
Опишите хромосомные основы расщепления и независимого перераспределения генов кратко
Обновлено: 02.07.2024
Биологическую роль хромосом сложно не заметить. Правила стабильности числа, парности, индивидуальности, сложное поведение мейоза и митоза укрепили биологов во мнении, что хромосомы значимы и напрямую связаны, в частности, с передачей наследственных признаков.
Если гены, определяющие комбинированные признаки, находятся в разных парах гомологических хромосом, то происходит независимое комбинирование признаков. Поэтому любой организм имеет количество генов, ограниченное числом пар хромосом, которые могут независимо комбинироваться в мейозе.
К примеру, у кукурузы насчитывается больше 500 генов, у мухи дрозофилы — свыше 1000. У человека больше 30 тысяч генов, при том, что хромосом — 10, 4 и 23 пары.
Это говорит о том, что каждая хромосома содержит огромное количество генов.
Находящиеся в одной хромосоме и образующие группу сцепления гены наследуются вместе.
Сцепленное наследование — это общее наследование генов по Томасу Хант Моргану.
Из этого положения следует, что специфика гаплоидного набора хромосом определяет количество групп сцепления.
Сцепление не всегда бывает абсолютным — к этому заключению пришел Томас Хант Морган в своих дальнейших исследованиях. Он проводил эксперименты на плодовых мушках дрозофилах и выяснил, что полное сцепление случается только в 83% случаев: у одной половины потомства были длинные крылья и серое тело (41,5%), у другой — короткие крылья и черное тело (признаки только родительских форм). 17% случаев связаны с перекомбинацией признаков: 8,5% имело короткие крылья и серое тело, а другие 8,5% — длинные крылья и черное тело.
Причина нарушения сцепления генов — в кроссинговере.
Что такое кроссинговер?
Кроссинговер — это перекрест хромосом, который происходит в профазе I мейоза.
Чем больше расстояние между генами в хромосомах, тем выше вероятность перекреста и больше количество гамет, образованных в результате перекомбинации генов. Частота кроссинговера пропорциональна расстоянию, на котором находятся гены друг от друга. Основное биологическое значение кроссинговера — в увеличении комбинативной изменчивости, которая дает материал для естественного отбора.
Если говорить о количестве, то расстояние между генами коррелирует с частотой, с которой появляются кроссинговерные организмы. В примере выше этот показатель равен 17%. Это расстояние описывает силу сцепления и обозначается процентами рекомбинации кроссинговера или морганидами.
Один процент кроссинговера = одна морганида.
У одних генов наблюдается высокий процент сцепления, а у других — почти не выявляется. При этом в варианте сцепленного наследования максимальная величина кроссенговера — не больше 50%. Если этот показатель будет выше, то пары аллелей смогут беспрепятственно комбинироваться — в таком случае кросенговер невозможно будет отличить от независимого наследования.
Процент кроссинговера учитывается при составлении генетических карт хромосом, с нанесенным на них относительным расстоянием между генами.
Биологическое значение кроссинговера сложно переоценить. Благодаря генетической рекомбинации создаются условия для образования новых комбинаций генов и обеспечивается более высокая жизнеспособность организмов в ходе эволюции.
Основные положения хромосомной теории наследования
Впервые наблюдение хромосом в гаметах и зиготах стало возможно в конце 19 века, когда появились усовершенствованные микроскопы и цитологические методы. Важную роль ядра в регулировании развития признаков организма показал в 1902 году ученый Бовери. Еще ранее, в 1882 году, ученый Флемминг описал поведение хромосом во время митоза. А в 1900 году повторно были открыты законы Менделя. Независимо друг от друга, их оценили трое ученых: Фриз, Корренс и Чермак.
Американский цитолог У. Сеттон и немецкий эмбриолог Т. Бовери, взяв за основу описанные выше данные, предположили, что хромосомы — носители наследственной информации. Так была сформулирована хромосомная теория наследственности.
Согласно этой теории, каждая пара генов находится в паре гомологических хромосом, при этом, каждая хромосома является носителем одного гена. Поскольку у каждого организма признаков намного больше, чем хромосом, то в каждой хромосоме должно находиться большое количество генов.
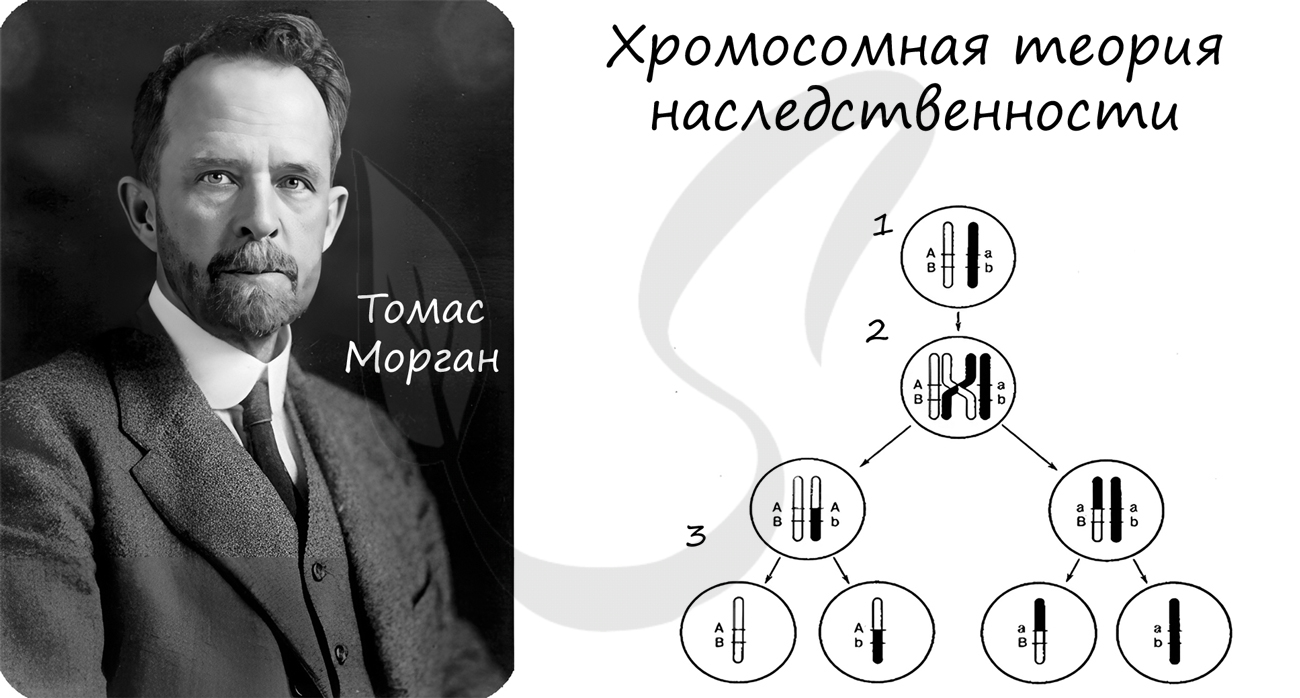
Автором хромосомной теории наследования признан американский генетик Томас Хант Морган, который обнаружил линейный порядок расположения генов в хромосоме.
Описанные им закономерности, которые подтвердились и углубились позже на различных объектах, получили название хромосомной теории наследования.
К концу XIX в. благодаря существенному улучшению оптических характеристик микроскопов и совершенствованию цитологических методов стало возможным наблюдать поведение хромосом в гаметах и зиготах. В 1875 г. Гертвиг обратил внимание на то, что при оплодотворении яиц морского ежа происходит слияние двух ядер — ядра спермия и ядра яйцеклетки. В 1902 г. Бовери продемонстрировал важную роль ядра в регуляции развития признаков организма, а в 1882 г. Флемминг описал поведение хромосом в процессе митоза.
Таблица 24.3. Соответствие событий, происходящих при мейозе и оплодотворении, гипотезе Менделя
Мейоз и оплодотворение
Диплоидные клетки содержат пары гомологичных хромосом
Признаки контролируются парами факторов
Гомологичные хромосомы расходятся во время мейоза
Парные факторы разделяются при образовании гамет
В каждую гамету попадает одна из гомологичных хромосом
Каждая гамета получает один фактор
Только ядро мужской гаметы сливается с ядром яйцеклетки
Факторы передаются из поколения в поколение как дискретные единицы
При оплодотворении пары гомологичных хромосом восстанавливаются; каждая гамета вносит одну из гомологичных хромосом
Каждый организм наследует по одному фактору от каждой из родительских особей
На основе всех изложенных выше данных Сэтгон и Бовери высказали предположение, что хромосомы служат носителями менделевских факторов, и сформулировали теорию, названную хромосомной теорией наследственности. Согласно этой теории, каждая пара факторов локализована в паре гомологичных хромосом, причем каждая хромосома несет по одному из этих факторов. Поскольку число признаков у любого организма во много раз больше числа его хромосом, видимых под микроскопом, каждая хромосома должна содержать множество факторов.
В 1909 г. Иогансен заменил термин фактор, означавший основную единицу наследственности, термином ген. Альтернативные формы гена, определяющие его проявление в фенотипе, назвали аллелями. Аллели — это конкретные формы, которыми может быть представлен ген; они занимают одни и те же места (локусы) в гомологичных хромосомах (рис. 24.5).
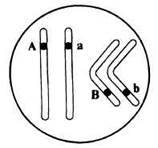
Рис. 24.5. Клетка с двумя парами гомологичных хромосом. Расположение двух разных генных локусов показано черными кружками. В данном случае два локуса находятся в разных парах гомологичных хромосом, и каждый ген представлен двумя аллелями.
Менделевский закон расщепления факторов можно объяснить теперь расхождением гомологичных хромосом, происходящим в анафазе I мейоза, и случайным распределением аллелей между гаметами. Эти события схематически представлены на рис. 24.6.

Рис. 24.6. Объяснение менделевского закона расщепления факторов (аллелей) А и а как результата расхождения гомологичных хромосом в мейозе.
24.2.1. Поведение хромосом как основа независимого распределения
Менделевский закон независимого распределения тоже можно объяснить особенностями перемещения хромосом во время мейоза. При образовании гамет распределение между ними аллелей из данной пары гомологичных хромосом происходит совершенно независимо от распределения аллелей из других пар (рис. 24.7). Именно случайное расположение гомологичных хромосом на экваторе веретена в метафазе I мейоза и их последующее расхождение в анафазе I ведет к разнообразию рекомбинаций аллелей в гаметах. Число возможных сочетаний аллелей в мужских или женских гаметах можно определить по общей формуле 2 n , где n — гаплоидное число хромосом. У человека n = 23, а возможное число различных сочетаний составляет 2 23 = 8 388 608.

Рис. 24.7. Объяснение менделевского закона независимого распределения факторов (аллелей) А, а, В, b как результата независимого расхождения разных пар гомологичных хромосом в мейозе (ср. с рис. 23.16).
24.5. Отложение крахмала в пыльцевых зернах кукурузы контролируется наличием одного из аллелей определенного гена. В присутствии другого аллеля отложения крахмала не происходит. Объясните, почему половина пыльцевых зерен, образующихся на данном растении кукурузы, содержит крахмал, а в другой половине зерен его нет.
24.6. Рассчитайте число возможных сочетаний хромосом в пыльцевых зернах шафрана (Crocus balansae), у которого диплоидное число хромосом равно шести (2n = 6).
Биологическая библиотека - материалы для студентов, учителей, учеников и их родителей.
Наш сайт не претендует на авторство размещенных материалов. Мы только конвертируем в удобный формат материалы, которые находятся в открытом доступе и присланные нашими посетителями.
Если вы являетесь обладателем авторского права на любой размещенный у нас материал и намерены удалить его или получить ссылки на место коммерческого размещения материалов, обратитесь для согласования к администратору сайта.
Разрешается копировать материалы с обязательной гипертекстовой ссылкой на сайт, будьте благодарными мы затратили много усилий чтобы привести информацию в удобный вид.
Расщепление генов происходит при гаметогенезе во время мей-отических делений. При мейозе хромосомы расходятся и проходят в разные гаметы, причем каждая гамета получает по одной хромосоме (гомологу) из пары хромосом. Независимое распределение генов также может быть объяснено поведением хромосом при мейозе (рис. 127).[ . ]
