2+ с помощью специальных ферментов аминоацил-тРНК-синтетаз, обозначаемых сокращенно АРСазы. Эти ферменты обеспечивают оба этапа процесса – активирование аминокислот и соединение их с тРНК. Каждый фермент обладает двойной специфичностью: к определенной аминокислоте и к соответствующей ей тРНК.
б) Перенос аминоациладенилатов к месту синтеза белка, к рибосомам.
Активированные аминокислоты должны быть доставлены к рибосомам и этот перенос осуществляется тРНК. тРНК связывает в единое целое мРНК, рибосому, специфическую аминокислоту. Присоединение активированной аминокислоты к специфической тРНК происходит путём образования сложноэфирной связи между СООН-группой соответствующей аминокислоты и 3-ОН группой концевого остатка адениловой кислоты тРНК.
Реакцию катализирует тот же фермент, что и реакцию активирования аминокислот – аминоацил- т РНК-синтетаза .

2. Процесс трансляции на рибосомах .
Процесс перевода нуклеотидной последовательности мРНК в аминокислотную получил название трансляции. Трансляция включает три этапа: инициацию (начало синтеза полипептидной цепи), элонгацию (ее рост) и терминацию (окончание синтеза).
. Каждый из белков закодирован в отдельном участке мРНК – цистроне, имеющем свои инициирующие и терминирующие триплеты.

Рис3 . Схема трансляции у прокариот
Образование пептидной связи (рис.3) осуществляется в такой последовательности:
1. В присутствии фермента разрывается макроэргическая связь на N-формилметионил-тРНК фмет . 2. К освободившейся связи при соединяется Н от -NH2- группы фенилаланил-тРНК фен . 3. Карбонильный остаток N-формилметионина переходит к -NH2-фенилаланил-тРНК фен – возникает первая пептидная связь с образованием в А-участке дипептида: N-формилметионил- фенилаланил-тРНК фен . 4. В процессе транслокации мРНК продвигается на один кодон. 5. тРНК фмет без инициирующей аминокислоты уходит с рибосомы. В освободившийся А-участок входит тРНК ала , возникает вторая пептидная связь и образуется трипептид состава N-формилметионил-фенилаланил-аланил-тРНК ала , который перемещается затем в П-участок; А-участок освобождается и процесс повторяется. 6. Рибосома достигает терминирующего триплета УГА и диссоциирует на 30S- и 50S-субчастицы, при этом отделяется полипептидная цепь с заданным расположением остатков аминокислот.
Вторичная и третичная структуры белков формируются в процессе трансляции по мере удлинения полипептидной цепи. Трехмерную конформацию белок окончательно принимает уже после своего отделения.
Ингибиторы биосинтеза белков.
Механизм действия антибиотиков
Прекращение матричного биосинтеза белков приводит к гибели клетки.
Ингибиторами биосинтеза белков могут быть различные вещества, в том числе антибиотики, токсины, алкалоиды, антиметаболиты (аналоги) структурных единиц нуклеиновых кислот и др. Они широко используются в биохимических исследованиях как инструменты для раскрытия механизма отдельных этапов процесса биосинтеза белков, так как оказалось, что среди них можно подобрать такие, которые избирательно ингибируют специфические фазы белкового синтеза.
Антибиотики – это вещества, продуцируемые микроорганизмами, плесенью, грибами, высшими растениями, животными тканями в процессе их жизнедеятельности, а также полученные синтетическим путем. Они обладают способностью оказывать бактериостатическое или бактерицидное действие. Антибиотики, взаимодействующие с ДНК, нарушают ее матричные функции и подавляют репликацию или транскрипцию, или оба эти процесса. Противоопухолевые антибиотики практически одинаково взаимодействуют с ДНК как опухолевых, так и нормальных клеток, т.е. они не отличаются избирательностью действия.
Антибиотики, ингибирующие процесс трансляции, взаимодействуя с белковыми факторами и рибосомами, применяются главным образом как противобактериальные средства. Они отличаются достаточно высокой избирательностью действия и часто сравнительно малотоксичны для человеческого организма. Это объясняется тем, что у микроорганизмов рибосомы (70S), отдельные ферменты и белковые факторы, входящие в рибосому, несколько отличаются от рибосом (80S) и соответствующих белков эукариот. Однако некоторые из них могут действовать на рибосомы митохондрий человека, так как рибосомы в митохондриях имеют меньший размер, чем 80S рибосомы в цитоплазме, поэтому митохондриальный биосинтез белка по своему механизму близок к синтезу белка у прокариот. Отмечается также в некоторой степени действие отдельных антибиотиков и на 80S рибосомы. Именно поэтому из описанных многих сотен антибиотиков лишь несколько десятков нашли применение в медицинской практике. Следовательно, успех антибиотикотерапии зависит от умелого, рационального их применения. Необходимо учитывать возможность осложнений и аллергических реакций. Ввиду этого совершенно недопустимо самолечение антибиотиками без назначения врача и отпуск их без рецепта.
Все ингибиторы матричного биосинтеза белка разделяют по механизму действия на: ингибиторы репликации, транскрипции, созревания (процессинга) и транспорта РНК, трансляции и др.
Препараты, усиливающие биосинтез белка
Читайте также: