
Описание развития зиготы кратко
Обновлено: 04.07.2024
Эмбриональное развитие-это сложный и длительный морфогенетический процесс, в ходе которого из отцовской и материнской половых клеток формируется новый многоклеточный организм, способный к самостоятельной жизнедеятельности в условиях внешней среды. Лежит в основе полового размножения и обеспечивает передачу наследственных признаков от родителей потомкам.
Оплодотворение,заключается в соединении спермотозоида с яйцом.Важнейшими этапами процесса оплодотворения является:
1)проникновение СП в яйцо;
2)активация в яйце разнообразных синтетических процессов ;
3)слияние ядер яйца и СП с восстановлением диплоидного набора хромосом.
Для того ,чтобы оплодотворен произошло, необходимо сближение женских и мужских половых клеток. Оно достигается благодаря осеменению.
Проникновению СП в яйцо способствует фермент гиалуронидиаза и другие биологические активн.вещества (спермолизин),которые повышают проницаемость основного межклеточного вещества.Ферменты выделяют акросомой в процессе акросомной реакции. Сущность ее заключается в следующем,в момент контакта с яйцеклеткой на вершине головки спермия плазматическая мембрана и прилежащая к ней мембрана растворяется, и происходит растворение прилежащего участка яйцевой оболочки.Акросомная мембрана выпячивается наружу и образует вырост в виде полой трубки.В области такого контакта возникает выпячивание или бугорок оплодотворения.ювслед за чем плазмат.мембраны обеих гамет сливаются и начинается объединение их содержимого.С этого момента СП и Я представляет собой единую клетку-зиготу.
Активация Я или кортикальная реакция ,развивающаяся в результате контакта со СП,имеет морфологические и биохимические проявления. Проявлением активации служат изменения поверхностного кортикального слоя ооплазмы и образование оболочки оплодотворения. Оболочка оплодотворения защищает яцйо от проникновения сверчисленных спермиев.
Зигота- диплоидная (содержащая полный двойной набор хромосом) клетка, образующаяся в результате оплодотворения (слияния яйцеклетки и сперматозоида). Зигота является тотипотентной (то есть, способной породить любую другую) клеткой.
У человека первое митотическое деление зиготы происходит спустя приблизительно 30 часов после оплодотворения, что обусловлено сложными процессами подготовки к первому делению дробления.
Дробление — это ряд последовательных митотических делений зиготы и заканчивающихся образованием многоклеточного зародыша — бластулы. Первое деление дробления начинается после объединения наследственного материала пронуклеусов и образования общей метафазной пластинки. Возникающие при дроблении клетки называют бластомерами (от греч. бласте—росток, зачаток). Особенностью митотических делений дробления является то, что с каждым делением клетки становятся все мельче и мельче, пока не достигнут обычного для соматических клеток соотношения объемов ядра и цитоплазмы.Сначала бластомеры прилегают друг к другу, образуя скопление клеток, называемое морулой. Затем между клетками образуется полость — бластоцель, заполненная жидкостью. Клетки оттесняются к периферии, образуя стенку бластулы — бластодерму.Общий размер зародыша к концу дробления на стадии бластулы не превышает размера зиготы.
Прогенез – гаметогенез (спермато– и овогенез) и оплодотворение.Сперматогенез осуществляется в извитых канальцах семенников и подразделяется на четыре периода: 1) период размножения – I; 2) период роста – II; 3) период созревания – III; 4) период формирования – IV. Овогенез осуществляется в яичниках и подразделяется на три периода:1) период размножения (в эмбриогенезе и в течение 1-го года постэмбрионального развития); 2) период роста (малого и большого); 3) период созревания.Яйцеклетка состоит из ядра с гаплоидным набором хромосом и выраженной цитоплазмы, в которой содержатся все органеллы, за исключением цитоцентра.
Дробление. Характеристика дробления. Основные типы яйцеклеток по расположению желтка. Связь строения яйцеклетки с типом дробления. Бластомеры и эмбриональные клетки. Строение и типы бластул.
Дробление —В основе этого процесса лежит митотическое деление клеток. Однако образующиеся в результате деления дочерние клетки не расходятся, а остаются тесно прилегающими друг к другу. В процессе дробления дочерние клетки прогрессивно уменьшаются. Каждому животному свойствен определенный тип дробления, обусловленный количеством и характером распределения желтка в яйцеклетке. Желток тормозит дробление, поэтому часть зиготы, перегруженная желтком, дробится медленнее или не дробится вовсе.

В изолецитальном, бедном желтком оплодотворенном яйце ланцетника, первая борозда дробления в виде щели начинается на анимальном полюсе и постепенно распространяется в продольном меридиональном направлении к вегетативному, разделяя яйцо на 2 клетки — 2 бластомера. Вторая борозда проходит перпендикулярно первой — образуются 4 бластомера. В результате ряда последовательных дроблений формируются группы клеток, тесно прилегающих друг к другу. У некоторых животных такой зародыш напоминает ягоду шелковицы или малины. Он получил название морулы (лат. morum — тутовая ягода) – многоклеточного шара без полости внутри.
В телолецитальных яйцах, перегруженных желтком — дробление может быть полным равномерным или неравномерным и неполным. Бластомеры вегетативного полюса из-за обилия инертного желтка всегда отстают в темпе дробления от бластомеров анимального полюса. Полное, но неравномерное дробление характерно для яиц амфибий. У рыб, птиц и некоторых других животных дробится лишь часть яйца, расположенная на анимальном полюсе; происходит неполное дискоидальное дробление.В процессе дробления увеличивается число бластомеров, однако бластомеры не вырастают до размеров исходной клетки, а с каждым дроблением становятся мельче. Это объясняется тем, что митотические циклы дробящейся зиготы не имеют типичной интерфазы; пресинтетический период (G1) отсутствует, а синтетический (S) начинается еще в телофазе предшествующего митоза.
Дробление яйца заканчивается образованием бластулы.
В полилецитальных яйцеклетках костистых рыб, пресмыкающихся, птиц, а также однопроходных млекопитающих дробление частичное, или мероб-ластическое, т.е. охватывает только свободную от желтка цитоплазму. Она располагается в виде тонкого диска на анимальном полюсе, поэтому такой тип дробления называют дискоидальным. При характеристике типа дробления учитывают также взаимное расположение и скорость деления бластомеров. Если бластомеры располагаются рядами друг над другом по радиусам, дробление называют радиальным.
Дробление может быть: детерминированным и регулятивным; полным (голобластическим) или неполным (меробластическим); равномерным (бластомеры более-менее одинаковы по величине) и неравномерным (бластомеры не одинаковы по величине, выделяются две — три размерные группы, обычно называемые макро- и микромерами)
Типы яйцеклеток:
Количество желтка- Олиголецетальные (ланцетник) Мезолецетальные (амфибии) Полилецетальные (рыбы, птицы)
Месторасположение-Изолецетальные (расположен диффузно, равномерно) . Содержат немного желтка, равномерно расположенного по всей клетке. Характерны для иглокожих, низших хордовый и млекопитающих. У млекопитающих — это аллециатльные яйцеклетки (желтка практически нет)
Телолецетальные ( с умеренным количествам желтка на нижнем вегетативном полюсе)
Резко телолецетальные (с большим количествам желтка, занимает всю яйцеклетку, кроме верхнего полюса. Желтка много, сконцентрирован на вегетативном полюсе. Выделяют 2 группы: умеренно телолецитальные (молюски, земноводные) и резколецитальные (рептили и птицы). На анамальном полюсе сосредоточены цитоплазма и ядро.
Центролецетальные(желтка немного, но плотно в центре). Желтка немного, расположен в центре. Характерно для членистоногих
Бластомеры- клетки, образующиеся в результате делений дробления яйца у многоклеточных животных. Характерная особенность Б.— отсутствие роста в период между делениями, вследствие чего при очередном делении объём каждого Б. уменьшается вдвое. При голобластич. дроблении в телолецитальных яйцах Б. различаются по размерам: крупные Б.— макромеры, средние — мезомеры, мелкие — микромеры. Во время синхронных делений дробления Б., как правило, однородны по форме, структура их цитоплазмы очень проста. Затем поверхностные Б. уплощаются, и яйцо переходит к заключит, фазе дробления — бластуляции.
Строение бластулы. Если образуется сплошной шар без полости внутри, то такой зародыш называют морулой. Образование бластулы или морулы зависит от свойств цитоплазмы. Бластула образуется при достаточной вязкости цитоплазмы, морула – при слабой вязкости. При достаточной вязкости цитоплазмы бластомеры сохраняют округлую форму и только в местах соприкосновения слегка сплющиваются. Вследствие этого между ними появляется щель, которая по мере дробления увеличивается, заполняется жидкостью и превращается в бластоцель. При слабой вязкости цитоплазмы бластомеры не округляются и располагаются тесно друг возле друга, щели нет и полость не образуется. Бластулы различны по своему строению и зависят от типа дробления.
Эмбриональное развитие-это сложный и длительный морфогенетический процесс, в ходе которого из отцовской и материнской половых клеток формируется новый многоклеточный организм, способный к самостоятельной жизнедеятельности в условиях внешней среды. Лежит в основе полового размножения и обеспечивает передачу наследственных признаков от родителей потомкам.
Оплодотворение,заключается в соединении спермотозоида с яйцом.Важнейшими этапами процесса оплодотворения является:
1)проникновение СП в яйцо;
2)активация в яйце разнообразных синтетических процессов ;
3)слияние ядер яйца и СП с восстановлением диплоидного набора хромосом.
Для того ,чтобы оплодотворен произошло, необходимо сближение женских и мужских половых клеток. Оно достигается благодаря осеменению.
Проникновению СП в яйцо способствует фермент гиалуронидиаза и другие биологические активн.вещества (спермолизин),которые повышают проницаемость основного межклеточного вещества.Ферменты выделяют акросомой в процессе акросомной реакции. Сущность ее заключается в следующем,в момент контакта с яйцеклеткой на вершине головки спермия плазматическая мембрана и прилежащая к ней мембрана растворяется, и происходит растворение прилежащего участка яйцевой оболочки.Акросомная мембрана выпячивается наружу и образует вырост в виде полой трубки.В области такого контакта возникает выпячивание или бугорок оплодотворения.ювслед за чем плазмат.мембраны обеих гамет сливаются и начинается объединение их содержимого.С этого момента СП и Я представляет собой единую клетку-зиготу.
Активация Я или кортикальная реакция ,развивающаяся в результате контакта со СП,имеет морфологические и биохимические проявления. Проявлением активации служат изменения поверхностного кортикального слоя ооплазмы и образование оболочки оплодотворения. Оболочка оплодотворения защищает яцйо от проникновения сверчисленных спермиев.
Зигота- диплоидная (содержащая полный двойной набор хромосом) клетка, образующаяся в результате оплодотворения (слияния яйцеклетки и сперматозоида). Зигота является тотипотентной (то есть, способной породить любую другую) клеткой.
У человека первое митотическое деление зиготы происходит спустя приблизительно 30 часов после оплодотворения, что обусловлено сложными процессами подготовки к первому делению дробления.
Дробление — это ряд последовательных митотических делений зиготы и заканчивающихся образованием многоклеточного зародыша — бластулы. Первое деление дробления начинается после объединения наследственного материала пронуклеусов и образования общей метафазной пластинки. Возникающие при дроблении клетки называют бластомерами (от греч. бласте—росток, зачаток). Особенностью митотических делений дробления является то, что с каждым делением клетки становятся все мельче и мельче, пока не достигнут обычного для соматических клеток соотношения объемов ядра и цитоплазмы.Сначала бластомеры прилегают друг к другу, образуя скопление клеток, называемое морулой. Затем между клетками образуется полость — бластоцель, заполненная жидкостью. Клетки оттесняются к периферии, образуя стенку бластулы — бластодерму.Общий размер зародыша к концу дробления на стадии бластулы не превышает размера зиготы.
Прогенез – гаметогенез (спермато– и овогенез) и оплодотворение.Сперматогенез осуществляется в извитых канальцах семенников и подразделяется на четыре периода: 1) период размножения – I; 2) период роста – II; 3) период созревания – III; 4) период формирования – IV. Овогенез осуществляется в яичниках и подразделяется на три периода:1) период размножения (в эмбриогенезе и в течение 1-го года постэмбрионального развития); 2) период роста (малого и большого); 3) период созревания.Яйцеклетка состоит из ядра с гаплоидным набором хромосом и выраженной цитоплазмы, в которой содержатся все органеллы, за исключением цитоцентра.
Дробление. Характеристика дробления. Основные типы яйцеклеток по расположению желтка. Связь строения яйцеклетки с типом дробления. Бластомеры и эмбриональные клетки. Строение и типы бластул.
Дробление —В основе этого процесса лежит митотическое деление клеток. Однако образующиеся в результате деления дочерние клетки не расходятся, а остаются тесно прилегающими друг к другу. В процессе дробления дочерние клетки прогрессивно уменьшаются. Каждому животному свойствен определенный тип дробления, обусловленный количеством и характером распределения желтка в яйцеклетке. Желток тормозит дробление, поэтому часть зиготы, перегруженная желтком, дробится медленнее или не дробится вовсе.
В изолецитальном, бедном желтком оплодотворенном яйце ланцетника, первая борозда дробления в виде щели начинается на анимальном полюсе и постепенно распространяется в продольном меридиональном направлении к вегетативному, разделяя яйцо на 2 клетки — 2 бластомера. Вторая борозда проходит перпендикулярно первой — образуются 4 бластомера. В результате ряда последовательных дроблений формируются группы клеток, тесно прилегающих друг к другу. У некоторых животных такой зародыш напоминает ягоду шелковицы или малины. Он получил название морулы (лат. morum — тутовая ягода) – многоклеточного шара без полости внутри.
В телолецитальных яйцах, перегруженных желтком — дробление может быть полным равномерным или неравномерным и неполным. Бластомеры вегетативного полюса из-за обилия инертного желтка всегда отстают в темпе дробления от бластомеров анимального полюса. Полное, но неравномерное дробление характерно для яиц амфибий. У рыб, птиц и некоторых других животных дробится лишь часть яйца, расположенная на анимальном полюсе; происходит неполное дискоидальное дробление.В процессе дробления увеличивается число бластомеров, однако бластомеры не вырастают до размеров исходной клетки, а с каждым дроблением становятся мельче. Это объясняется тем, что митотические циклы дробящейся зиготы не имеют типичной интерфазы; пресинтетический период (G1) отсутствует, а синтетический (S) начинается еще в телофазе предшествующего митоза.
Дробление яйца заканчивается образованием бластулы.
В полилецитальных яйцеклетках костистых рыб, пресмыкающихся, птиц, а также однопроходных млекопитающих дробление частичное, или мероб-ластическое, т.е. охватывает только свободную от желтка цитоплазму. Она располагается в виде тонкого диска на анимальном полюсе, поэтому такой тип дробления называют дискоидальным. При характеристике типа дробления учитывают также взаимное расположение и скорость деления бластомеров. Если бластомеры располагаются рядами друг над другом по радиусам, дробление называют радиальным.
Дробление может быть: детерминированным и регулятивным; полным (голобластическим) или неполным (меробластическим); равномерным (бластомеры более-менее одинаковы по величине) и неравномерным (бластомеры не одинаковы по величине, выделяются две — три размерные группы, обычно называемые макро- и микромерами)
Типы яйцеклеток:
Количество желтка- Олиголецетальные (ланцетник) Мезолецетальные (амфибии) Полилецетальные (рыбы, птицы)
Месторасположение-Изолецетальные (расположен диффузно, равномерно) . Содержат немного желтка, равномерно расположенного по всей клетке. Характерны для иглокожих, низших хордовый и млекопитающих. У млекопитающих — это аллециатльные яйцеклетки (желтка практически нет)
Телолецетальные ( с умеренным количествам желтка на нижнем вегетативном полюсе)
Резко телолецетальные (с большим количествам желтка, занимает всю яйцеклетку, кроме верхнего полюса. Желтка много, сконцентрирован на вегетативном полюсе. Выделяют 2 группы: умеренно телолецитальные (молюски, земноводные) и резколецитальные (рептили и птицы). На анамальном полюсе сосредоточены цитоплазма и ядро.
Центролецетальные(желтка немного, но плотно в центре). Желтка немного, расположен в центре. Характерно для членистоногих
Бластомеры- клетки, образующиеся в результате делений дробления яйца у многоклеточных животных. Характерная особенность Б.— отсутствие роста в период между делениями, вследствие чего при очередном делении объём каждого Б. уменьшается вдвое. При голобластич. дроблении в телолецитальных яйцах Б. различаются по размерам: крупные Б.— макромеры, средние — мезомеры, мелкие — микромеры. Во время синхронных делений дробления Б., как правило, однородны по форме, структура их цитоплазмы очень проста. Затем поверхностные Б. уплощаются, и яйцо переходит к заключит, фазе дробления — бластуляции.
Строение бластулы. Если образуется сплошной шар без полости внутри, то такой зародыш называют морулой. Образование бластулы или морулы зависит от свойств цитоплазмы. Бластула образуется при достаточной вязкости цитоплазмы, морула – при слабой вязкости. При достаточной вязкости цитоплазмы бластомеры сохраняют округлую форму и только в местах соприкосновения слегка сплющиваются. Вследствие этого между ними появляется щель, которая по мере дробления увеличивается, заполняется жидкостью и превращается в бластоцель. При слабой вязкости цитоплазмы бластомеры не округляются и располагаются тесно друг возле друга, щели нет и полость не образуется. Бластулы различны по своему строению и зависят от типа дробления.
От момента образования зиготы и до выхода зародыша из яйцевых оболочек длится эмбриональный период развития.
Дробление зиготы
После того, как произошло оплодотворение - слияние сперматозоида и яйцеклетки, образовавшаяся зигота начинает интенсивно делиться. Ее множественные митотические деления называют дроблением.
Важная особенность дробления в том, что не происходит увеличение в размере зародыша: клетки дробятся (делятся) настолько быстро, что не успевают накопить цитоплазматическую массу. Дробление зиготы человека является полным неравномерным асинхронным.
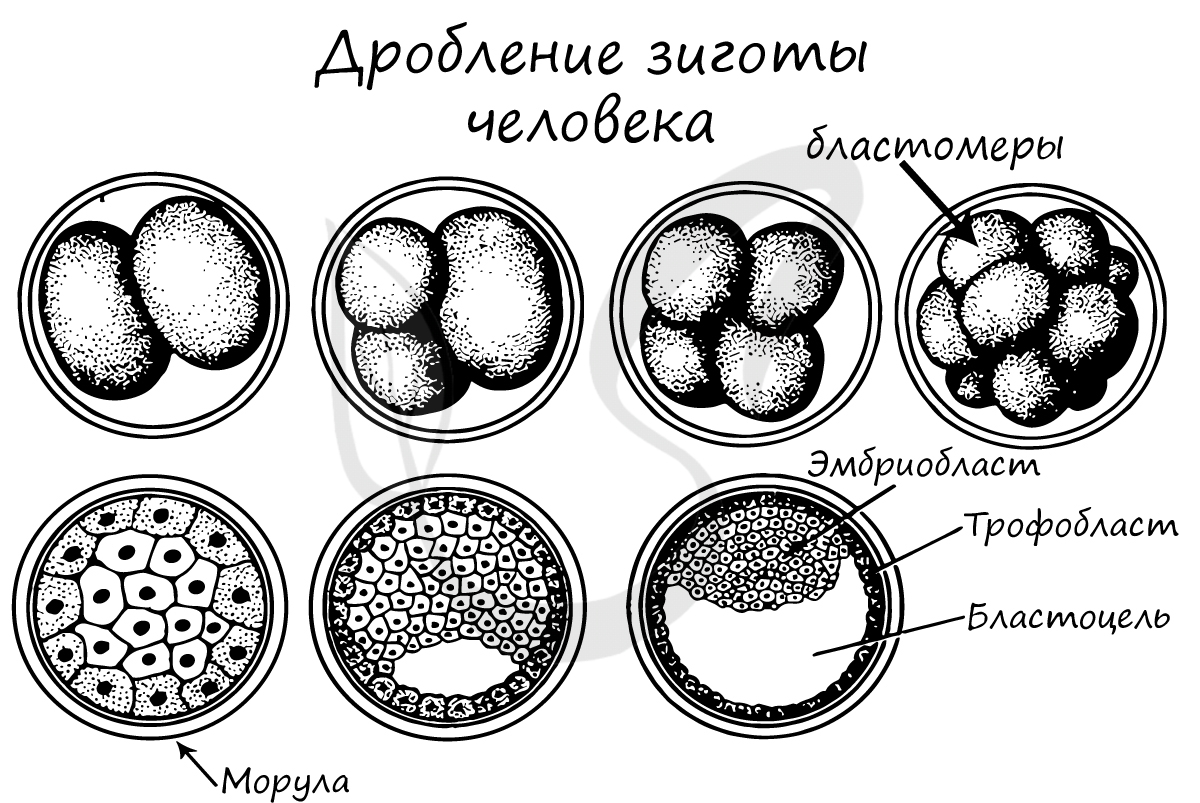
В результате дробления образуется морула. Морула (лат. morum - ягода тутового дерева) - клетка на стадии этапа дробления, когда зародыш представляет собой компактную совокупность клеток (без полости внутри).
Бластуляция
Бластуляция - заключительный период дробления, в который зародыш называется бластулой.
После очередных этапов многократного деления образуется однослойный зародыш с полостью внутри - бластула (греч. blastos — зачаток).
Стенки бластулы состоят из бластомеров, которые окружают центральную полость - бластоцель (греч. koilos — полый). Соединяясь друг с другом, бластомеры образуют бластодерму из одного слоя клеток.

Гаструляция (греч. gaster — желудок, чрево)
Гаструляцией называют стадию эмбрионального развития, в ходе которой клетки, возникшие в результате дробления зиготы, формируют три зародышевых листка: эктодерму, мезодерму и энтодерму.
Стенка бластулы начинается впячиваться внутрь - происходит инвагинация стенки. По итогу такого впячивания зародыш становится двухслойным. Двухслойный зародыш называется - гаструла. Полость гаструлы называется гастроцель (полость первичной кишки), а отверстие, соединяющее гастроцель и внешнюю среду - первичный рот (бластопор).

У первичноротых животных на месте первичного рта (бластопора) образуется ротовое отверстие. К первичноротым относятся: кишечнополостные, плоские, круглые и кольчатые черви, моллюски, членистоногие.
У вторичноротых на месте бластопора формируется анальное отверстие, а ротовое отверстие образуется на противоположном полюсе. К вторичноротым относят хордовых и иглокожих (морских звезд, морских ежей).

При впячивании части бластулы (инвагинации) клетки бластодермы мигрируют внутрь и становятся энтодермой (греч. entós — внутренний). Оставшаяся часть бластодермы снаружи называется эктодермой (греч. ἔκτος - наружный).
Между энто- и эктодермой из группы клеток формируется третий зародышевый листок - мезодерма (греч. μέσος — средний).

Нейрула
Эта стадия следует за гаструлой. Ранняя нейрула представляет собой трехслойный зародыш, состоящий из энто-, экто- и мезодермы. На этапе нейрулы происходит закладка отдельных органов.
Важно отметить, что на стадии нейрулы происходит процесс нейруляции - закладывание нервной трубки. Нервная пластинка, образовавшаяся на ранних этапах, прогибается внутрь, при этом ее края сближаются и, замыкаясь, формируют нервную трубку.

Итак, как уже было сказано, на стадии нейрулы закладываются отдельные органы. Эктодерма образует покровный эпителий и нервную пластинку, мезодерма (из которой в дальнейшем появятся все соединительные ткани), энтодерма - окружает полость первичной кишки (гастроцель), образуя кишечник. От энтодермы отшнуровывается хорда.
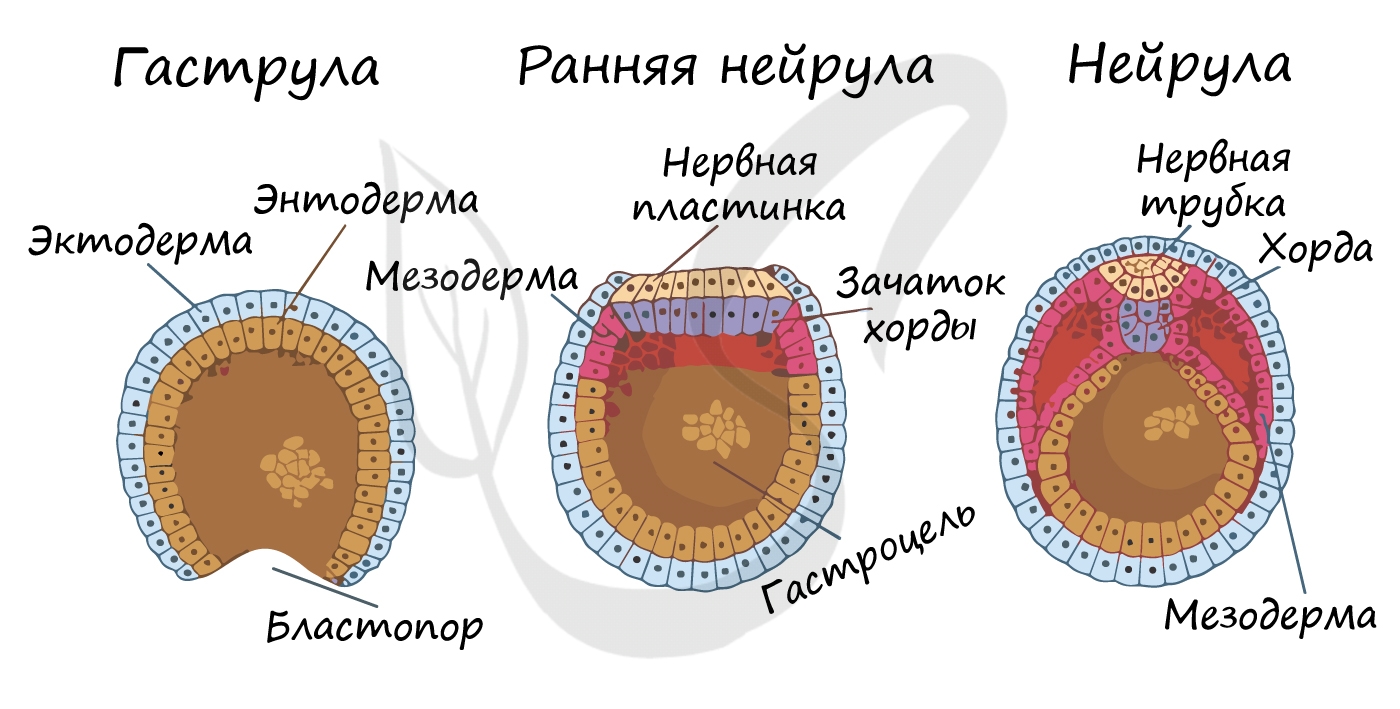
Все три зародышевых листка требуют нашего особого внимания, а также понимания того, какие органы и структуры из них образуются.
Эктодерма (греч. ἔκτος - наружный) - наружный зародышевый листок, образует головной и спинной мозг, органы чувств, периферические нервы, эпителий кожи, эмаль зубов, эпителий ротовой полости, эпителий промежуточного и анального отделов прямой кишки, гипофиз, гипоталамус.
Мезодерма (греч. μέσος — средний) - средний зародышевый листок, образует соединительные ткани: кровеносную и лимфатическую системы, костную и хрящевую ткань, мышечные ткани, дентин и цемент зубов, а также выделительную (почки) и половую системы (семенники, яичники).
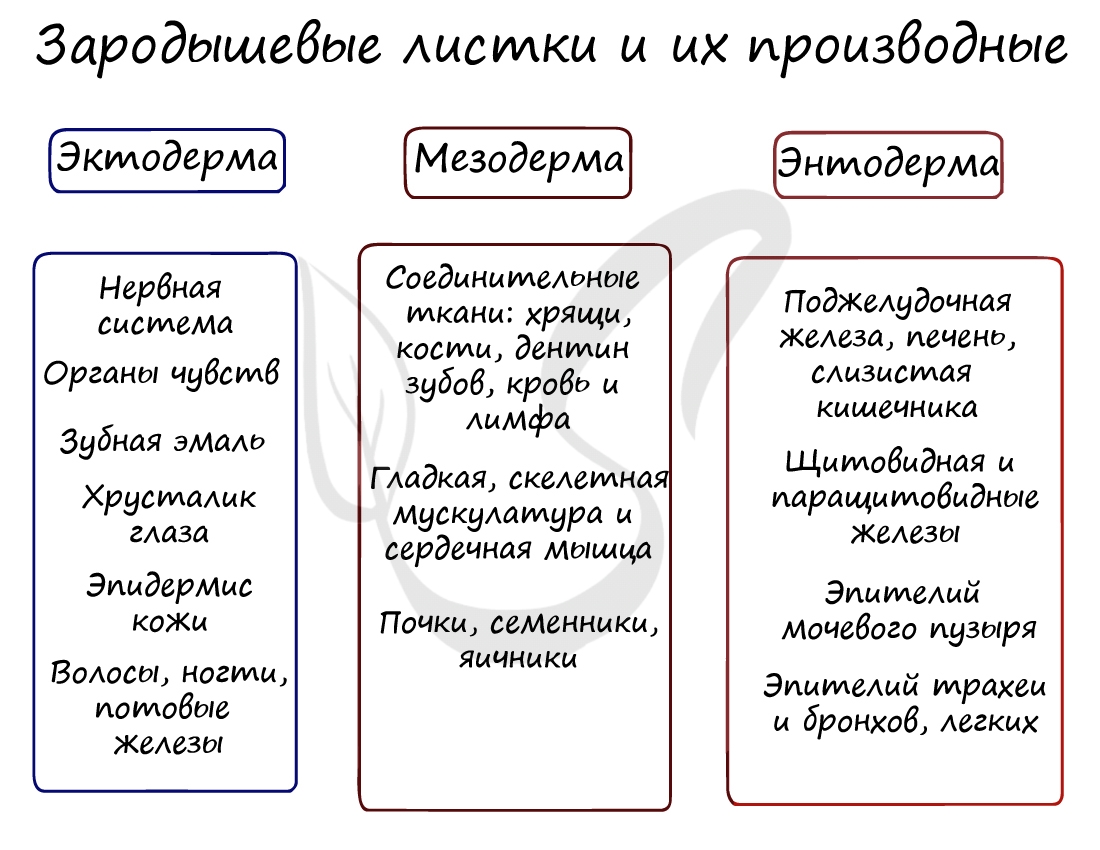
Из зародышевых листков образуются ткани, органы и системы органов. Такой процесс называется органогенезом. В период закладки органов важное значение имеет воздержание матери от вредных привычек (алкоголь, курение), которые могут нарушить процесс дифференцировки клеток и привести к тяжелейшим аномалиям, уродствам плода.
Некоторые лекарства также могут оказывать на плод тератогенный эффект (греч. τέρας — чудовище, урод), приводя к развитию уродств. Периоды закладки органов и система органов вследствие их большой важности носят название критических периодов эмбриогенеза.
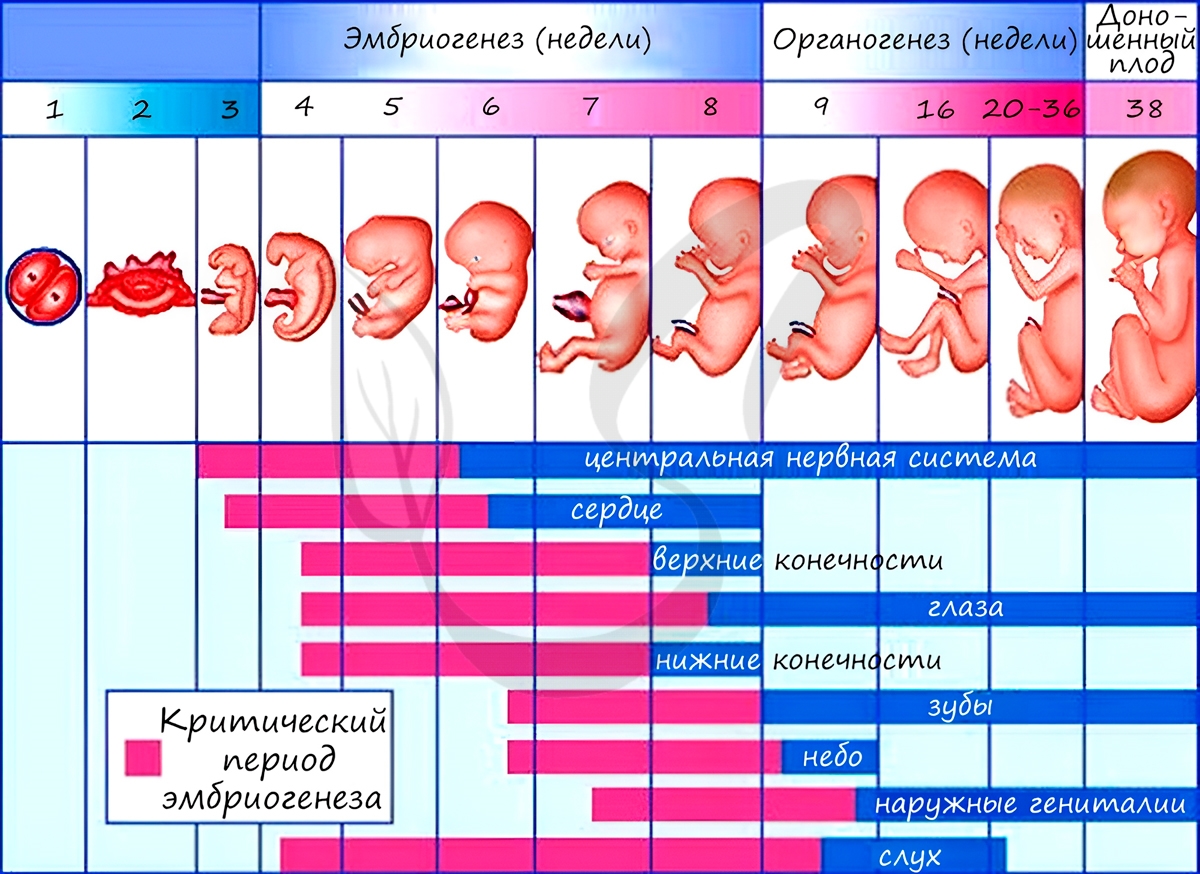
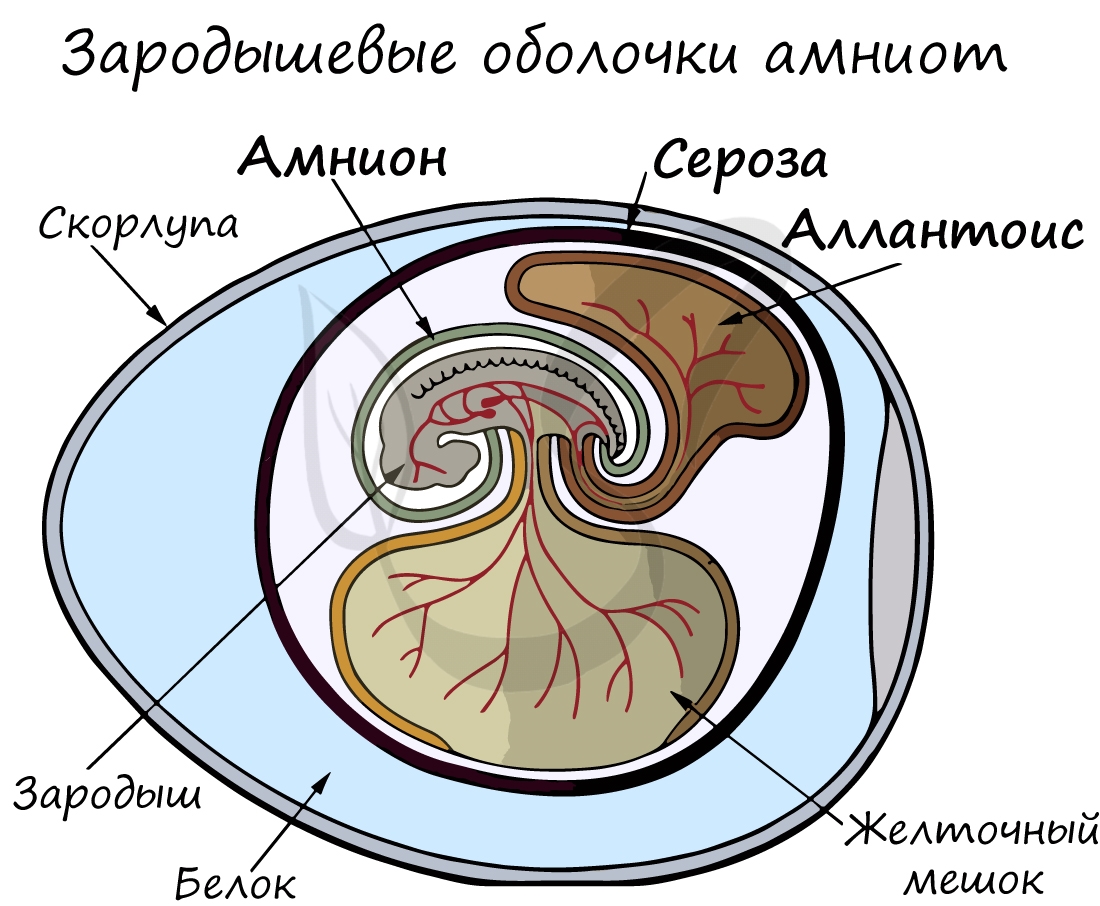
Анамнии и амниоты
Анамнии, или низшие позвоночные - группа животных, не имеющая зародышевых оболочек (зародышевого органа - аллантоиса и амниона). Анамнии проводят большую часть жизни в воде, без которой невозможно их размножение.
К анамниям относятся рыбы, земноводные.

Амниоты - группа высших позвоночных, характеризующаяся наличием зародышевых оболочек. К амниотам относятся пресмыкающиеся, птицы и млекопитающие.
Зародышевый орган, аллантоис, является органом дыхания и выделения.
За счет особых оболочек, развивающихся в ходе эмбрионального развития, амниона и серозы, у амниот формируется амниотическая полость. В ней находится зародыш, окруженный околоплодными водами. Благодаря такому гениальному устройству, амниотам для размножения и развития более не нужно постоянное нахождение в водоеме, они "обрели независимость" от него.
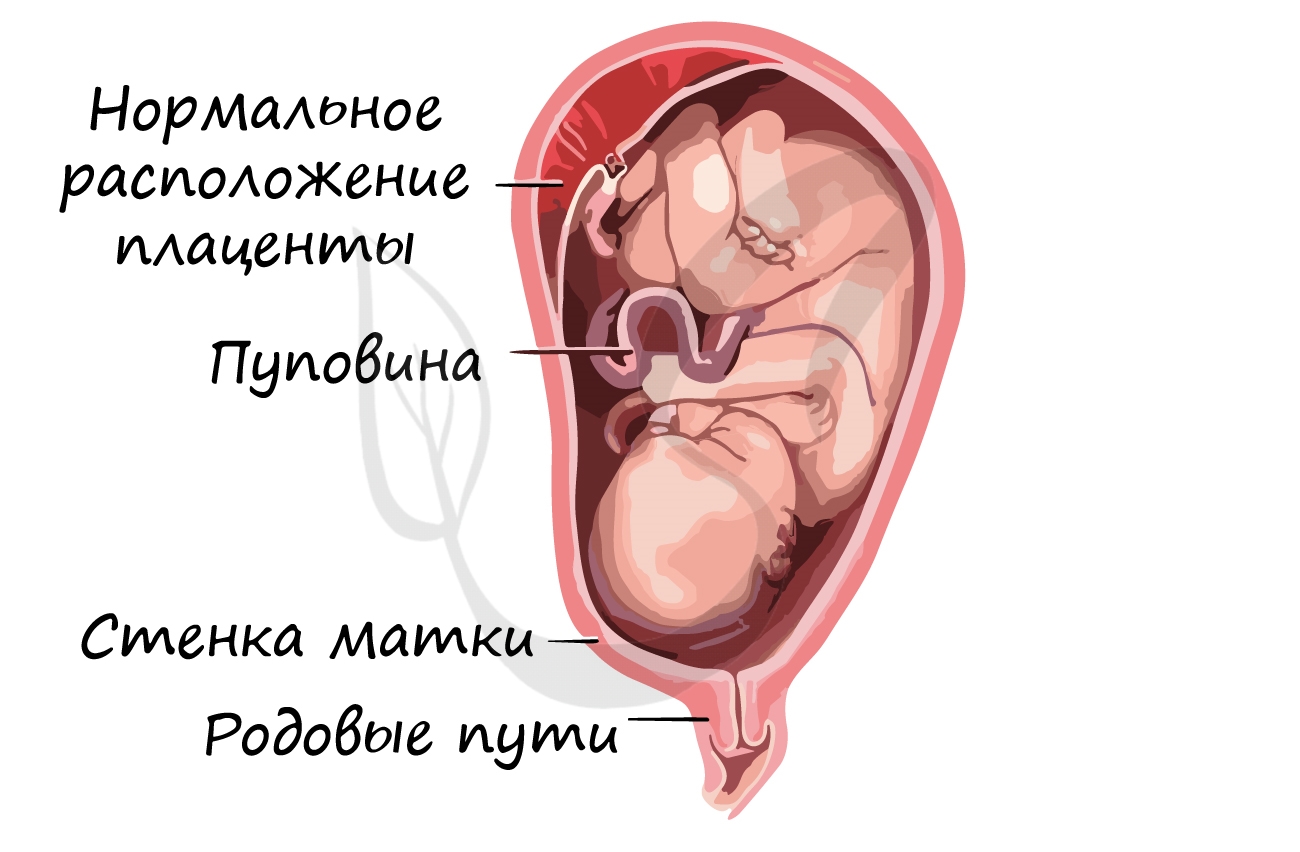
Развитие плода происходит в мышечном органе - матке, которая, сокращаясь во время родов, стимулирует изгнание плода через родовые пути. Питание осуществляется через плаценту - "детское место" - орган, который с одной стороны омывается кровью матери, а с другой - кровью плода. Через плаценту происходит транспорт питательных веществ и газообмен.
Соединяет плаценту и плод особый орган - пуповина, внутри которой проходят артерии, вены.
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Эмбриональное (зародышевое) развитие охватывает процессы от первого деления зиготы до выхода из яйца или рождения и у большинства животных включает три основных этапа: дробление, гаструляцию и органогенез.
При дроблении дочерние клетки ( бластомеры ) не расходятся и не увеличиваются в размерах. С каждым следующим делением их размеры уменьшаются.

Яйцеклетки с небольшим запасом питательных веществ делятся полностью, т. е. происходит полное дробление. Если яйцеклетка содержит большое количество желтка, то наблюдается частичное дробление — делится только диск цитоплазмы с ядром, а сам желток остаётся без изменений (например, у птиц).
Бластула — это шарообразный зародыш, стенка которого ( бластодерма ) образована одним слоем клеток, а внутри — полость ( бластоцель ).
После дробления начинается гаструляция — часть клеток бластодермы перемещается внутрь зародыша. В результате этих перемещений образуется гаструла.
Гаструла — двухслойный зародыш, состоящий из двух зародышевых листков: наружного ( эктодермы ) и внутреннего ( энтодермы ).
У ланцетника образование гаструлы происходит в результате впячивания части бластодермы внутрь бластоцеля.
Внутренняя полость гаструлы называется первичной кишкой . Её связывает с внешней средой отверстие ( бластопор ), которое становится первичным ртом .

На стадии гаструлы (двух зародышевых листков) прекращается развитие двухслойных животных — губок и кишечнополостных.
У всех остальных животных развитие продолжается, и образуется третий зародышевый листок — мезодерма . Она формируется из энтодермы и всегда расположена между экто- и энтодермой в первичной полости тела.
Дальнейшая специализация клеток зародышевых листков обеспечивает формирование тканей и органов, т. е. гисто- и органогенез .
Из энтодермы образуется хорда — внутренний скелет в виде гибкого тяжа, расположенный на спинной стороне. Позже вместо хорды у позвоночных развивается позвоночник, и только у некоторых животных (например, у хрящевых рыб) её остатки сохраняются в течение всей жизни.
Из эктодермы, расположенной над самой хордой, выделяется нервная пластинка . Затем края пластинки поднимаются и смыкаются. Образуется нервная трубка — зачаток центральной нервной системы. Формируется нейрула .

Нервная трубка, хорда и кишечник создают осевой комплекс органов зародыша, который определяет двустороннюю симметрию тела.

Из эктодермы у позвоночных животных образуется нервная система, органы чувств, покровный эпителий с его железами и производными структурами (волосы, перья, копыта, когти и т. п.).
Из энтодермы формируются органы пищеварительной и дыхательной системы: эпителий средней кишки, печень и поджелудочная железа, жабры, лёгкие, плавательный пузырь, а также щитовидная железа.
Из мезодермы формируются все виды мышечной и соединительной ткани (например, дерма кожи, тела позвонков), кровеносная система, органы выделения, половые железы.
Эмбриология человека – это направление науки, занимающееся изучением развития зародыша, то есть организма на ранних стадиях развития до рождения. Знания в области эмбриологии человека необходимы всем врачам, особенно работающим в направлении педиатрии и акушерства.
Знания эмбриологии оказывают помощь при диагностике нарушений в системе мать-плод, выявлении болезней детей после рождения, а также выявлении причин уродств.
На сегодняшний день знания в сфере эмбриологии применяют для выявления и ликвидации причин бесплодия, разработки противозачаточных препаратов, трансплантации фетальных органов. Приобрели актуальность проблемы трансплантации зародыша в матку, экстракорпорального оплодотворения и культивирования яйцеклеток.
Эмбриология изучает несколько стадий развития зародыша:
- оплодотворение с дальнейшим образованием зиготы;
- дробление и образование бластоцисты;
- гаструляцию – процесс образования зародышевых листов и осевых органов;
- органогенез и гистогенез внезародышевых и зародышевых органов;
- системогенез.
Внутриутробное развитие делится на три основных периода:
- начальный – первая неделя;
- зародышевый – вторая-восьмая недели;
- плодный – начинается с девятой недели и завершается рождением ребенка.
В среднем внутриутробное развитие человека продолжается 280 суток.
Эмбриология: стадия оплодотворения и образования зиготы
Оплодотворение – процесс слияния мужских и женских половых клеток, в результате которого восстанавливается диплоидный набор хромосом и возникает новая клетка – оплодотворенная яйцеклетка (зигота). Для возможности оплодотворения концентрация в эякуляте сперматозоидов должна соответствовать 20-200 млн/мл, а их общее количество – 150 млн/мл.
Процесс оплодотворения состоит из трех фаз:
- дистантного взаимодействия и сближения гамет;
- контактного взаимодействия с активацией яйцеклетки;
- проникновения сперматозоида в яйцеклетку с последующей сингамией (слиянием).
Дистантное взаимодействие обеспечивает хемотаксис - совокупность специфических факторов, отвечающих за повышение вероятности встречи мужских и женских половых клеток. В этом процессе важную роль играют вырабатываемые половыми клетками химические вещества.
Сразу после эякуляции происходит процесс капацитации – сперматозоиды под воздействием секрета женских половых путей приобретают оплодотворяющую способность. На механизм капацитации большое влияние оказывают гормональные факторы (например, прогестерон), активизирующие секрецию маточных труб.
Оплодотворение происходит в маточных трубах, ему предшествует осеменение, обусловленное хемотаксисом.
При контактном взаимодействии сперматозоиды приближаются к яйцеклетке, а затем вступают в контакт с ее оболочкой.
Далее происходит процесс проникновения головки и хвоста спермия в овоплазму. На периферии овоплазмы образуется оболочка оплодотворения.
В организме женщины в течение 12 часов после сближения мужского и женского пронуклеусов образуется одноклеточный зародыш – зигота.
Эмбриология: стадия дробления и образования бластоцисты
Дробление – это последовательный процесс деления зиготы без роста бластомеров. У человека дробление полное, асинхронное и неравномерное.
После первого дробления в организме женщины образуются два бластомера. Один из бластомеров обладает более крупными размерами и темной окраской, второй – светлый и более мелкий.
Из крупного бластомера происходит образование зародыша и большинства провизорных органов: плодной части плаценты и соединительной ткани хориона, желточного мешка, амниона, аллантоиса. Из второго бластомера развивается трофобласт.
Образование бластулы
Мелкие клетки в процессе дробления делятся быстрее крупных и обрастают их снаружи. Таким образом, образуется морула – скопление клеток. Внутри нее расположены крупные клетки, названные эмбриобластом, а снаружи мелкие клетки, названные трофобластом.
В ходе деления клеток морула увеличивается в размерах, клетками зародыша начинает секретироваться жидкость и накапливаться под трофобластом.
В дальнейшем объем жидкости увеличивается, образуется полость внутри зародыша, наполненная такой жидкостью, эмбриобласт оттесняется к периферии и прилипает к трофобласту. Образуется бластоциста.
Трофобласт образует выросты – ворсинки, вследствие чего поверхность бластулы неровная. Трофобласт – это первый провизорный орган, образующийся у зародыша. В дальнейшем трофобласт войдет в состав плаценты. Посредством трофобласта происходит имплантация зародыша в слизистую оболочку матки.
Эмбриология: стадия гаструляции
В результате перемещения клеток после образования бластулы образуется гаструла – двуслойный зародыш. Процесс образования гаструлы назван гаструляцией.
В процессе гаструляции происходит интенсивное перемещение клеток – будущие зачатки тканей перемещаются в соответствии с планом структурной организации будущего полноценного организма.
На стадии гаструляции зародыш состоит из зародышевых листков - разделенных пластов клеток. Наружный слой – эктодерма, внутренний – энтодерма. У позвоночных животных образуется третий слой (средний) – мезодерма.
Из эктодермы развиваются:
- эпителий кожи;
- нервная система;
- эмаль зубов;
- органы чувств.
Из энтодермы развиваются:
- эпителий легких;
- пищеварительные железы;
- эпителий средней кишки.
Из мезодермы развиваются:
- кровеносная система;
- соединительная и мышечная ткани;
- половые железы;
- почки и др.
Выделяют несколько способов гаструляции:
- инвагинация – осуществляется путем втягивания в бластоцель стенки бластулы;
- деляминация – в эпителиальный пласт эктодермы преобразуются клетки, располагающиеся снаружи, а оставшиеся формируют энтодерму. Деляминация характерна для кишечнополостных;
- эпиболия – обрастание клетками при неполном дроблении внутренней массы желтка или обрастание клеток другими быстро делящимися клетками;
- иммиграция – миграция внутрь бластоцеля части клеток стенки бластулы;
- инволюция – вворачивание наружного пласта клеток, увеличивающего в размерах, внутрь зародыша.
Эмбриология: стадия гистогенеза и органогенеза внезародышевых и зародышевых органов
Органогенез – совокупность процессов, приводящих к формированию зачатков органов и их последующей дифференциации в процессе эмбрионального развития.
В органогенезе выделяют:
- нейруляцию – процесс образования нейрулы. В нейруле закладывается мезодерма, состоящая, в свою очередь, из зародышевых листков и осевого комплекса органов – хорды, нервной трубки и кишки. Клетки комплекса органов влияют друг на друга. Такое влияние носит название эмбриональной индукции.
- гистогенез – ряд процессов, обеспечивающих образование и восстановление тканей в ходе онтогенеза.
На сегодняшний день эмбриология стала одним из важнейших направлений науки. В медицине ее применение не ограничивается областью гистологии и анатомии. Эмбриология имеет важное значение в развитии профилактической медицины, направленной на разработку и тестирование новых медицинских препаратов, борьбу с наследственными заболеваниями. Эмбриология имеет большие перспективы, связанные с развитием генетики и ряда других наук.
Также эмбриология тесно связана с ЭКО, так как эмбриологический период является одним из важнейших этапов программы экстракорпорального оплодотворения.
Клиническая эмбриология изучает причины нарушений эмбрионального развития, механизмы развития уродств, а также способы влияния на эмбриогенез.
Разработки в области ЭКО стали возможными благодаря использованию высокотехнологической медицины и развитию клинической эмбриологии. Исход экстракорпорального оплодотворения в большой степени зависит от знаний и опыта специалиста-эмбриолога.
Эмбриональное развитие человека. Оплодотворение - фертилизация.
Онтогенез, или индивидуальное развитие, включает пренатальный (внутриутробный) период, который длится примерно 280 суток, или 10 лунных месяцев, и постнатальный (внеутробный) период, продолжительность которого у разных людей варьирует и во многом определяется как внутренними, так и внешними по отношению к человеку факторами.
Изучение пренатального развития человека (эмбриогенеза) встречается с рядом трудностей, связанных не только с получением необходимого для исследований материала, но этическими и религиозными нормами, существующими в общественном сознании. Ранние зародыши человека — "редкая находка". Только в 1944 г. впервые был исследован 7,5-суточный зародыш человека, а в 1946 г. — 2-5-суточные зародыши. Наиболее полная коллекция человеческих эмбрионов находится в институте Карнеги (Балтимор, США). Описания ранних эмбрионов человека даны Гертигом, Рокком и Стритером. В отечественной эмбриологии ранние стадии развития зародыша человека изучены и описаны А.Г. Кнорре (зародыш "ВМА-1") и Б.П. Хватовым (зародыш "Крым"). Развитие технологии искусственного оплодотворения позволило в деталях исследовать механизмы оплодотворения и дробления зиготы у человека.
Оплодотворение (фертилизация)
У человека оплодотворение внутреннее. По клиническим наблюдениям, зачатие чаще всего происходит у женщин вплоть до 2-й недели после менструации, хотя другие авторы указывают в качестве наиболее подходящих для зачатия сроков —11-17-е сутки менструального цикла.
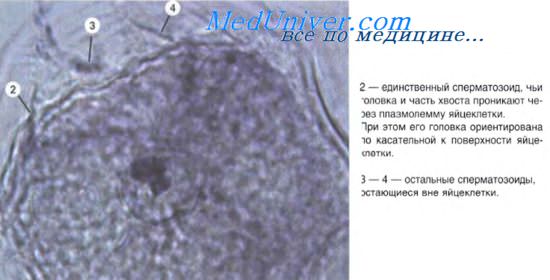
В результате гаметогенеза у человека образуется генетически однородная популяция овоцитов (яйцеклеток), содержащих 22 соматические и одну половую Х- хромосомы; и два рода спермиев с различными генетическими характеристиками (22+Х и 22+Y). Последние образуются в равном количестве, поэтому яйцеклетка имеет статистически равные возможности на встречу как с Х-, так и с Y-спермием и, соответственно, ожидается рождение мальчиков и девочек в равном соотношении. Однако физиологические условия оплодотворения вносят поправку в эти результаты (100: 106 в пользу рождения мальчиков).
Процесс направленного перемещения спермиев по органам женского полового тракта из влагалища в маточную трубу продолжается около 10 часов и является, по сути, преодолением клетками с ограниченными метаболическими потенциями огромного расстояния. Благодаря тому, что в эякуляте содержится в среднем около 200-300 млн сперматозоидов, существует высокая вероятность того, что небольшая часть спермиев (около 1% от первоначального числа) сохранит жизнеспособность, достигнет маточной трубы и будет участвовать в оплодотворении. Скорость самостоятельного передвижения спермиев очень невелика — около 2-4 мм/мин.
Женская половая клетка при овуляции попадает в маточную трубу благодаря набуханию фимбрий и их тесному контакту с поверхностью яичника.
При взаимодействии сперматозоидов с органами женского репродуктивного тракта поисходит их капацитация — приобретение оплодотворяющей способности. При капацитации под влиянием секреторных продуктов женского полового тракта с поверхности сперматозоидов удаляются вещества, которые блокируют рецепторно-трансдукторную систему сперматозоида, взаимодействующую с поверхностью женской половой клетки. Собственно процесс оплодотворения условно разделяют на фазы — дистантного и контактного взаимодействия и завершается оплодотворение активацией метаболизма зиготы.
В фазе дистантного взаимодействия происходит встреча половых клеток (гамет) в половых путях женщины. Важными механизмами дистантного взаимодействия являются положительные хемо- и реотаксис, а также электростатическое взаимодействие гамет (на близком расстоянии).
В фазе контактного взаимодействия спермин разрушает оболочки овоцита — лучистый венец, прозрачную зону и плазмолемму. В процессе оплодотворения не должно произойти полиспермии — проникновения в женскую половую клетку нескольких сперматозоидов. Полагают, что первый этап взаимодействия спермиев с женской половой клеткой — это механическое удаление части клеток лучистого венца, которое осуществляют биения жгутиков спермиев. Дальнейшие события контактного взаимодействия связаны со взаимодействием рецепторов двух клеток, акросомной реакцией сперматозоида и кортикальной реакцией женской половой клетки. При контакте с женской половой клеткой, под действием активирующих субстанций (одна из которых — фертилизин), инициируется активное поступление катионов кальция в головку спермия. В результате происходят очаговые слияния плазмолеммы овоцита и акросомальной мембраны сперматозоида и их разрушение с появлением микроперфораций.
Через образовавшиеся микроотверстия выделяются ферменты-лизины спермия (гиалуронидаза, трипсиноподобный фермент и др.), которые разобщают контакты между клетками лучистого венца, а также между ними и плазмолеммой овоцита. Диссоциация лучистого венца прогрессирует и наконец обнажается небольшой участок более глубоко расположенной прозрачной зоны. Выделяемый акросомой спермия акрозин разрушает в этом участке гликозаминогликаны прозрачной зоны и образует "окно", через которое спермий может проникнуть к женской половой клетке. Пенетрация прозрачной зоны продолжается около 20 минут. После разрушения участка прозрачной зоны спермий попадает в заполненное жидкой средой перивителлиновое пространство между прозрачной зоной и плазмолеммой овоцита. В месте соприкосновения головки спермия с плазмолеммой овоцита цитоплазма женской половой клетки формирует выступ — бугорок оплодотворения (в этом участке овоцита активируется полимеризация актина) и здесь же происходит слияние внешних мембран женской и мужской гамет.
Слившиеся участки мембран затем разрушаются и через образовавшееся отверстие спермий проникает в женскую половую клетку. Его плазмолемма при этом "сползает" и закрывает дефект, образовавшийся в плазмолемме овоцита. Из цитоплазматических структур спермия кроме ядра в овоцит попадают проксимальная центриоль и шейка (хвост остается снаружи и отпадает). Благодаря тому, что участок мембраны, привнесенный в плазмолемму овоцита спермием, хорошо проницаем для катионов натрия, последние начинают активно поступать внутрь женской половой клетки и изменяют ее мембранный потенциал. В течение очень короткого времени (около 1/10 сек) мембранный потенциал овоцита резко падает, и женская половая клетка становится невосприимчивой к контактам с другими спермиями. Затем наступает кортикальная реакция овоцита. Это происходит в результате поступления катионов кальция в женскую половую клетку, что вызывает слияние мембран кортикальных гранул с плазмолеммой овоцита и экзоцитоз их ферментов в перивителлиновое пространство. При этом прозрачная зона уплотняется, утолщается, теряет рецепторные белки к сперматозоидам. Так возникает оболочка оплодотворения, препятствующая проникновению других спермиев в овоцит.
В момент встречи со спермием овоцит находится в блоке метафазы второго мейо-тического деления. После проникновения спермия в овоплазму женская половая клетка завершает второе деление созревания. При этом выделяется полярное тельце с лишними хромосомами. Пока овоцит завершает мейоз, пронуклеус спермия округляется и принимает вид интерфазного. В нем происходит синтез ДНК, и пронуклеус приобретает набор двойных (реплицированных) хромосом. Ядро женской половой клетки по завершении второго деления мейоза претерпевает точно такие же изменения. Затем оба пронуклеуса переходят в профазу митоза. Центриоль, привнесенная спермием, делится, формируя две центросомы. Последние прикрепляются к веретену деления, образующемуся между пронуклеусами и, таким образом, хромосомы мужского и женского пронуклеусов оказываются расположенными в экваториальной плоскости — происходит метафаза митоза. Далее следуют ана- и телофаза — зигота завершает первое деление дробления, в результате которого образуются первые две дочерние клетки — бластомеры — каждая с диплоидным набором хромосом.
Читайте также:
- Фз об аккредитации в национальной системе аккредитации кратко
- Этапы развития документа кратко
- Оформление фотовыставки ко дню матери в детском саду младшая группа
- Выступление учителя английского языка на родительском собрании в начальной школе
- Проектирование воспитательной работы школы на основе системы классных часов