Как видно, расщепление и независимое перераспределение генов (рекомбинация) является причиной гигантского генетического разнообразия (полиморфизма) людей. В природе нет двух индивидуумов с одинаковым генотипом. Каждый человек несет специфический набор генов, вследствие чего каждый человек генетически индивидуален и неповторим. Исключение составляют лишь однояйцовые близнецы, для которых характерны одинаковые генотипы.[ . ]
Обнаружение расщепления признаков во втором поколении гибридных растений садового гороха имело огромное значение, ибо, исходя из данных о частоте расщепления, Г. Мендель раскрыл внутреннее содержание этого явления, сформулировав гипотезу о существовании факторов (единиц) наследственности, которая затем переросла в теорию гена.[ . ]
Цитологические основы расщепления генов (первого закона наследственности) определяются парностью хромосом, поведением хромосом диплоидных клеток при мейозе (спаривании и расхождении гомологичных хромосом) и последующим оплодотворением половых клеток, хромосомы которых несут по одному аллелю генных пар. У соматических клеток один аллель одной пары генов располагается на одном члене хромосомной пары, тогда как другой аллель представлен в другом члене хромосомной пары.[ . ]
По мере повышения числа генов, обусловливающих расщепление родительских форм по признаку длины початков от 1 до п, число генотипов Р2 возрастает от 3 до 3й. И наоборот, число генотипов, сходных с родительскими формами, снижается с 2(1/4) в Р2 до 2(1/4)п. Если допустить, что длина початков определяется только 10 генами, тогда Р2 включает З10, или 59 049 различных генотипов. Для восстановления каждого генотипа популяция должна составлять минимум 410, т. е. должно быть выращено 1 048 576 растений.[ . ]
Сегрегационный груз обусловлен сегрегацией (расщеплением) генов в соответствии с законами Менделя, проявляется вы-щеплением менее приспособленных гомозиготных особей при популяционном отборе в поль зу гетерозиготных особей, например как в случае с серповидно-клеточной анемией. Чаще всего это рецессивные мутации, передающиеся от родителей к детям в результате расщепления генов согласно законам Менделя. К таким признакам относятся наследственные дефекты ферментов — фенилкетонурия. галактоземия, муковисцидоз и др.[ . ]
Таким образом, не исключено, что для клонирования генов рестрикции-модификации, по меньшей мере в некоторых случаях, необходимо их вырезание из исходной ДНК в соответствующем окружении. Необходимость выполнения этого требования несомненно должна значительно снизить вероятность успешного клонирования таких генов и связана с необходимостью получения больших клонотек, желательно на основе фрагментов клонируемой ДНК, полученных путем ее расщепления наиболее случайным образом.[ . ]
Считают, что человек является самым гетерозиготным видом. В соответствии с законами расщепления и независимого перераспределения генов, открытыми Г. Менделем, одиночная половая клетка человека содержит лишь половину (гаплоидное число) хромосом и генов, представленных в соматических клетках организма в диплоидном наборе. Если, например, какой-либо индивидуум является гетерозиготным организмом по трем парам генов Аа, Вв и Сс, то его половые клетки несут лишь половину таких генов (А, В, С или а, в, с). Следовательно, потомству такого индивида будет передана также половина генов родительского организма, например, генов А, В и С. Между тем разные половые клетки одного и того же родителя несут разные наборы хромосом, а оплодотворение той или иной клетки является делом случая. Поэтому от одних и тех же родителей дети наследуют разные наборы генов как от одного, так и от другого родителя, вследствие чего братья и сестры всегда имеют разные генотипы, а следовательно и разные фенотипы. Одинаковые генотипы и фенотипы, как уже отмечено выше, присущи лишь монозиготным близнецам.[ . ]
Мейоз представляет логически необходимую часть жизненного цикла, размножающегося половым путем, Мейоз обеспечивает расщепление генов - отдельных участков ДНК по отдельным гаметам, в результате чего происходит разнообразное сочетание генов в гаметах. В отношении поддержания постоянства хромосом в клетке оплодотворение составляет антитезу (противоположное) мейозу, о процессе оплодотворения происходит слияние гаплоидных ядер двух разнополых гамет с образованием одной клетки - зиготы с диплоидным ядром.[ . ]
Итак, из результатов этих экспериментов следует, что каждый наследуемый признак находится под контролем пары одинаковых (аллельных) генов, из которых один аллель (один ген пары) происходит от одного родителя, тогда как другой аллель (другой ген пары) берет начало от другого родителя. Находясь в гибриде первого поколения, различные аллели не смешиваются и не влияют один на другой. Когда у зрелого организма наступает процесс образования половых клеток, то два аллеля генной пары расходятся (сегрегируют) чистыми и оказываются в разных гаметах, продуцируемых одним и тем же гибридом. Гаметы несут лишь по одному аллелю из каждой генной пары. Появление одного аллельного гена в той или иной гамете — дело случая. В процессе последующего оплодотворения женских половых клеток мужскими и образования гибридов второго поколения гены вновь собираются в пары. Заключение Г. Менделя, в соответствии с которым члены какой-либо генной пары расщепляются и расходятся в разные гаметы, известно под названием закона расщепления (сегрегации), или первого закона наследственности (первого закона Г. Менделя), и лежит в основе теории гена.[ . ]
Вегетативная нервная система — автономная нервная система, функционирующая независимо от воли и сознания, регулирующая деятельность органов и процессы обмена веществ; разделяется на симпатический и парасимпатический отделы Ген — единица наследственного материала, ответственная за формирование какого-либо признака организма; в генах закодирован состав всех синтезируемых организмом белков Геном — совокупность генов см., содержащихся в наборе хромосом см. Гипоксия — кислородное голодание, понижение содержания или парциального давления кислорода в атмосфере, организме или в тех или иных его органах и тканях Гликогенолиз — расщепление гликогена ферментом фосфорилазой см.[ . ]
Генетический анализ в принципе подобен химическому анализу, задача которого заключается в разложении сложных химических соединений на более простые компоненты. Однако в отличие от химического анализа, например нуклеопротеидов, расщепление которых на структурные части основано на гидролизе, классический генетический анализ основывается на расщеплении (сегрегации) и рекомбинации генов в мейозе и осуществляется путем скрещиваний особей с разными признаками и учета результатов скрещиваний.[ . ]
Наконец, в антропогенезе действовали факторы, связанные с возникновением сельского хозяйства. Например, введение в питание молока вело к отбору индивидов, обладающих способностью утилизировать (расщеплять) лактозу. Известно, что в популяциях современных людей, не использующих молоко, концентрация генов, контролирующих расщепление лактозы, ничтожно мала.[ . ]
Эти методы направлены на создание организмов, способных разлагать только одно или два определенных соединения. Недавнее открытие того, что лигниназные системы способны катализировать разложение галогенированных субстратов, может указать путь к использованию очень низкоспецифичных ферментных систем [703]. Подразумевается сшивание генов низкоспецифичных ферментов, которые катализируют первые этапы разложения ароматических соединений, а также способны катализировать трансформацию их галогенированных аналогов, с генами модифицированного пути расщепления кольца, например у Pseudomonas В13. В сущности, будет получен организм, способный разлагать целый ряд моно- и дигалогенированных ароматических соединений (рис. 10.7).[ . ]
Многие структурные и ферментные белки содержат две или более субъединиц (мономеров), соединенных между собой иековалентными связями. Расщепление фермента на субъединицы лишает его активности или резко снижает ее. Из субъединиц построены и сократительные белки мышц — миозин и актин: первый — из пяти субъединиц, а второй — из очень большого числа глобулярных молекул й-актина. Расщепление этих сложных структур высоким давлением и препятствие сборке субъединиц О-актина в фибриллярный Е-актин делает невозможным взаимодействие между этими белками и мышечное сокращение. Наконец, высокое давление изменяет структуры клеточных мембран и уменьшает их проницаемость.[ . ]
Значительная изменчивость ката-болических плазмид в этом роде объясняет широкие катаболические возможности, которыми обладают его представители. Физический размер этих плазмид позволяет им кодировать большое количество генов. Плазмида длиной 150 тыс. п. н. содержит ДНК в количестве, достаточном для кодирования приблизительно 150 генов.[ . ]
Имея в виду соображения, приведенные выше, вернемся к гибридам Р1, которые являются гетерозиготами Иг и продуцируют гаметы К и г в равных количествах. Допуская, что при оплодотворении гаметы объединяются случайно (нет преимуществ для объединения гамет К и Я или гиг, или И и г) и что все формирующиеся гибридные растения жизнеспособны, можно предположить, что в результате расщепления генов формируемые гибриды будут различными как по фенотипу, так и по генотипу (см. рис. 124).[ . ]
Ряд ученых на основании современных цитогенетических исследований считают, что потенции к развитию зиготы в известной степени зависят от градиентов эндоплазматических материалов и неоднородности эктоплазмы самой зиготы. Это явление (4. Д. Кравченко определяет как градиентное отношение в онтогении. Клетки, получающие во время дробления зиготы нехромосомный материал, формируют впоследствии различные ткани зародыша. Расщепление нехромосомного материала во время дробления зиготы—первая стадия дифференциации, и только позднее причиной ее становятся изменения ядериого материала. Гены активно включаются в процесс развития, начиная с фазы гасгруляции, создающей химическую дифференциацию зародыша. В дальнейшем любые изменения, происходящие в формирующемся организме, обусловливаются генетическими факторами.[ . ]
Четкий пример группового отбора описан Левонтичом (1962), который провел машинное моделирование судьбы трех стерильных ¿-аллелей в популяциях домовых мышей. Каждый из этих аллелей в гомозиготном состоянии обусловливает мужскую стерильность, однако среди самцов, гетерозиготных по тому или иному аллелю, соответствующий аллель содержится в сперме 85—99% самцов, а не 50%, как следовало ожидать. Точная частота передачи зависит от конкретного аллеля. В больших популяциях частота аллеля должна стабилизироваться на уровне, при котором скорость выпадения в гомозиготность уравновешивается его замещением при аберрантном коэффициенте расщепления у гетерозиготных самцов. Этот уровень q=2m—1, где т — коэффициент передачи. Для аллеля tw2 с /и=0,86 и теоретическим ¿7 = 0,70 в одной популяции было получено ¿7 = 0,37. Левонтин нашел, что при машинном моделировании можно получить приближение для наблюдавшейся частоты гена, если принять такую структуру популяции, при которой размножение происходит в небольших семейных группах, состоящих из двух самцов и шести самок. В 70% семей i-аллель закрепляется либо у всех особей, либо хотя бы у самцов и вызывает вымирание семьи. Однако в 30% семей i-аллель утрачивается в результате дрейфа генов, и все популяции стабилизируются после 24 поколений. Таким образом, ¿-аллель был бы элиминирован, если бы миграции не заносили его вновь в некоторые свободные от этих аллелей популяции; реальная частота любого аллеля в популяции зависит от вероятности вымирания несущих его групп и от частоты его заноса при миграции в свободные от этого аллеля семейные группы.[ . ]
На примере львиного зева (АпйггЫпшп та и8 Ь.) исследовали связь между окраской цветка и количественными признаками. Это классический объект генетических исследований, он характеризуется исключительным разнообразием расцветок - от чисто белого до темно-пурпурового. По окраске и форме цветка для вида характерно неполное доминирование одной из двух пар аллелей. Красная окраска цветка определяется доминантной аллелью с неполным доминированием над рецессивной аллелью, вызывающей белую окраску. В гибридах Р получается промежуточная розовая окраска. Неполное доминирование при дигибридном скрещивании по одной или обеим парам аллелей генов видоизменяет расщепление в Р2 по фенотипу. Природа неполного доминирования недостаточно изучена. Исходная форма данного вида имеет розовую окраску, которая под влиянием внешних факторов переходит в малиновую с одной стороны и в красную - с другой. Под влиянием солнечной инсоляции, температуры, влажности и других факторов, вызывающих смешение пигментов и изменение pH клеточного сока, цветок может приобретать разную окраску. По окраске цветков популяцию разбили на десять дифференцированных групп, способных переходить в соседние классы. Исследовали среднюю высоту, число цветков, размеры органов у растений этих групп. Выборки брали в разные годы из популяции, состоящей из растений с разной окраской цветков (см. таблицу).[ . ]
Вторичное мейотическое деление у человека наступает, когда вторичный овоцит (развивающаяся яйцеклетка) проходит из яичника в фаллопиеву трубу. Однако это деление не завершается до тех пор, пока во вторичный овоцит не проникает ядерное содержимое сперматозоида, что происходит обычно в фаллопиевой трубе. Когда ядро сперматозоида проникает во вторичный овоцит, то последний делится, в результате чего образуется овотида (зрелая яйцеклетка) с пронуклеусом, содержащим одиночный набор из 23 материнских хромосом. У некоторых других видов образуются яйцеклетки, определяющие как мужской пол, так и женский. Важно подчеркнуть, что здесь происходит также расщепление и рекомбинация генов, основу которой создает расхождение хромосом. Другая клетка, образующаяся в результате второго мейотического деления у человека, является вторым полярным тельцем, не способным к дальнейшему развитию. В это время подвергается делению надвое и полярное (редукционное) тельце. Таким образом, развитие одного овоцита первого порядка сопровождается образованием одной овотиды и трех редукционных телец. В яичниках таким путем на протяжении жизни обычно созревает 300-400 ово-цитов, но в месяц созревает лишь один овоцит. В течение дифференциации яйцеклеток формируются мембраны, уменьшается в размере их ядро.[ . ]
Концепция данной теории заключается в том, что передача наследственной информации в ряду поколений осуществляется путем передачи хромосом, в которых в определенной линейной последовательности расположены гены.
Данная теория была сформулирована в начале XX века. Значительный вклад в ее развитие внес американский генетик Томас Морган.
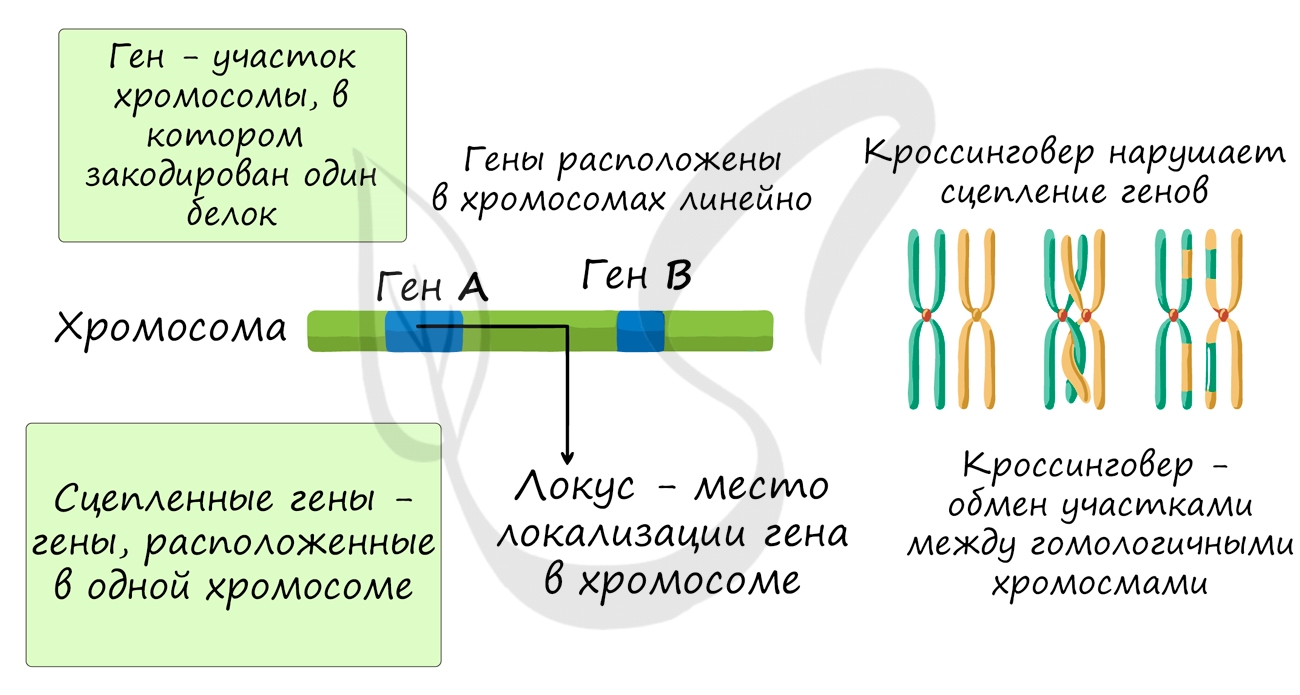
- Гены расположены в хромосомах в линейном порядке
- Каждый ген занимает в хромосоме определенное место - локус
- Гены, расположенные в одной хромосоме, образуют группу сцепления
- Сцепление генов может нарушаться в результате кроссинговера
- Частота кроссинговера между генами прямо пропорциональна расстоянию между ними
- Расстояние между генами измеряется в морганидах (1 морганида - 1% кроссинговера)
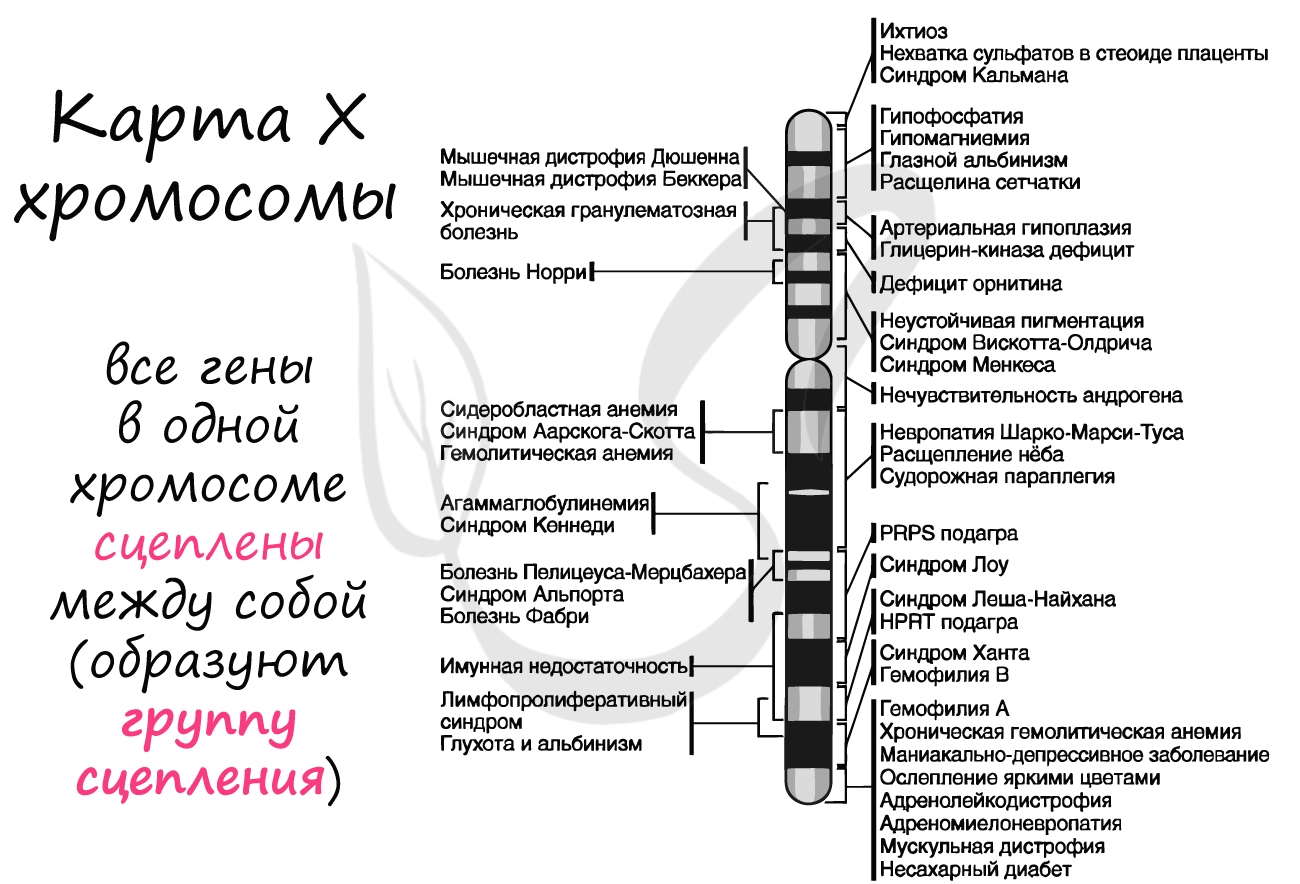
Группы сцепления
В предыдущей статье были раскрыты суть и применение в задачах III закона Менделя, закона независимого наследования, в основе которого лежат гены, расположенные в разных хромосомах. Но что если гены лежат в одной хромосоме? Такие гены образуют группу сцепления, в этом случае говорят о сцепленном наследовании.
Группа сцепления - совокупность всех генов, расположенных в одной хромосоме, вследствие чего они наследуются совместно. Число групп сцепления равно гаплоидному набору хромосом: у женщины 23 группы сцепления (23 пара - половые хромосомы XX), а у мужчины - 24 группы сцепления (X и Y представляют собой две отдельные группы).
Сцепление генов
Томас Морган в своих экспериментах изучал наследование признаков плодовых мушек дрозофил: серый (A) - черный (a) цвет тела, длинные (B) - зачаточные (b) крылья. В первом эксперименте Морган скрестил чистые линии плодовых мушек: серых с длинными крыльями (AABB) и черных с зачаточными (aabb).
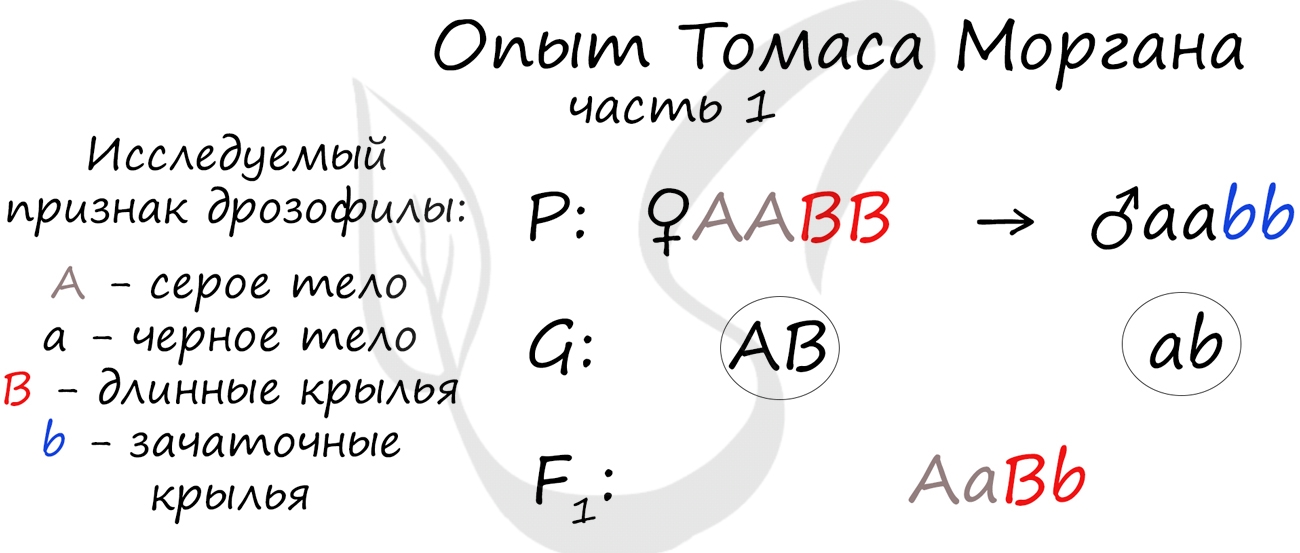
Только что вы видели первый закон Менделя (единообразия) в действии, правда, в несколько ином варианте - при дигибридном скрещивании. Но суть та же: в первом поколении все особи получаются единообразны по исследуемому признаку, с генотипом AaBb - с серым телом и длинными крыльями.
Далее Морган применил анализирующее скрещивание. Полученную в первом поколении дигетерозиготу (AaBb) он скрестил с черной особью с зачаточными крыльями (aabb). Результат весьма удивил Моргана и его коллег: помимо потомства с ожидаемыми фенотипами (серое тело + длинные крылья, черное тело + зачаточные крылья) были получены особи со смешанными признаками.

Потомство со смешанными признаками подразумевает под собой особи Aabb (серое тело + зачаточные крылья) и aaBb (черные тело + длинные крылья). Но откуда они могли взяться, если гены A и B находятся в одной хромосоме? Значит, образовались еще какие-то дополнительные гаметы, помимо AB и ab?
Объясняя полученные в потомстве фенотипы, которые содержали смешанные признаки, Томас Морган пришел к выводу, что между гомологичными хромосомами произошел кроссинговер, в результате которого образовались гаметы Ab, aB - кроссоверные гаметы.
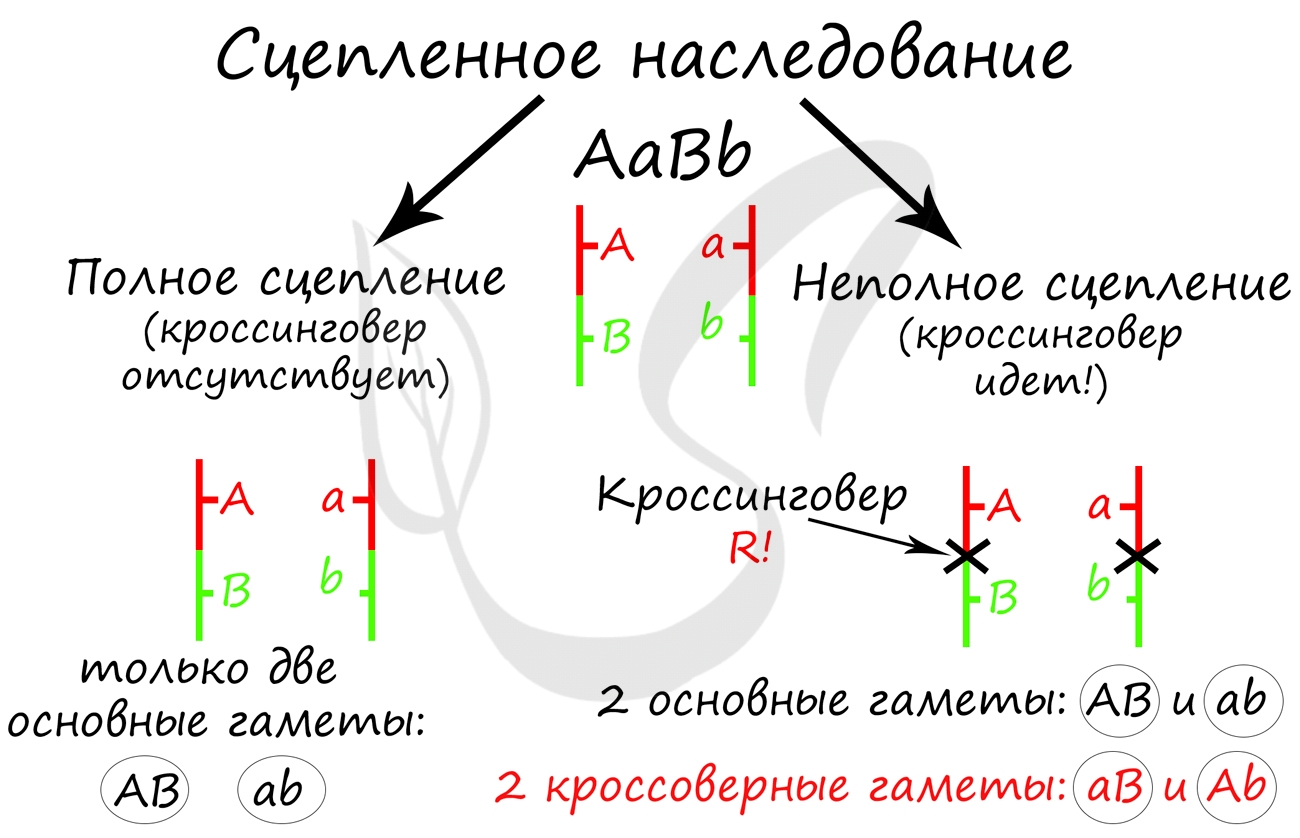
Очевидно, что в данном случае расстояние между генами A и B было 17 морганид, так как каждой кроссоверной гаметы (соответственно и особей) образовалось по 8.5%. Не забывайте, что процент кроссинговера равен расстоянию между генами. Поскольку расстояние было 17 морганид = 17%, то на каждую из кроссоверных гамет приходится половина - 8.5%
Пример решения генетической задачи №1
"Катаракта и полидактилия у человека обусловлены доминантными аутосомными генами, расположенными в одной хромосоме. Гены полностью сцеплены. Какова вероятность родить здорового ребенка в семье, где муж нормален, жена гетерозиготна по обоим признакам, мать жены также страдала обеими аномалиями, а отец был нормален".
Очень важно обратить внимание на то, что "гены полностью сцеплены" - это говорит об отсутствии кроссинговера, и то, что мы заметили это, обеспечивает верное решение задачи.

Самое главное, что вам следует усвоить: поскольку гены полностью сцеплены (кроссинговер отсутствует), женщина с генотипом AaBb может образовать только два типа гамет - AB, ab. Кроссоверные гаметы (Ab, aB) не образуются. Всего возможных генотипов потомков получается два, из которых здоров только один - aabb. Шанс родить здорового ребенка в такой семье ½ (50%).
Пример решения генетической задачи №2
"Гены доминантных признаков катаракты и эллиптоцитоза локализованы в 1-й аутосоме. Гены неполностью сцеплены. Женщина, болеющая катарактой и эллиптоцитозом, отец которой был здоров, выходит замуж за здорового мужчину. Определите возможные фенотипы потомства и вероятность рождения больного обеими аномалиями ребенка в этой семье".
Ключевые слова в тексте этой задачи, на которые следует обратить внимание: "гены неполностью сцеплены". Это означает, что между ними происходит кроссинговер.
Генотип женщины остается неясен из текста задачи. Раз она больна, то он может быть: AaBb, AABB, AABb, AaBB. Однако в тексте дано то, что развеет сомнения: "отец которой был здоров". Если ее отец был здоров, то его генотип был aabb, значит он передал дочери гамету ab. Теперь становится очевидно, что генотип дочери AaBb - она дигетерозиготна.

В данном случае между генами A и B произошел кроссинговер, их сцепление нарушилось. В результате образовались кроссоверные гаметы Ab, aB - которые привели к образованию особей с со смешанными признаками (Aabb, aaBb). Вероятность рождения в этой семье ребенка, больного обеими аномалиями, составляет ¼ (25%).
Наследование, сцепленное с полом
Половые хромосомы X и Y определяют пол человека. Генотип XX характерен для женщин, а XY - для мужчин. Мужская Y-хромосома не содержит аллелей многих генов, которые есть в X-хромосоме, вследствие этого наследственными заболеваниями, сцепленными с полом, чаще болеют мужчины.
Природа, несомненно, бережет женских особей. Женщины имеют две гомологичные хромосомы XX, и если ген наследственного заболевания попал в одну из X-хромосом, то чаще всего в другой X-хромосоме окажется "здоровый" ген, доминантный, которой подавит действие рецессивного гена. С генетической точки зрения, женщина будет носительницей заболевания, может его передать по поколению, но сама болеть не будет.
У мужчин если ген заболевания оказался в X-хромосоме, то не проявиться он не может. Именно по этой причине мужчины чаще страдают дальтонизмом, гемофилией и т.д.
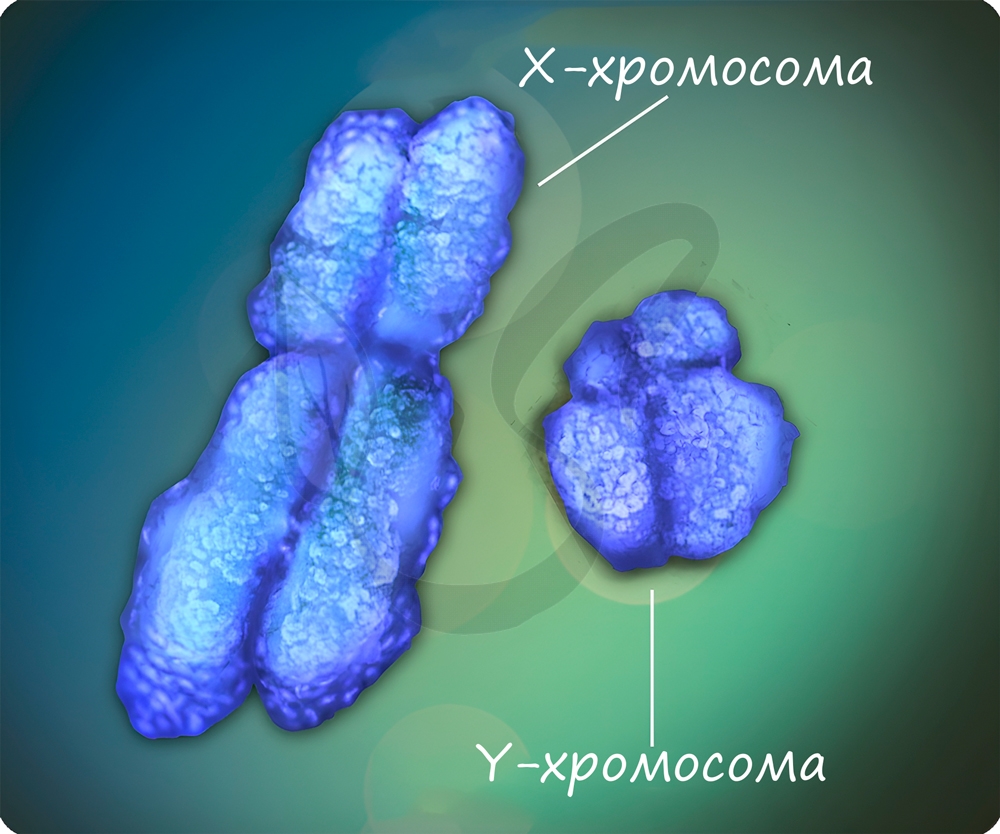
Не у всех организмов особь мужского пола характеризуется набором хромосом XY, а женского - XX. У пресмыкающихся, птиц, бабочек женские особи имеют гетерогаметный пол- XY, а мужские - XX. То же самое относится к домашним курам: петух - XX, курица - XY.
Решим несколько задач по теме наследования, сцепленного с полом. Речь в них будет идти о сцепленных с полом признаками - признаками, гены которых лежат не в аутосомах, а в гетеросомах (половых хромосомах).
Пример решения генетической задачи №3
"Рецессивный ген дальтонизма располагается в X-хромосоме. Женщина с нормальным зрением (отец был дальтоник) выходит замуж за мужчину с нормальным зрением, отец которого был дальтоником. Определите возможные фенотипы потомства".
Подробности о родословной важны и помогают заполнить белые пятна. Если отец женщины был дальтоником (X d Y), то очевидно, что он передал ей хромосому X d , так как от отца дочери всегда передается X-хромосома. Значит женщина гетерозиготна по данному признаку, а у мужчины возможен лишь один вариант здорового генотипа - X D Y. То, что его отец был дальтоником несущественно, ведь отец всегда передает сыну Y-хромосому.

- X D X D , X D X d - фенотипически здоровые девочки
- X D Y - здоровый мальчик
- X d Y - мальчик, который болен дальтонизмом
Пример решения генетической задачи №4
"Гипоплазия зубной эмали наследуется как сцепленный с X-хромосомой доминантный признак, шестипалость - как аутосомно-доминантный. В семье, где мать шестипалая, а у отца гипоплазия, родился пятипалый здоровый мальчик. Напишите генотипы всех членов семьи по данным признакам. Возможно ли у них рождение ребенка с двумя аномалиями одновременно?"
Ответ на вопрос: "Каковы генотипы матери и отца?" - лежат в потомстве. Пятипалый здоровый мальчик имеет генотип aaX b Y. Чтобы сформировался такой генотип, от матери должна прийти гамета aX b , а от отца - aY. Выходит, что единственно возможный генотип матери - AaX b X b , а генотип отца - aaX B Y.

Рождение ребенка с двумя аномалиями возможно - AaX B X b , вероятность такого события ¼ (25%).
Пример решения генетической задачи №5
"Рецессивные гены, кодирующие признаки дальтонизма и гемофилии, сцеплены с X-хромосомой. Мужчина с нормальным цветовым зрением и гемофилией женится на здоровой женщине, отец которой был дальтоником, но не гемофиликом. Известно, что мать женщины была гомозиготна по исследуемым признакам. Какое потомство получится от брака их дочери со здоровым мужчиной?"
Генотип мужчины вопросов не вызывает, так как единственный возможный вариант - X hD Y. Генотип женщины дает возможность узнать ее отец (X Hd Y), который передал ей гамету X Hd (отец всегда передает дочке X хромосому, а сыну - Y), следовательно, ее генотип - X HD X Hd
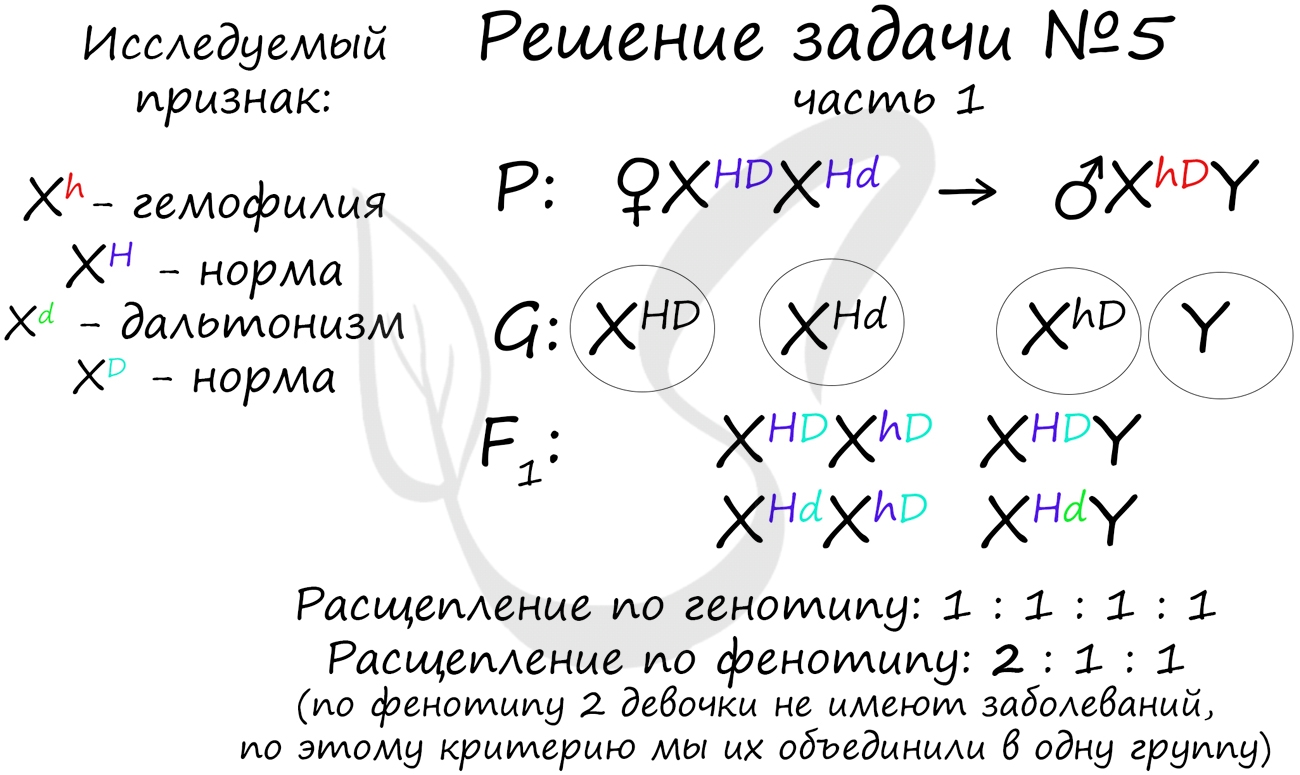
Как оказалось, возможны два варианта генотипа дочери: X HD X hD , X Hd X hD . Генотип здорового мужчины X HD Y. Следуя логике задачи, мы рассмотрим два возможных варианта брака.
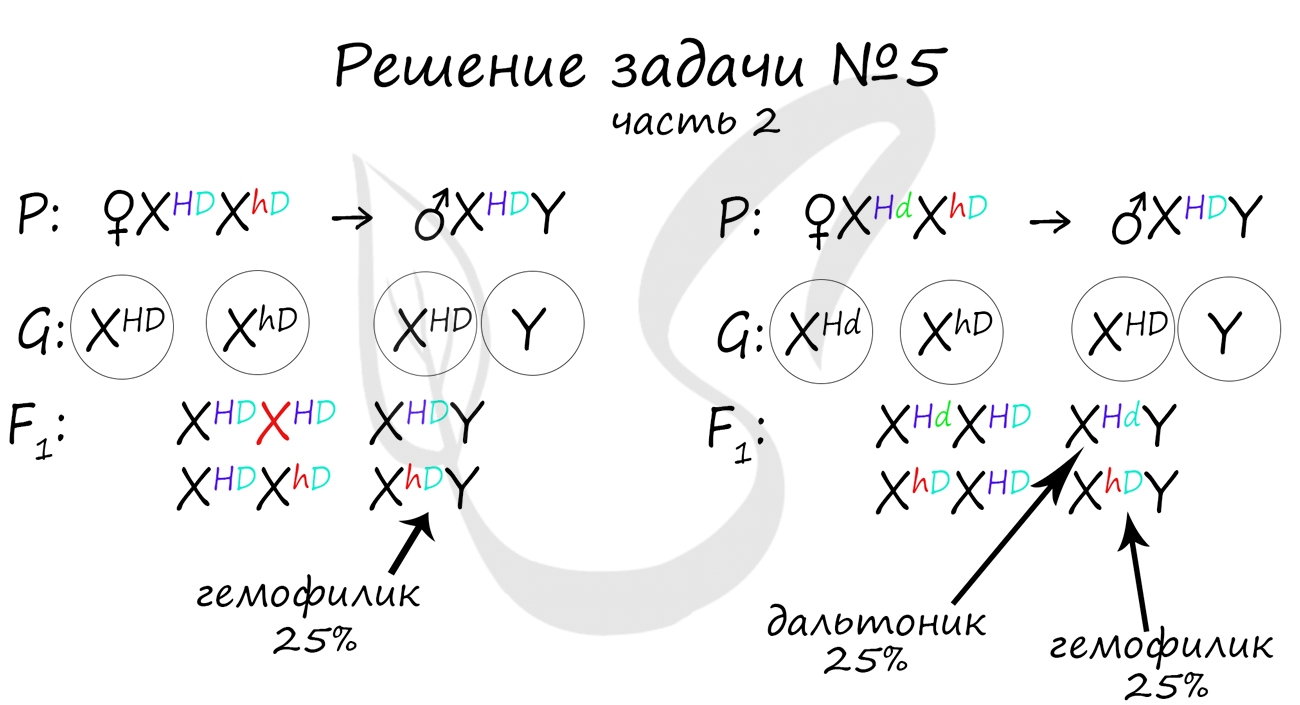
Не забывайте, что на экзамене схема задачи не является ответом. Ответ начинается только после того, как вы напишите слово "Ответ: . ". В ответе должны быть указаны все фенотипы потомства, их описание, что возможно покажется рутинными при большом числе потомков, но весьма приятным, если вы верно решили задачу и получили за нее заслуженные баллы :)
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Читайте также:
- Примеры модернизма из школьной программы
- Особенности воспитания детей в городской школе
- Операционные системы выясните достоинства и недостатки самых популярных ос кратко
- Высшая школа экономики мировой рейтинг
- Организация проведения капитального ремонта общего имущества в многоквартирных домах кратко