
Процесс созревания ирнк кратко
Обновлено: 05.07.2024
Процессинг — это этап формирования функционально активных молекул РНК из первоначальных транскриптов. Процессинг рассматривают как посттранскрипционные модификации РНК, характерные для эукариот. (У прокариот процессы транскрипции и трансляции иРНК идут почти одновременно. Этот тип РНК у них процессинга не претерпевает.)
В результате процессинга первичные транскрипты РНК превращаются в зрелые РНК. Поскольку существует несколько различных типов РНК, то для каждого из них характерны свои модификации.
Процессинг информационной (матричной) РНК
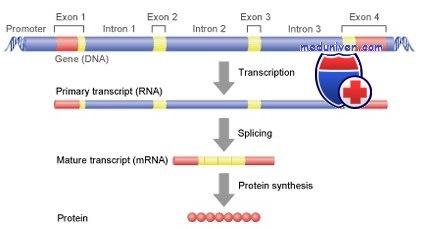
На участках ДНК, кодирующих структуру белка, образуется предшественник информационной (матричной) РНК (пре-иРНК). Пре-иРНК копирует всю нуклеотидную последовательность ДНК от промотора до терминатора транскриптона. То есть она включает концевые нетранслируемые области (5' и 3'), интроны и экзоны.
Процессинг пре-иРНК включает в себя кэпирование, полиаденилирование, сплайсинг, а также некоторые другие процессы (метилирование, редактирование).
Кэпирование — это присоединение 7-метил-ГТФ (7-метилгуанозинтрифосфат) к 5'-концу РНК, а также метилирование рибозы двух первых нуклеотидов.
В результате полиаденилирования к 3'-концу РНК присоединяется полиадениловый участок (поли-А) длинной примерно 100-200 нуклеотидов (содержащих аденин). Данные реакции обеспечивает фермент поли-А-полимераза. Сигналом к полиаденилированию служит последовательность AAUAAACA на 3'-конце. В месте -CA происходит разрезание молекулы иРНК.
Поли-А защищает молекулу РНК от ферментативного распада.
Кэпирование и полиаденилирование происходят еще на этапе транскрипции. Кэп образуется сразу после высвобождения из РНК-полимеразы 5'-конца синтезируемой РНК, а поли-А образуется сразу после терминации транскрипции.
Сплайсинг представляет собой вырезание интронов и соединение экзонов. Экзоны могут соединяться по-разному. Таким образом из одного транскрипта могут образовываться разные иРНК. В сплайсинге информационной РНК участвуют малые ядерные РНК, которые имеют участки, комплементарные концам интронов и связываются с ними. Кроме мяРНК в сплайсинге участвуют различные белки. Все вместе (белки и мяРНК) формируют нуклеопротеидный комплекс — сплайсосому.
После процессинга иРНК становится короче своего предшественника иногда в десятки раз.
Процессинг других видов РНК
При процессинге молекул рибосомальных и транспортных РНК не происходит кэпирования и полиаденилирования. Модификации данных видов РНК происходят не только у эукариот, но и у прокариот.
Три вида рибосомальной РНК эукариот образуются в результате расщепления одного транскрипта (45S-РНК).
Процессинг ряда транспортных РНК может также включать расщепление одного транскрипта, другие тРНК получаются без расщепления. Особенностью процессинга тРНК является то, что молекула РНК проходит длинную цепь модификаций нуклеотидов: метилирование, дезаминирование и др.
Процесс формирования зрелых (то есть функционально активных) молекул РНК из предшественников, называемый процессингом – важнейший этап образования многих видов и-РНК. Все стабильные РНК бактерий также синтезируются в виде первичных транскриптов, из которых затем образуются зрелые р-РНК или т-РНК. Аналогичным путем происходит образование и эукариотических р-РНК и т-РНК.
В отличие от эукариотических, большинство бактериальных и-РНК представляют собой первичные транскрипты, которые в состоянии непосредственно взаимодействовать с рибосомами, так что только в очень редких случаях их трансляции должен предшествовать процессинг.
У эукариот, напротив, образование зрелой и-РНК, кодируемой прерывистым геном – наиболее сложный процесс из всех видов процессинга РНК, включающий большое число реакций сплайсинга(вырезания), обеспечивающих соединение экзоновпутем удаления интронов.
Все реакции процессинга являются высокоспецифичными, что приводит к образованию зрелых молекул РНК с уникальными 5'- и 3'-концами. Реакции процессинга осуществляются специфическими ферментами (рибонуклеазами), способными расщеплять фосфодиэфирные в молекулах РНК.

Рис. Общая схема процессинга рибосомных и транспортных РНК прокариот.
Синтез информационной РНК (иРНК): строение генов, транскрипция
Большинство процессов метаболизма в организме катализируются белковыми ферментами. Кроме того, белки — основные структурные компоненты тела человека. Аминокислотные последовательности всех белков зашифрованы в ДНК, а процесс превращения закодированной информации в сам белок включает её транскрипцию на гяРНК, процессинг на иРНК, трансляцию на полипептид и окончательную сборку белка.
Строение гена
В отличие от прокариот у эукариот большинство генов имеют участок ДНК, который прерывает кодирующую последовательность. Данные некодирующие фрагменты называют нитронами, в то время как другие, кодирующие участки— экзонами. У обеих групп после кодирующего участка присутствуют лидерная и трейлерная последовательности, а также ряд последовательностей, контролирующих процесс транскрипции.
Фрагмент, расположенный ниже участка начала транскрипции (5'—3'), называют лидерной последовательностью, он не транслируется. Затем следует кодирующий участок, обычно прерываемый одним или несколькими интронами, а после — некодирующий трейлерный (концевой) участок, на конце которого участок полиаденилирования (поли-А-сайт), имеющий вариабельную последовательность наподобие 5'-ААТАА-3' (5'-ААУАА-3' на РНК-транскрипте) длиной 10-30 пар нуклеотидов в направлении 3'—5'.
Интроны начинаются последовательностью ГТА(/Г)ГАГТ и заканчиваются серией из Ц- или Т-оснований, предшествующих АГ. Для удаления интрона значение имеют первые основания Г и Т (Г и У в гяРНК) и последние АГ, а также остаток аденина в составе последовательности ближе к 5'-концу. Участок, находящийся ближе к 5'-концу, известен под названием донор, ближе к З'-концу — акцептор, а остаток аденина называют участком ветви.
Транскрипция при синтезе иРНК
Таким образом формируется транскрипционное вздутие. Как только он достигает участка начала транскрипции, происходит отщепление одного из факторов транскрипции и присоединение другого, после чего начинается процесс синтеза РНК.
Используя в качестве матрицы цепь в направлении 3'—5' (слева направо), РНКаза II поочерёдно захватывает рибонуклеотиды и соединяет их друг с другом, образуя комплементарную последовательность РНК, ориентированную в обратном направлении (то есть от 5' к 3').
Другими словами, используя правила комплементарного спаривания оснований при взаимодействии с матричной цепью, РНКаза создаёт точную РНК-копию кодирующей цепи. Фермент транскрибирует лидерный и трейлерный участки, экзоны, интроны и (по всей видимости, напрасно) продвигается дальше в направлении 5'—3'.
Факторы транскрипции при синтезе иРНК
Факторы транскрипции — белки, прикрепляющиеся к промоторной последовательности и запускающие процесс транскрипции. В их состав обычно входят активационный домен и ДНК-связывающий домен. Активационные домены богаты глутаматом, а также аспартатом или пролином, которые облегчают формирование транскрипционного комплекса. Кроме того, различают четыре типа ДНК-связывающих доменов.
• Спираль-петля-спираль состоит из двух белковых а-спиралей, которые соединены длинной, гибкой петлёй, позволяющей параллельно упаковывать их близко друг к другу. Считают, что данная структура осуществляет контроль процесса транскрипции путём блокирования других регуляторных белков гена.
• Спираль-поворот-спираль состоит из двух коротких а-спиралей, разделённых аминокислотной последовательностью, слишком короткой, чтобы позволить им лежать в одной плоскости. Этот фрагмент — характерный признак гомеобокса (см. главу 12).
Процессинг РНК
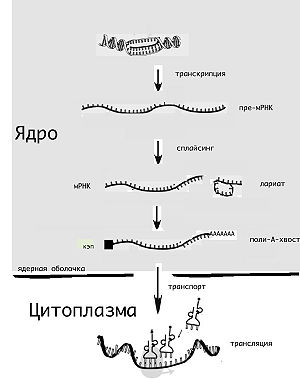
Для того чтобы только что синтезированные гяРНК стали кодирующими матрицами для последующей трансляции и образования полипептидов, они претерпевают ковалентное видоизменение. При этом вначале к 5'-концу в обратном направлении прикрепляется 7-метил-ГТФ (кэп). Как только на цепи гяРНК возникает участок полиаденилирования, она в этом месте расщепляется, а затем при помощи полиА-полимеразы происходит присоединение 100—200 остатков адениловой кислоты и таким образом формируется поли-А-хвост (полнаденильный хвост).
Наряду с кэпом поли-А-хвост предположительно защищает молекулу от разрушения экзонуклеазами, служит так называемым паспортом, необходимым для её попадания в цитоплазму, а позже становится сигнальным участком для рибосомы, указывающим на возможность начала трансляции.
Молекула гяРНК в среднем содержит около 7000 нуклеотидов, количество которых в иРНК сокращается до 1200 путём удаления примерно 50 интронов. Характерная особенность гистонных генов — отсутствие интронов.
Рибонуклеиновые комплексы, которые удаляют нитроны, называют сплайсомами. Они имеют в своём составе несколько малых ядерных РНК (U1—U6), каждая из которых соединена со специфическим белком. Рибонуклеопротеин, содержащий малую ядерную РНК U1 (Ul-малая ядерная РНК), благодаря наличию комплементарной последовательности, присоединяется к участку начала сплайсинга в направлении 3'—5'.
К участку ветви прикрепляется малая ядерная РНК U2, которая затем связывается с U1, в результате чего возникает петля гяРНК. После этого U2 отсекает гяРНК в направлении 3'—5' сразу после последовательности Г—У (см. выше) и соединяет ближний к 5'-конец интрона с участком соединения, образуя так называемое лассо. Конец интрона, находящийся ближе к 3', отсекается сразу после последовательности А—Г, распуская лассо РНК. При этом сплайсома соединяет между собой экзоны.
Иногда в некоторых транскриптах (особенно при производстве антител) обнаруживают альтернативные механизмы сплайсинга, однако ошибки в данном процессе играют важную роль в развитии многих генетических заболеваний. Так, церебральный паралич и задержка умственного развития при синдроме Жильбера обусловлены внедрением Т—А в нормальную последовательность ТАТАА промотора гена УДФ-гликозилтрансферазы. А-аманитин, содержащийся в бледной поганке (Amanita phalloides), блокирует действие РНКазы II.
Антибиотик рифампицин блокирует транскрипцию у бактерий путём связывания с b-субъединицей бактериальной РНК-полимеразы, в то время как актиномицин внедряется между парами оснований Г—С .
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Длина типичной зрелой мРНК составляет от нескольких сотен до нескольких тысяч нуклеотидов. Самые длинные мРНК отмечены у (+)оц РНК-содержащих вирусов, например пикорнавирусов, однако следует помнить, что у этих вирусов мРНК образует весь их геном.
ДНК нередко сравнивают с чертежами для изготовления белков. Развивая эту инженерно-производственную аналогию, можно сказать, что, если ДНК — это полный набор чертежей для изготовления белков, находящийся на хранении в сейфе директора завода, то мРНК — временная рабочая копия чертежа, выдаваемая в сборочный цех.
Гипотеза о значении РНК в синтезе белков была высказана Торбьёрном Касперссоном (Torbjörn Caspersson) на основе исследований 1937—1939 гг., в результате которых было показано, что клетки, активно синтезирующие белок, содержат большое количество РНК. Подтверждение гипотезы было получено Юбером Шантренне (Hubert Chantrenne).
Содержание
Транскрипция
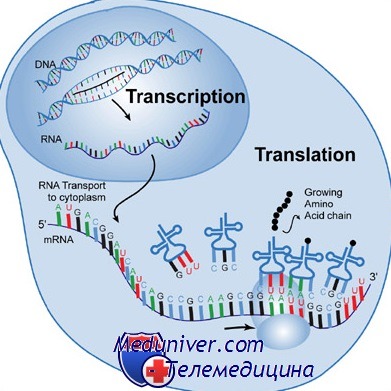
Транскрипцией называют процесс копирования генетической информации с ДНК на РНК, в частности на мРНК. Транскрипция осуществляется ферментом РНК-полимеразой, строящей, согласно принципу комплементарности, копию участка ДНК на основании одной из цепей двойной спирали. Этот процесс как у эукариот, так и у прокариот организован одинаково. Основное различие между про- и эукариотами состоит в том, что у эукариот РНК-полимераза во время транскрипции ассоциируется с мРНК-обрабатывающими ферментами, поэтому у них обработка мРНК и транскрипция могут проходить одновременно. Короткоживущие необработанные или частично обработанные продукты транскрипции называются пред-мРНК; после полной обработки — зрелая мРНК.
Обработка эукариотической пред-мРНК
Сплайсинг

Схема сплайсинга, в процессе которого пре-мРНК созревает в зрелую РНК. Зелёный — нетранслируемые участки (UnTranslated Regions, UTR), синий — интроны, красный — транслируемые (кодирующие белок) участки.
Сплайсинг — это процесс, в котором из пре-мРНК удаляются участки некодирующей белок последовательности, называемые интронами; последовательности, которые остаются, включают в себя кодирующие белки нуклеотиды, и называются экзонами. Иногда продукты сплайсинга пре-мРНК могут быть соединены разными способами, позволяя одному гену кодировать несколько белков. Этот процесс называется альтернативным сплайсингом. Сплайсинг обычно производится РНК-белковым комплексом, который называется сплайсосома, но некоторые молекулы мРНК также могут катализировать сплайсинг без участия белков(см. рибозимы) [2] .
Транспорт
Другое различие между эукариотами и прокариотами — транспорт мРНК. Из-за того, что эукариотические транскрипция и трансляция пространственно разделены, эукариотические мРНК должны быть выведены из ядра в цитоплазму [3] . Зрелые мРНК распознаются по наличию модификаций и покидают ядро через ядерные поры, в цитоплазме мРНК образует нуклеопротеидные комплексы — информосомы, в составе которых транспортируется к рибосомам.
Трансляция
Поскольку прокариотическая мРНК не нуждается в обработке и транспортировке, трансляция рибосомой может начаться немедленно после транскрипции. Следовательно, можно сказать, что трансляция у прокариот совмещена с транскрипцией и происходит ко-транскрипционнно.
Эукариотическая мРНК должна быть обработана и доставлена из ядра в цитоплазму, и только тогда может быть транслирована рибосомой. Трансляция может происходить как на рибосомах, находящихся в цитоплазме в свободном виде, так и на рибосомах, ассоциированных со стенками эндоплазматического ретикулума. Таким образом, у эукариот трансляция не совмещена напрямую с транскрипцией.
Регуляция трансляции
Так как у прокариот транскрипция совмещена с трансляцией, прокариотическая клетка может быстро реагировать на изменения в окружающей среде путём синтеза новых белков, то есть регуляция происходит, в основном, на уровне транскрипции. У эукариот из-за необходимости в редактировании и транспорте мРНК ответ на внешние стимулы занимает больше времени. Поэтому их синтез белка интенсивно регулируется на посттранскрипционном уровне. Не всякая зрелая мРНК транслируется, поскольку в клетке существуют механизмы регуляции экспрессии белков на пост-транскрипционном уровне, например, РНК-интерференция.
Разрушение
По прошествии некоторого времени, определяемого её нуклеотидной последовательностью, в частности, длиной полиаденинового участка на 3' конце молекулы, мРНК разрушается на составляющие её нуклеотиды с участием РНКаз. Как правило, разрушение начинается с удаления кэпа на 5' конце, полиаденинового хвоста на 3' конце и затем нуклеазы одновременно разрушают мРНК в 5' ->3' и 3' ->5' направлении. мРНК, в которой сигнал завершения синтеза белка, стоп-кодон, в результате ошибки транскрипции находится в середине кодирующей последовательности, подвержена особой быстрой форме деградации, НМД, (nonsense-mediated decay).
Строение зрелой мРНК
5' Кэп
5' кэп (или кап) (от англ. cap — шапочка) — это модифицированный гуанидиновый нуклеотид, который добавляется на 5' (передний) конец незрелой мРНК. Эта модификация очень важна для узнавания мРНК при инициации трансляции, а также для защиты от 5’нуклеаз — ферментов, разрушающих цепи нуклеиновых кислот с незащищённым 5'-концом.
Кодирующие области
Кодирующие области состоят из кодонов — следующих непосредственно друг за другом последовательностей из трёх нуклеотидов, каждая из которых соответствует в генетическом коде определённой аминокислоте или началу и концу синтеза белка. Кодирующие области начинаются со старт-кодона и заканчиваются одним из трёх стоп-кодонов. Считывание последовательности кодонов и сборка на её основе последовательности аминокислот синтезируемой молекулы белка осуществляется рибосомами при участии транспортных РНК в процессе трансляции. В дополнение к кодированию белков, части кодирующих областей могут служить управляющими последовательностями. Например, вторичная структура РНК в некоторых случаях определяет результат трансляции.
Моноцистронная и полицистронная мРНК
мРНК называют моноцистронной, если она содержит информацию, необходимую для трансляции только одного белка (один цистрон). Полицистронная мРНК кодирует несколько белков. Гены (цистроны) в такой мРНК разделены интергенными, некодирующими последовательностями. Полицистронные мРНК характерны для прокариот и вирусов, у эукариот большая часть мРНК является моноцистронной [4] . Полицистронные мРНК встречаются у эукариот в митохондриях.
Нетранслируемые области
Нетранслируемые области — участки РНК, расположенные до старт-кодона и после стоп-кодона, которые не кодируют белок. Они называются 5'-нетранслируемая область и 3'-нетранслируемая область, соответственно. Эти области транскрибируются в составе того же самого транскрипта, что и кодирующий участок. Нетранслируемые области имеют несколько функций в жизненном цикле мРНК, включая регуляцию стабильности мРНК, локализации мРНК и эффективности трансляции. Стабильность мРНК может контролироваться 5'- и/или 3'-областью из-за различной чувствительности к ферментам, которые отвечают за деградацию РНК — РНКазам и регуляторным белкам, которые убыстряют или замедляют деградацию [5] .
3' полиадениновый хвост
Вторичная структура
Примерами вторичной структуры могут служить стебель-петля и псевдоузел [7]
Вторичные структуры в мРНК служат для регуляции трансляции. Например, вставка в белки необычных аминокислот, селенометионина и пирролизина, зависит от стебля-петли, расположенной в 3' нетранслируемой области. Псевдоузлы служат для программированного изменения рамки считывания генов.
Сразу после синтеза первичные транскрипты РНК по разным причинам еще не имеют активности, являются "незрелыми" и в дальнейшем претерпевают ряд изменений, которые называются процессинг . У эукариот процессингу подвергаются все виды пре-РНК, у прокариот – только предшественники рРНК и тРНК.
Процессинг предшественника матричной РНК
При транскрипции участков ДНК, несущих информацию о белках, образуются гетерогенные ядерные РНК, по размеру намного превосходящие мРНК. Дело в том, что из-за мозаичной структуры генов эти гетерогенные РНК включают в себя информативные ( экзоны ) и неинформативные ( интроны ) участки.
1. Сплайсинг (англ. splice – склеивать встык) – особый процесс, в котором при участии малых ядерных РНК происходит удаление интронов и сохранение экзонов.
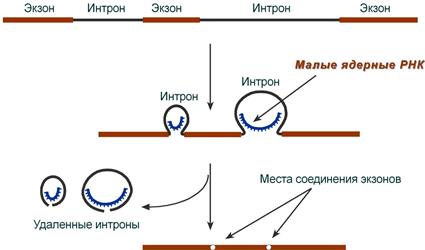
Последовательность событий сплайсинга
2. Кэпирование (англ. cap – шапка) – происходит еще во время транскрипции. Процесс состоит в присоединении к 5'-трифосфату концевого нуклеотида пре-мРНК 5'-углерода N 7 -метил-гуанозина.
"Кэп" необходим для защиты молекулы РНК от экзонуклеаз, работающих с 5'-конца, а также для связывания мРНК с рибосомой и для начала трансляции.
3. Полиаденилирование – при помощи полиаденилат-полимеразы с использованием молекул АТФ происходит присоединение к 3'-концу РНК от 100 до 200 адениловых нуклеотидов, формирующих полиадениловый фрагмент – поли(А)-хвост. Поли(А)-хвост необходим для защиты молекулы РНК от экзонуклеаз, работающих с 3'-конца.
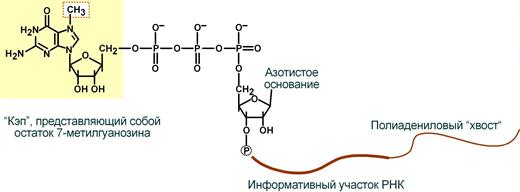
Схематичное представление матричной РНК после процессинга
Процессинг предшественника рибосомальной РНК
Предшественники рРНК являются более крупными молекулами по сравнению со зрелыми рРНК. Их созревание сводится к разрезанию прерибосомной РНК на более мелкие формы, которые уже непосредственно участвуют в формировании рибосомы. У эукариот существуют четыре типа рРНК – 5S-, 5,8S-, 18S- и 28S-рРНК . При этом 5S-рРНК синтезируется отдельно, а большая прерибосомная 45S-РНК расщепляется специфичными нуклеазами с образованием 5,8S-рРНК, 18S-рРНК и 28S-рРНК.
У прокариот молекулы рибосомальной РНК совсем иные по своим свойствам (5S-, 16S-, 23S-рРНК), что является основой изобретения и использования ряда антибиотиков в медицине.
Процессинг предшественника транспортной РНК
1. Модификация нуклеотидов в молекуле путем дезаминирования, метилирования, восстановления.
Например, образование псевдоуридина и дигидроуридина.
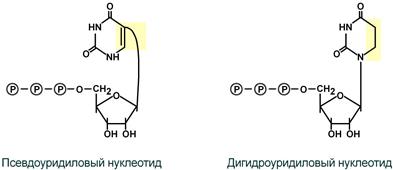
Строение модифицированных уридиловых нуклеотидов
2. Формирование антикодоновой петли происходит путем сплайсинга (вернуться вверх) и удаления интрона в средней части пре-тРНК.
3. Формирование на 3'-конце последовательности ЦЦА. Для этого у одних пре-тРНК с 3'-конца удаляются лишние нуклеотиды до "обнажения" триплета ЦЦА, у других идет присоединение этой последовательности.
Читайте также: