
Нарушители основного правила молекулярной биологии кратко
Обновлено: 03.07.2024
МОЛЕКУЛЯ́РНАЯ БИОЛО́ГИЯ, изучает молекулярные основы жизнедеятельности организмов. Гл. направления исследований связаны с изучением механизмов хранения, воспроизведения и передачи генетич. информации. Исходя из этого, осн. объектами исследования М. б. являются нуклеиновые кислоты – дезоксирибонуклеиновые (ДНК) и рибонуклеиновые (РНК) – и белки, а также их макромолекулярные комплексы – хромосомы, рибосомы, мультиферментные системы, обеспечивающие биосинтез нуклеиновых кислот и белков. М. б. граничит, а по объектам и методам исследования частично совпадает с молекулярной генетикой, вирусологией, систематикой организмов, биохимией, клеточной биологией, биофизикой, биоорганич. химией, кристаллографией, компьютерным моделированием, биоинформатикой и др. М. б. не только зависит от прогресса этих наук, но и оказывает существенное влияние на их развитие, ставя перед ними весьма нетривиальные задачи.
В любой науке есть свод законов, которые ученые открыли, но до конца объяснить их причину не смогли. Так центральной догмой молекулярной биологии является обобщение:
Действительно, генетическая информация закодирована в ДНК. С помощью другой нуклеиновой кислоты - РНК - эта информация копируется частями и раскодируется в белковые молекулы. Эти процессы называются транскрипцией и трансляцией .
Однако в природе не наблюдается обратного явления. Ни в одной клетке, ни в одном организме нет ферментных систем, способных по белковой молекуле воссоздать участок ДНК. Хотя по логике вещей, если ты кодируешь в одну сторону, то можешь раскодировать в обратную.
Другими словами, если представить гипотетический организм, в котором по его ДНК были синтезированы все белки, а потом ДНК исчезло, то мы не сможем восстановить исходное ДНК, опираясь на последовательность аминокислот в имеющихся белках.
И дело здесь не только в отсутствии химической возможности. Если бы в процессе эволюции возникла потребность в таком процессе, то соответствующие ферменты появились бы.
По всей видимости важную роль сыграли особенности самого генетического кода ( статья о единстве генетического кода для всего живого ). В нем почти каждой аминокислоте, входящей в состав белка, соответствует далеко не один вариант триплета в нуклеиновой кислоте.
В результате по участку ДНК мы можем однозначно синтезировать конкретный белок. А вот по белковой молекуле не можем однозначно восстановить участок ДНК. Потому что каждую аминокислоту белка можно раскодировать не одним способом, а несколькими. А каким точно, мы не знаем, если до этого не видели исходной ДНК.
А в чем собственно проблема? Пусть по белку нельзя восстановить исходную ДНК, но даже немного другая ДНК будет кодировать все те же белки. На этот вопрос ответить сложнее. Возможно оказалась важна слаженность генома, его тонкая эволюционная настройка. В результате порядок нуклеотидов стал важен не только для синтеза конкретного белка, но и для работы ферментов, выполняющих этот синтез.

Обзор
Молекулярный биолог Пробирочка
Автор
Редакторы



Спонсором приза зрительских симпатий выступила компания BioVitrum.
1. Введение. Сущность молекулярной биологии
Молекулярная биология изучает основы жизнедеятельности организмов на уровне макромолекул. Целью молекулярной биологии является установление роли и механизмов функционирования этих макромолекул на основе знаний об их структурах и свойствах.
Исторически молекулярная биология сформировалась в ходе развития направлений биохимии, изучающих нуклеиновые кислоты и белки. В то время как биохимия исследует обмен веществ, химический состав живых клеток, организмов и осуществляемые в них химические процессы, молекулярная биология главное внимание сосредоточивает на изучении механизмов передачи, воспроизведения и хранения генетической информации.
А объектом изучения молекулярной биологии являются сами нуклеиновые кислоты — дезоксирибонуклеиновые (ДНК), рибонуклеиновые (РНК) — и белки, а также их макромолекулярные комплексы — хромосомы, рибосомы, мультиферментные системы, обеспечивающие биосинтез белков и нуклеиновых кислот. Молекулярная биология также граничит по объектам исследования и частично совпадает с молекулярной генетикой, вирусологией, биохимией и рядом других смежных биологических наук.
2. Исторический экскурс по этапам развития молекулярной биологии
Как отдельное направление биохимии, молекулярная биология начала развиваться в 30-х годах прошлого века. Еще тогда возникла необходимость понимания феномена жизни на молекулярном уровне для исследований процессов передачи и хранения генетической информации. Как раз в то время установилась задача молекулярной биологии в изучении свойств, структуры и взаимодействия белков и нуклеиновых кислот.
В 1944 году американский биолог Освальд Эвери с коллегами (Колином Маклеодом и Маклином Маккарти) доказал, что веществом, вызывающим трансформацию бактерий, является ДНК, а не белки. Эксперимент послужил доказательством роли ДНК в передаче наследственной информации, перечеркнув устаревшие знания о белковой природе генов.
В начале 50-х годов Фредерик Сенгер показал, что белковая цепь — уникальная последовательность аминокислотных остатков. В 1951 и 1952 годах ученый определил полную последовательность двух полипептидных цепей — бычьего инсулина В (30 аминокислотных остатков) и А (21 аминокислотный остаток) соответственно.
Примерно в то же время, в 1951–1953 гг., Эрвин Чаргафф сформулировал правила о соотношении азотистых оснований в ДНК. Согласно правилу, вне зависимости от видовых различий живых организмов в их ДНК количество аденина (A) равно количеству тимина (T), а количество гуанина (G) равно количеству цитозина (C).
В 1953 году доказана генетическая роль ДНК. Джеймс Уотсон и Фрэнсис Крик на основе рентгенограммы ДНК, полученной Розалинд Франклин и Морисом Уилкинсом, установили пространственную структуру ДНК и выдвинули подтвердившееся позднее предположение о механизме ее репликации (удвоении), лежащем в основе наследственности.
1958 год — формирование центральной догмы молекулярной биологии Фрэнсисом Криком: перенос генетической информации идет в направлении ДНК → РНК → белок.
Суть догмы состоит в том, что в клетках имеется определенный направленный поток информации от ДНК, которая, в свою очередь, представляет собой исходный генетический текст, состоящий из четырех букв: A, T, G и C. Он записан в двойной спирали ДНК в виде последовательностей этих букв — нуклеотидов.
Этот текст транскрибируется. А сам процесс называется транскрипцией. В ходе данного процесса происходит синтез РНК, которая является идентичной генетическому тексту, но с отличием: в РНК вместо T стоит U (урацил).
Данная РНК называется информационной РНК (иРНК), или матричной (мРНК). Трансляция иРНК осуществляется при помощи генетического кода в виде триплетных последовательностей нуклеотидов. В ходе этого процесса происходит перевод текста нуклеиновых кислот ДНК и РНК из четырехбуквенного текста в двадцатибуквенный текст аминокислот.
Природных аминокислот существует всего двадцать, а букв в тексте нуклеиновых кислот четыре. Из-за этого происходит перевод из четырехбуквенного алфавита в двадцатибуквенный посредством генетического кода, в котором каждым трем нуклеотидам соответствует какая-либо аминокислота. Так можно сделать из четырех букв целые 64 трехбуквенные комбинации, притом что аминокислот 20. Из этого следует, что генетический код обязательно должен иметь свойство вырожденности. Однако в то время генетический код не был известен, к тому же его даже не начали расшифровывать, но Крик уже сформулировал свою центральную догму.
Тем не менее была уверенность в том, что код должен существовать. К тому времени было доказано, что этот код обладает триплетностью. Это означает, что конкретно три буквы в нуклеиновых кислотах (кодóны) отвечают какой-либо аминокислоте. Этих кодонов всего 64, они кодируют 20 аминокислот. Это означает, что каждой аминокислоте отвечает сразу несколько кодонов.
Таким образом, можно сделать вывод, что центральная догма является постулатом, который гласит о том, что в клетке происходит направленный поток информации: ДНК → РНК → белок. Крик сделал акцент на главном содержании центральной догмы: обратного потока информации происходить не может, белок не способен изменять генетическую информацию.
В этом и заключается основной смысл центральной догмы: белок не в состоянии изменять и преобразовывать информацию в ДНК (или РНК), поток всегда идет лишь в одну сторону.
Спустя время после этого был открыт новый фермент, который не был известен во времена формулировки центральной догмы, — обратная транскриптаза, которая синтезирует ДНК по РНК. Фермент был открыт в вирусах, у которых генетическая информация закодирована в РНК, а не в ДНК. Такие вирусы называют ретровирусами. Они имеют вирусную капсулу с заключенными в нее РНК и специальным ферментом. Фермент и есть обратная транскриптаза, которая синтезирует ДНК по матрице этой вирусной РНК, а эта ДНК потом уже служит генетическим материалом для дальнейшего развития вируса в клетке.
Конечно, данное открытие вызвало большой шок и множество споров среди молекулярных биологов, поскольку считалось, что, исходя из центральной догмы, этого быть не может. Однако Крик сразу объяснил, что он никогда не говорил, что это невозможно. Он говорил лишь то, что никогда не может происходить поток информации от белка к нуклеиновым кислотам, а уже внутри нуклеиновых кислот любого рода процессы вполне возможны: синтез ДНК на ДНК, ДНК на РНК, РНК на ДНК и РНК на РНК.
После формулирования центральной догмы по-прежнему оставался ряд вопросов: как алфавит из четырех нуклеотидов, составляющих ДНК (или РНК), кодирует 20-буквенный алфавит аминокислот, из которых состоят белки? В чем состоит сущность генетического кода?
Первые идеи о существовании генетического кода сформулировали Александр Даунс (1952 г.) и Георгий Гамов (1954 г.). Ученые показали, что последовательность нуклеотидов должна включать в себя не менее трех звеньев. Позднее было доказано, что такая последовательность состоит из трех нуклеотидов, названных кодоном (триплетом). Тем не менее вопрос о том, какие нуклеотиды ответственны за включение какой аминокислоты в белковую молекулу, оставался открытым до 1961 года.
А в 1961 году Маршалл Ниренберг вместе с Генрих Маттеи использовали систему для трансляции in vitro. В роли матрицы взяли олигонуклеотид. В его состав входили только остатки урацила, а пептид, синтезированный с него, включал только аминокислоту фенилаланин. Таким образом впервые было установлено значение кодона: кодон UUU кодирует фенилаланин. Поле них Хар Корана выяснил, что последовательность нуклеотидов UCUCUCUCUCUC кодирует набор аминокислот серин—лейцин—серин—лейцин. По большому счету, благодаря работам Ниренберга и Кораны, к 1965 году генетический код был полностью разгадан. Выяснилось, что каждый триплет кодирует определенную аминокислоту. А порядок кодонов определяет порядок аминокислот в белке.
Главные принципы функционирования белков и нуклеиновых кислот сформулировали к началу 70-х годов. Было зафиксировано, что синтез белков и нуклеиновых кислот осуществляется по матричному механизму. Молекула-матрица несет закодированную информацию о последовательности аминокислот или нуклеотидов. При репликации или транскрипции матрицей служит ДНК, при трансляции и обратной транскрипции — иРНК.
Так были созданы предпосылки для формирования направлений молекулярной биологии, в том числе и генной инженерии. А в 1972 году Пол Берг с коллегами разработал технологию молекулярного клонирования. Ученые получили первую рекомбинантную ДНК in vitro. Эти выдающиеся открытия легли в основу нового направления молекулярной биологии, а 1972 год с тех пор считается датой рождения генной инженерии.
3. Методы молекулярной биологии
Колоссальные успехи в изучении нуклеиновых кислот, строении ДНК и биосинтеза белка привели к созданию ряда методов, имеющих большое значение в медицине, сельском хозяйстве и науке в целом.
После изучения генетического кода и основных принципов хранения, передачи и реализации наследственной информации для дальнейшего развития молекулярной биологии стали необходимы специальные методы. Эти методы позволили бы проводить манипуляции с генами, изменять и выделять их.
Появление таких методов произошло в 1970–1980-х годах. Это дало огромный толчок развитию молекулярной биологии. В первую очередь, эти методы напрямую связаны с получением генов и их внедрением в клетки других организмов, а еще с возможностью определения последовательности нуклеотидов в генах.
3.1. Электрофорез ДНК
Электрофорез ДНК является базовым методом работы с ДНК. Электрофорез ДНК применяется вместе почти со всеми остальными методами для выделения нужных молекул и дальнейшего анализа результатов. Сам метод электрофореза в геле используется для разделения фрагментов ДНК по длине.
Предварительно или после электрофореза гель обрабатывается красителями, которые способны связаться с ДНК. Красители флуоресцируют в ультрафиолетовом свете, получается картина из полос в геле. Для определения длин фрагментов ДНК их можно сравнить с мáркерами — наборами фрагментов стандартных длин, которые наносятся на тот же гель.
Флуоресцентные белки
При исследовании эукариотических организмов в качестве генов-мáркеров сподручно использовать флуоресцентные белки. Ген первого зеленого флуоресцентного белка (green fluorescent protein, GFP) выделили из медузы Aqeuorea victoria, после чего внедрили в различные организмы. После выделяли гены флуоресцентных белков других цветов: синих, желтых, красных. Чтобы получить белки с интересующими свойствами, такие гены были модифицированы искусственно.
Вообще, важнейшими инструментами для работы с молекулой ДНК являются ферменты, осуществляющие ряд превращений ДНК в клетках: ДНК-полимеразы, ДНК-лигазы и рестриктазы (рестрикционные эндонуклеазы).
Трансгенез
Трансгенезом называется перенос генов из одного организма в другой. А такие организмы называются трансгенными.
Рекомбинантные белковые препараты как раз получают методом переноса генов в клетки микроорганизмов. В основном такими белковыми препаратами являются интерфероны, инсулин, некоторые белковые гормоны, а также белки для производства ряда вакцин.
В иных случаях применяют клеточные культуры эукариот или трансгенных животных, по большей степени, скот, который выделяет нужные белки в молоко. Таким образом получают антитела, факторы свертывания крови и другие белки. Метод трансгенеза используют для получения культурных растений, устойчивых к вредителям и гербицидам, а при помощи трансгенных микроорганизмов очищают сточные воды.
Помимо всего перечисленного, трансгенные технологии незаменимы в научных исследованиях, ведь развитие биологии происходит быстрее с применением методов модификации и переноса генов.
Рестриктазы
Распознаваемые рестриктазами последовательности являются симметричными, поэтому всякого рода разрывы могут происходить либо в середине такой последовательности, либо со сдвигом в одной или обеих нитях молекулы ДНК.
При расщеплении любой ДНК рестриктазой, последовательности на концах фрагментов будут одинаковыми. Они смогут снова соединяться, поскольку имеют комплементарные участки.
Получить единую молекулу можно, сшив данные последовательности при помощи ДНК-лигазы. За счет этого возможно объединять фрагменты двух разных ДНК и получать рекомбинантные ДНК.
3.2. ПЦР
В основе метода лежит способность ДНК-полимераз достраивать вторую нить ДНК по комплементарной нити так же, как при процессе репликации ДНК в клетке.
3.3. Секвенирование ДНК
Стремительное развитие метода секвенирования позволяет эффективно определять особенности исследуемого организма на уровне его генома. Главным преимуществом таких геномных и постгеномных технологий является увеличение возможностей исследования и изучения генетической природы заболеваний человека, для того чтобы заранее принять необходимые меры и избежать болезней.
За счет крупных исследований возможно получать необходимые данные о различных генетических характеристиках разных групп людей, тем самым развивая методы медицины. Из-за этого выявление генетической расположенности к различным заболеваниям сегодня пользуется огромной популярностью.
Подобные методы широко применимы практически во всем мире, в том числе и в России. Из-за научного прогресса происходит внедрение таких методов в медицинские исследования и медицинскую практику в целом.
4. Биотехнология
Биотехнология — дисциплина, изучающая возможности использования живых организмов или их систем для решения технологических задач, а еще создания живых организмов с нужными свойствами путем генной инженерии. Биотехнология применяет методы химии, микробиологии, биохимии и, конечно же, молекулярной биологии.
Основные направления развития биотехнологии (принципы биотехнологических процессов внедряют в производство всех отраслей):
- Создание и производство новых видов продуктов питания и кормов для животных.
- Получение и изучение новых штаммов микроорганизмов.
- Выведение новых сортов растений, а также создание средств для защиты растений от болезней и вредителей.
- Применение методов биотехнологии для нужд экологии. Такие методы биотехнологии используют для переработки утилизации отходов, очистки сточных вод, отработанного воздуха и санации почв.
- Изготовление витаминов, гормонов, ферментов, сывороток для нужд медицины. Биотехнологи разрабатывают усовершенствованные лекарственные препараты, которые ранее считались неизлечимыми.
Крупным достижением биотехнологии является генная инженерия.
Генная инженерия — совокупность технологий и методов получения рекомбинантных молекул РНК и ДНК, выделения отдельных генов из клеток, осуществление манипуляций с генами и введение их в другие организмы (бактерий, дрожжи, млекопитающих). Такие организмы способны производить конечные продукты с нужными, измененными свойствами.
Методы генной инженерии направлены на конструирование новых, ранее не существовавших сочетаний генов в природе.
Говоря о достижениях генной инженерии, невозможно не затронуть тему клонирования. Клонирование — это один из методов биотехнологии, применяемый для получения идентичных потомков различных организмов при помощи бесполого размножения.
Иными словами, клонирование можно представить как процесс создания генетически идентичных копий организма или клетки. А клонированные организмы похожи или вовсе идентичны не только по внешним признакам, но и по генетическому содержанию.
Небезызвестная овечка Долли в 1966 году стала первым клонированным млекопитающим. Она была получена за счет пересадки ядра соматической клетки в цитоплазму яйцеклетки. Долли являлась генетической копией овцы-донора ядра клетки. В естественных условиях особь формируется из одной оплодотворенной яйцеклетки, получив по половине генетического материала от двух родителей. Однако при клонировании генетический материал взяли из клетки одной особи. Сначала из зиготы удалили ядро, в котором находится сама ДНК. После чего извлекли ядро из клетки взрослой особи овцы и имплантировали его в ту лишенную ядра зиготу, а затем ее пересадили в матку взрослой особи и предоставили возможность для роста и развития.
Тем не менее не все попытки клонирования оказывались удачными. Параллельно с клонированием Долли эксперимент по замене ДНК был проведен на 273 других яйцеклетках. Но только в одном случае смогло полноценно развиться и вырасти живое взрослое животное. После Долли ученые пробовали клонировать и другие виды млекопитающих.
Одним их видов генной инженерии является редактирование генома.
Инструмент CRISPR/Cas базируется на элементе иммунной защитной системы бактерий, который ученые приспособили для внедрения каких-либо изменений в ДНК животных или растений.
CRISPR/Cas является одним из биотехнологических методов манипулирования отдельными генами в клетках. Существует огромное множество применений такой технологии. CRISPR/Cas позволяет исследователям выяснять функцию разных генов. Для этого нужно просто вырезать исследуемый ген из ДНК и изучить, какие функции организма были затронуты.
Некоторые практические применения системы:
Швейцарские ученые значительно усовершенствовали и модернизировали метод редактирования генома CRISPR/Cas, тем самым расширив его возможности. Тем не менее ученые могли модифицировать только один ген за раз, используя CRISPR/Cas-систему. Но сейчас исследователи Швейцарской высшей технической школы Цюриха разработали метод, с помощью которого возможно одновременно модифицировать 25 генов в клетке.
Для новейшей методики специалисты использовали фермент Cas12a, а не фермент Cas9, применяемый в большинстве методов CRISPR/Cas.
I. Белок - субстрат жизни
Основу жизнедеятельности живых организмов составляют процессы превращения веществ (окисление, восстановление, расщепление, синтез). В течение жизни каждая клетка усваивает и продуцирует различные вещества, строит и обновляет свои структуры, выполняет определенные функции. Основным строительным материалом в клетке является белок (матриксы цитоплазмы, ядра, митохондрий, пластид; мембранные и немембранные органоиды) - это структурные белки.
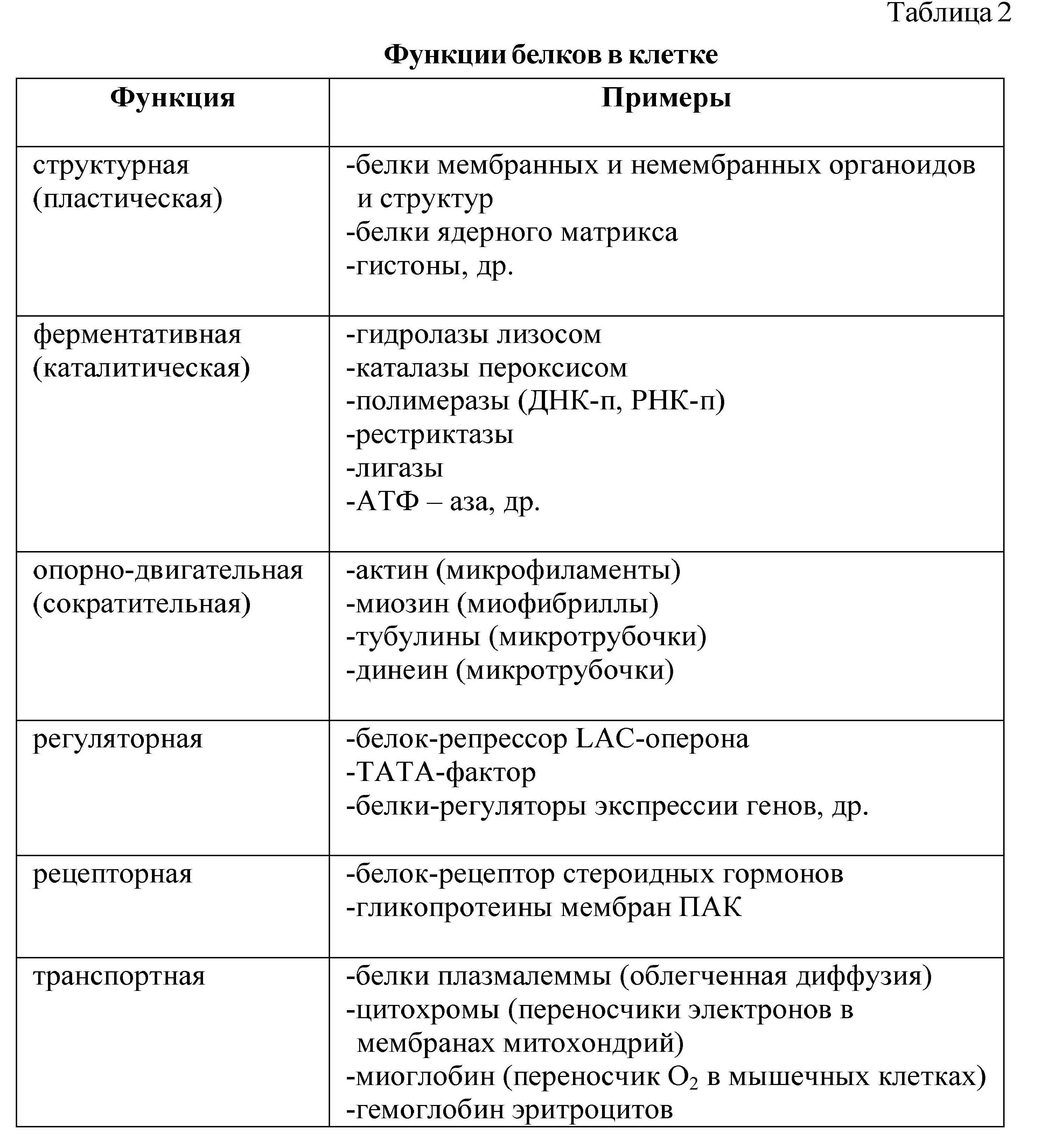
Особую группу структурных белков составляют сократительные белки, которые формируют опорно-двигательные элементы клетки (микротрубочки, микрофиламенты, микрофибриллы) и определяют движение клеток, деление, фагоцитоз и др. К таким белкам относятся: актин, тубулин, миозин и др. Превращение веществ в клетке осуществляется с помощью ферментов, химической основой которых являются белки. Таким образом, структурная (пластическая) и каталитическая (ферментативная) функции являются главными функциями белка в любой клетке, именно белки определяют и строение клетки и процессы ее жизнедеятельности. Кроме этого, белки выполняют многочисленные функции в клетке и организме (табл. 2).
Белки являются универсальными молекулами и имеют принципиально сходное строение у животных, растений, бактерий и вирусов. Каждый белок в своей первичной структуре представляет собой цепочку аминокислот, соединенных пептидными связями (полипептид). Но в то же время, организмы разных видов различаются своими белками; разные ткани одного и того же организма построены из разных белков (соединительная ткань - коллаген; мышечная ткань - актин, миозин, миоглобин; ногти, волосы - кератин и т.д.); имеются индивидуальные отличия организмов по строению белков - следовательно, белки обладают специфичностью. Специфичность белков обусловлена особенностью первичной структуры. Полипептидные цепи различаются между собой набором аминокислот, последовательностью их расположения и количеством. Разнообразие белков огромно.
II. Нуклеиновые кислоты
В многоклеточном организме клетки дифференцируются и поэтому, клетки одной ткани сходны, а клетки разных тканей различаются по морфологии и функциям. При делении каждая клетка образует себе подобные дочерние клетки (из клеток печени образуются клетки печени; из клеток кожи - клетки кожи). Чтобы синтезировать белки, характерные для данного типа клеток, чтобы воспроизводить себе подобных - необходимо иметь информацию, заключенную в каком-то материальном субстрате, которую: а) можно использовать в процессе жизнедеятельности и б) передавать дочерним клеткам при делении. Это обеспечивает преемственность в строении и функции клеток и организмов в поколениях. Материальным субстратом - носителем генетической информации является ДНК (у некоторых вирусов - РНК). Реализация генетической информации происходит с участием различных РНК (мРНК, тРНК, рРНК). Нуклеиновые кислоты - биополимеры, состоящие из мономеров-нуклеотидов. Любой нуклеотид состоит из трёх частей: углевода, остатка фосфорной кислоты и азотистого основания. Каждая молекула нуклеиновой кислоты - это определённые последовательности нуклеотидов. При соединении нуклеотидов в цепь образуются связи между углеводом и остатком фосфорной кислоты. Углеродный атом в 5 положении рибозы (дезоксирибозы) одного нуклеотида соединяется через фосфатную группу с углеродным атомом в 3 положении сахара предыдущего нуклеотида.
Таким образом, первый нуклеотид в цепи имеет свободный углеродный атом в 5 положении, а последний - в 3 положении (рис. 9), поэтому концы
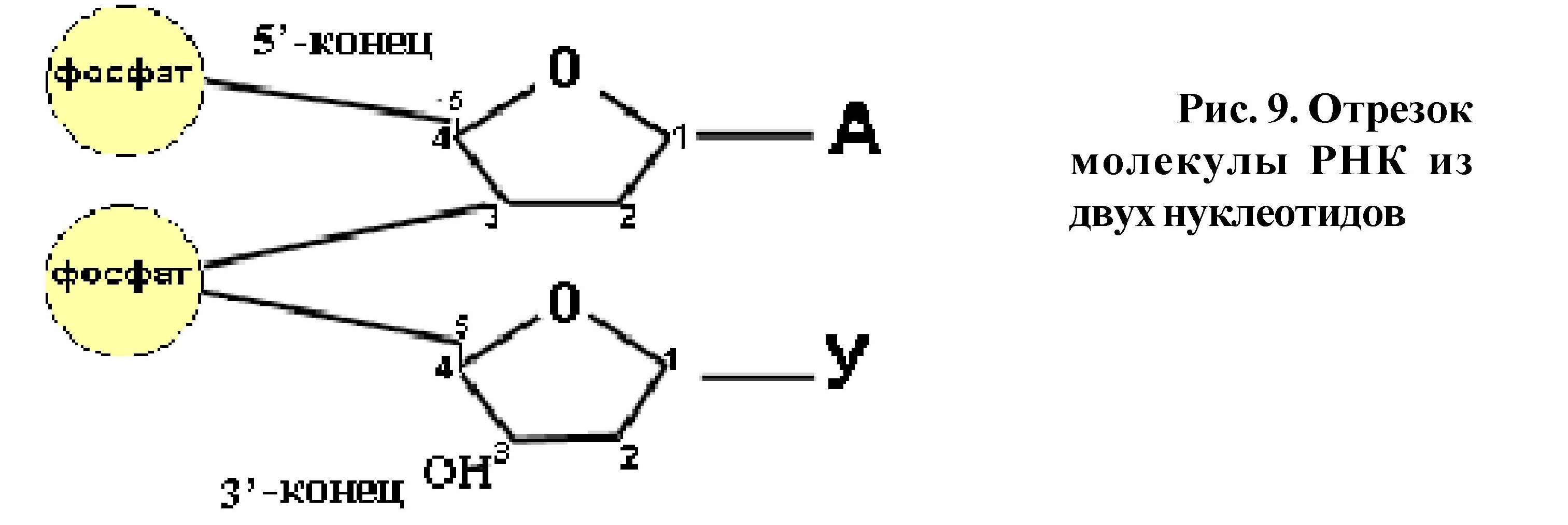
полинуклеотидных цепей обозначаются как 5/ и 3/. В молекуле ДНК две полинуклеотидные цепи, они антипараллельны, то есть там, где у одной цепи 5/ конец - у второй - 3/ конец и наоборот.
Принципиально строение ДНК и РНК сходно, но есть и отличия: молекула ДНК состоит из двух полинуклеотидных цепей (РНК - одна цепь), в состав ДНК входит углевод дезоксирибоза (РНК - рибоза), в состав ДНК входят азотистые основания - аденин, тимин, гуанин, цитозин (в РНК вместо тимина входит урацил). Нуклеотиды одной цепи ДНК (РНК) различаются между собой только азотистым основанием. Две цепи нуклеотидов ДНК соединяются водородными связями по правилу комплементарности: А-Т; Г-Ц.
ДНК содержится в ядре клетки, РНК - в ядре (в основном, в ядрышке) и цитоплазме (гиалоплазма, рибосомы). Кроме того, некоторые органоиды имеют собственную ДНК и все виды РНК (митохондрии, пластиды).
ДНК была открыта в 1869 году (Мишер), но только в 1953 году было расшифровано строение этой молекулы (Уотсон и Крик).
Основная биологическая роль ДНК состоит в хранении, передаче и самовоспроизведении наследственной (генетической) информации.
Строение ДНК универсально (принципиально одинаково у всех живых организмов), но разные молекулы ДНК различаются между собой.
Специфичность ДНК зависит от нуклеотидного состава, последовательности нуклеотидов, количества нуклеотидов. Таким образом, от того какие нуклеотиды входят в состав молекулы, как они расположены и сколько их, зависит объём информации и её смысл.
В строении ДНК содержится информация о структуре белков организма и рибонуклеиновых кислот (тРНК, рРНК).
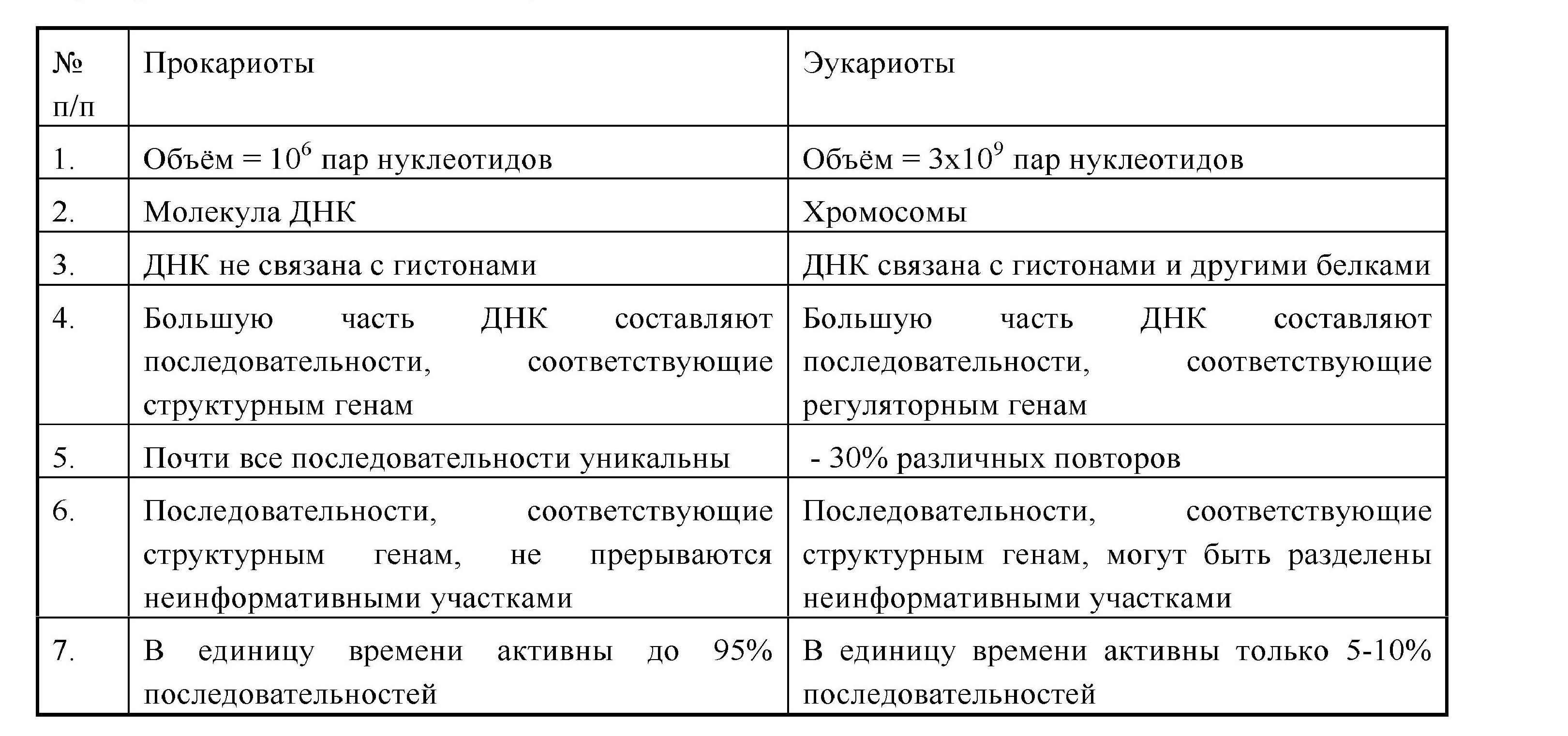
Наследственный аппарат организован по-разному у вирусов, прокариот и эукариот. У вирусов - это может быть молекула ДНК или РНК (различной структурной организации).
У прокариот генетический аппарат представлен двухцепочечной кольцевой молекулой ДНК (нуклеоид, генофор), в которой содержится основная видовая наследственная информация, и плазмоном - совокупностью автономных генетических элементов. Это мелкие кольцевые молекулы ДНК - плазмиды и эписомы, содержащие ограниченную информацию о некоторых признаках данного организма (в плазмидах R находятся гены устойчивости к антибиотикам;эписомы F определяют способность к размножению). Плазмиды и эписомы способны к репликации и перемещению из клетки в клетку при конъюгации.
У эукариот генетический аппарат представлен надмолекулярными структурами - хромосомами, химической основой которых является хроматин (ДНК + белки). Хроматин может быть конденсирован, неактивный - гетерохроматин, или деконденсирован, активный - эухроматин (см. стр. 24). Не вся ДНК эукариот является информативной. Большая часть ее представлена регуляторными последовательностями. Многие участки повторяются в геноме (умеренные и высокие повторы).
Основные различия в организации генетического материала у про- и эукариот сведены в таблицу 3.
III. Генетический код, его характеристика
Смысл генетической информации зашифрован в молекуле ДНК. Генетический код - это система записи генетической информации, которая используется клеткой в процессе жизнедеятельности. Другими словами - это система расположения нуклеотидов в молекуле ДНК,
определяющая последовательность аминокислот в молекуле белка (правило коллинеарности). Единицей генетического кода является триплет нуклеотидов в молекуле ДНК (кодон), который соответств ует одной аминокислоте.
Генетический код характеризуется:
а) универсальностью (другого способа записи генетической информации в природе нет)
б) триплетностью (единица генетического кода - триплет нуклеотидов - кодон)
в) избыточностью (вырожденностью)
г) однозначностью
д) наличием смысловых, терминирующих и инициирующих кодонов.
IV. Реализация генетической информации в клетке
Реализация генетической информации происходит в течение всей жизни клетки в процессе биосинтеза белков, характерных для данного вида организмов (клеток).
Интенсивность биосинтеза белка наибольшая в интерфазе, снижается к началу деления, почти нулевая при делении и возрастает сразу после деления. Биосинтез белка можно разделить на два этапа: транскрипция (происходит в ядре на ДНК) и трансляция (происходит в цитоплазме на рибосомах).
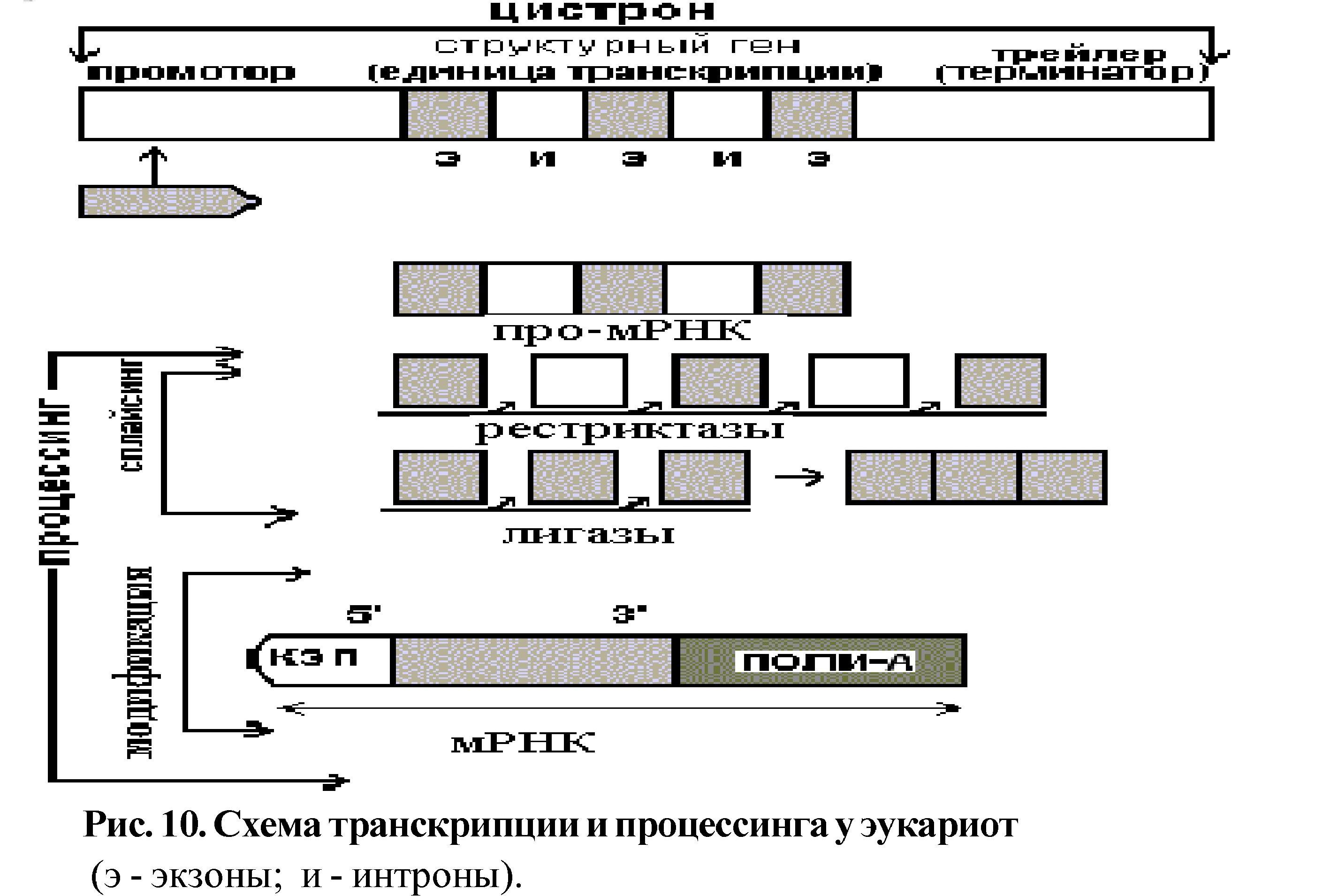
Функциональной единицей, которая участвует в транскрипции, является цистрон - отрезок ДНК состоящий из трёх частей:
а) промотор (около 40 пар последовательностей), с которым связывается фермент РНК-полимераза;
б) последовательности, соответствующие структурному гену;
в) терминальный участок (трейлер), где заканчивается транскрипция.
Биологической сущностью транскрипции является "переписывание" генетической информации с молекулы ДНК на РНК, а химической - синтез молекулы мРНК. Биологической сущностью трансляции является перевод информации с языка нуклеотидов на язык аминокислот (расшифровка генетического кода), а химической - синтез полипептидной цепочки.
Оба процесса относятся к реакциям матричного синтеза, которые характеризуются: наличием молекулы-образца (матрицы), энергии, специфического фермента, выполнением правила комплементарности и протекают в три этапа (инициация, элонгация, терминация). У прокариот синтезированная мРНК сразу может служить матрицей для трансляции.
У эукариот информативные последовательности структурного гена (экзоны) разделены неинформативными (интроны). Первичный транскрипт включает как экзоны, так и интроны - это незрелая или про-мРНК. Затем начинается процессинг (созревание мРНК), в результате которого удаляются интроны и образуется зрелая мРНК, состоящая только из экзонов. Процессинг состоит из двух этапов: сплайсинга и модификации (рис. 10).
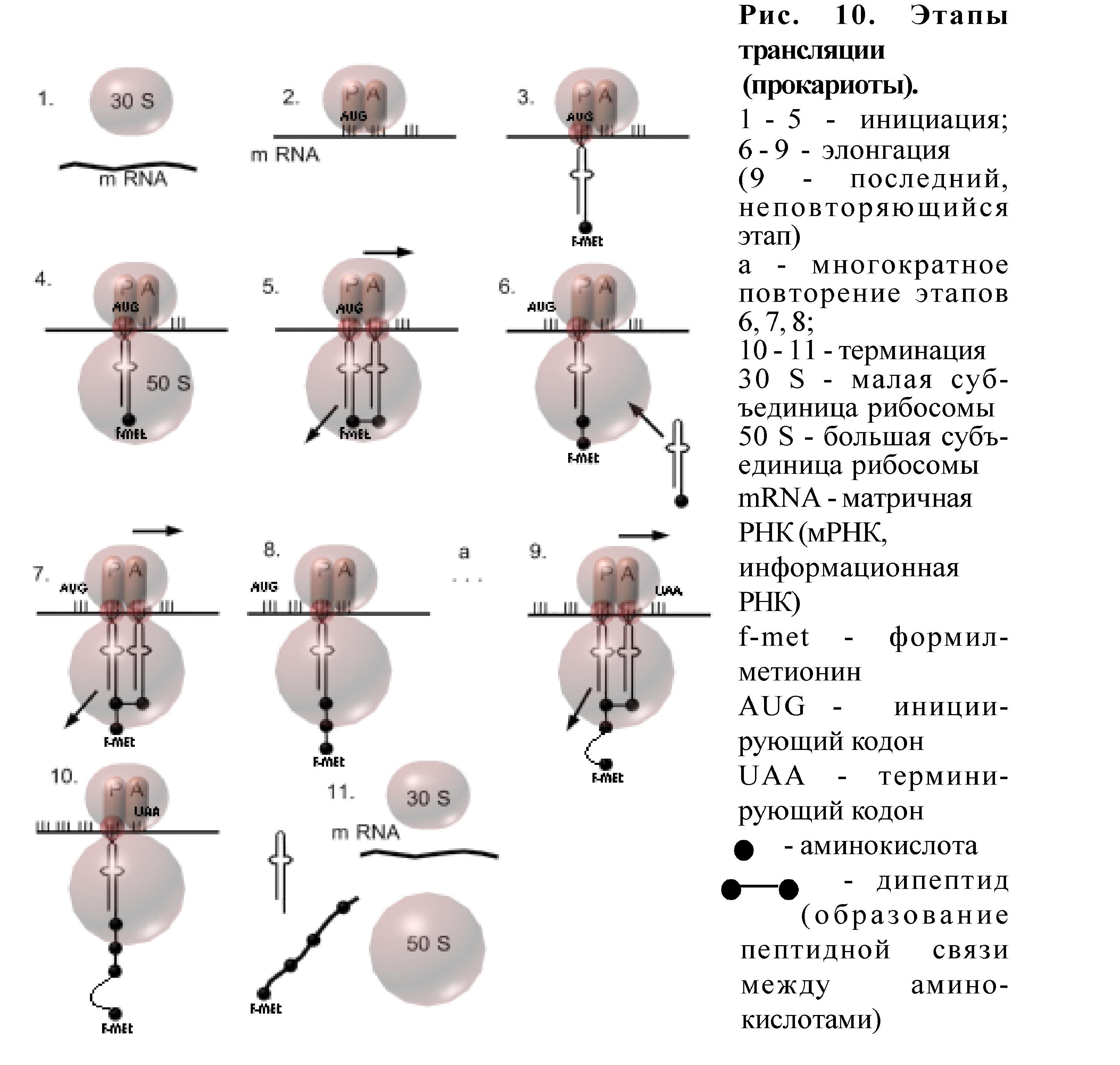
Трансляция (рис.10)
также состоит из инициации, элонгации и терминации.
Местом трансляции являются рибосомы. В рибосомах есть два активных функциональных центра - пептидильный и аминоацильный. Не работающая рибосома диссоциирована на две субъединицы: малую и большую.
Инициация трансляции начинается со связывания мРНК с малой субъединицей рибосомы, причём необходимо чтобы в её пептидильном центре оказался триплет АУГ - это инициирующий кодон. С этим кодоном связывается тРНК-f-метионин, а затем малая и большая субъединицы рибосомы объединяются. Рибосома готова к работе. В аминоацильном центре рибосомы находится другой триплет нуклеотидов мРНК, с которым может связаться тРНК, имеющая комплементарный антикодон. Когда это произойдёт, то между двумя аминокислотами (одна - f-метионин в пептидильном центре, вторая - в аминоацильном центре) возникает пептидная связь - образуется дипептид, инициация завершилась. Рибосома передвигается по мРНК на один триплет, который оказывается в аминоацильном центре, тРНК из него перемещается в пептидильный центр; она связана с дипептидом, а первая тРНК уходит в цитоплазму. Аминоацильный центр свободен, в нем находится новый кодон, с которым может связаться тРНК с комплементарным антикодоном. Так, передвигаясь по мРНК, рибосома "прочитывает" информацию, переводит её на язык аминокислот и полипептидная цепь наращивается. Это - элонгация. Элонгация происходит до тех пор, пока на пути рибосомы в А-центре не окажется кодон-терминатор. Тогда полипептидная цепь отсоединяется от рибосомы, мРНК тоже отделяется от рибосомы, рибосома диссоциируется на субъединицы, происходит терминация Этапы трансляции (рис.10):
Инициация 1-5. Начало матричного синтеза (трансляции) происходит поэтапно:
1- связывание мРНК с малой (30 S) субъединицей рибосомы
2 - установка в пептидильном центре (Р) инициирующего кодона АУГ (AUG)
3 - связывание тРНК с аминокислотой формил-метионин (тРНК - f-met) с кодоном АУГ (образование инициирующего комплекса)
4 - присоединение большой (50 S) субъединицы рибосомы
5.1-образование комплекса кодон-антикодон в аминоацильном (А) центре
5.2 - образование пептидной связи между формил-метионином и второй аминокислотой(образование дипептида)
5.3 - транспозиция рибосомы (перемещение) по мРНК на один триплет (при этом первая тРНК покидает рибосому, вторая тРНК, с которой связан дипептид, перемещается из А - в Р центр, а в А центре появляется новый кодон).
Элонгация 6 - 9. Углубление и ускорение процесса трансляции, результатом чего является наращивание полипептидной цепи. Состоит из многократно повторяющихся этапов:
6 - транспортировка аминокислот в рибосому с помощью тРНК
7.1-образование комплекса кодон-антикодон в А центре
7.2 - образование пептидной связи между аминокислотами
8 - транспозиция рибосомы по мРНК на один триплет
Терминация 10-11. Окончание трансляции.
10 - появление в А центре после очередной транспозиции рибосомы терминирующего кодона (УАА, УАГ, УГА)
11.1-в Р- центре дестабилизируется и утрачивается связь между тРНК и мРНК
11.2 - полипептид отщепляется от тРНК
11.3 - мРНК покидает рибосому
11.4 - диссоциация рибосомы на субъединицы
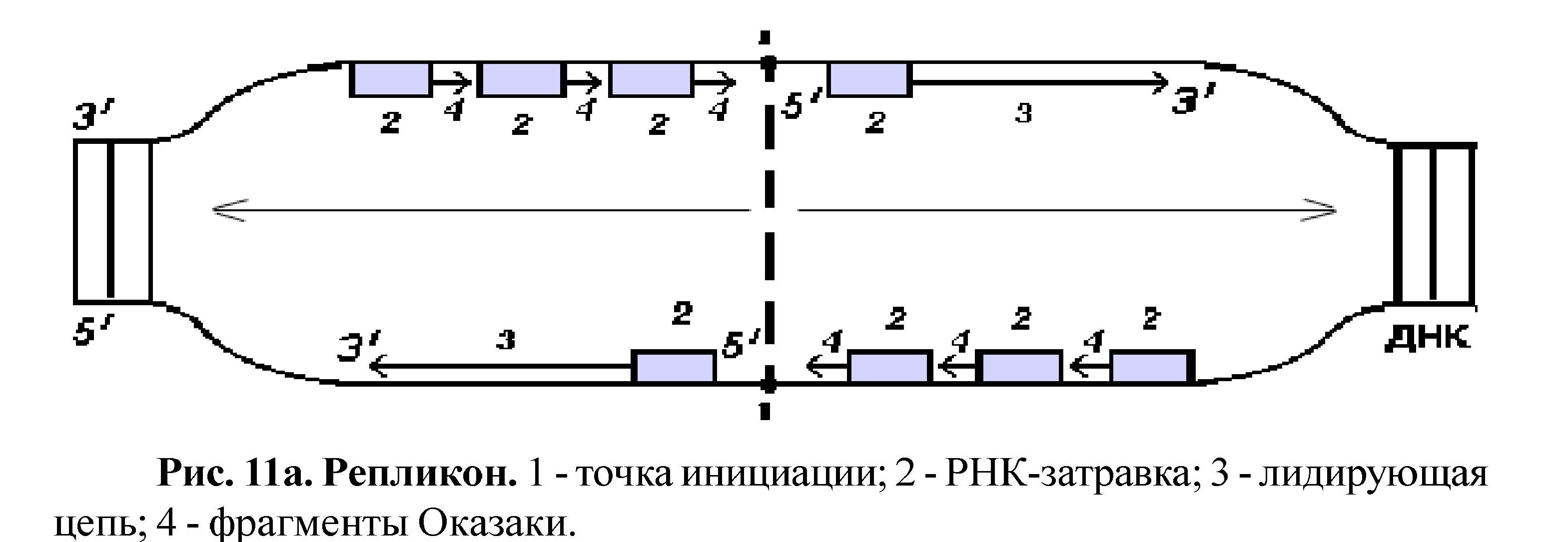
V. Репликация ДНК
Самовоспроизведение (ауторепродукция) ДНК называется репликацией. Репликация ДНК происходит перед делением клетки; в результате этого процесса содержание ДНК в клетке удваивается, а так как репликация протекает по правилу комплементарности, то две дочерние молекулы идентичны материнской и друг другу. Следовательно, каждая новая клетка получает информацию в количественном и качественном отношении одинаковую с родительской клеткой. Разъединение двух цепей ДНК у эукариот начинается одновременно в нескольких участках (у прокариот в одном месте). Такой участок называется - репликон (рис. 11а). В эукариотической клетке может быть более 2000 репликонов. Репликация - это реакция матричного синтеза; матрицей служит молекула ДНК, основными ферментами являются ДНК-полимераза, лигаза, рестриктаза.
Начинается процесс с разрыва водородных связей между азотистыми основаниями ДНК на участке, включающем около 300 пар нуклеотидов - это место называется точка инициации. Так как разъединение цепей ДНК от точки инициации идет вправо и влево одновременно, цепи ДНК антипараллельны, а фермент ДНК-полимераза может работать только в одном направлении (соединяя нуклеотиды от 5 углерода последующего к 3 углероду предыдущего), то синтез дочерних цепей идет по-разному на разных участках одного репликона. Одна цепь - лидирующая, синтезируется непрерывно, а вторая - отстающая, синтезируется фрагментарно (Рис. 11б).
На цепи 3/_5/ рядом с точкой инициации есть особая последовательность нуклеотидов - сайт инициации, на котором синтезируется небольшая молекула РНК (РНК-затравка). У РНК-затравки свободен 3/ - конец, к которому присоединяется первый нуклеотид ДНК, к нему второй и т. д. В результате синтезируется лидирующая дочерняя цепь. На противоположной, антипараллельной цепи (5/-3/) сайта инициации нет и проходит время, пока в
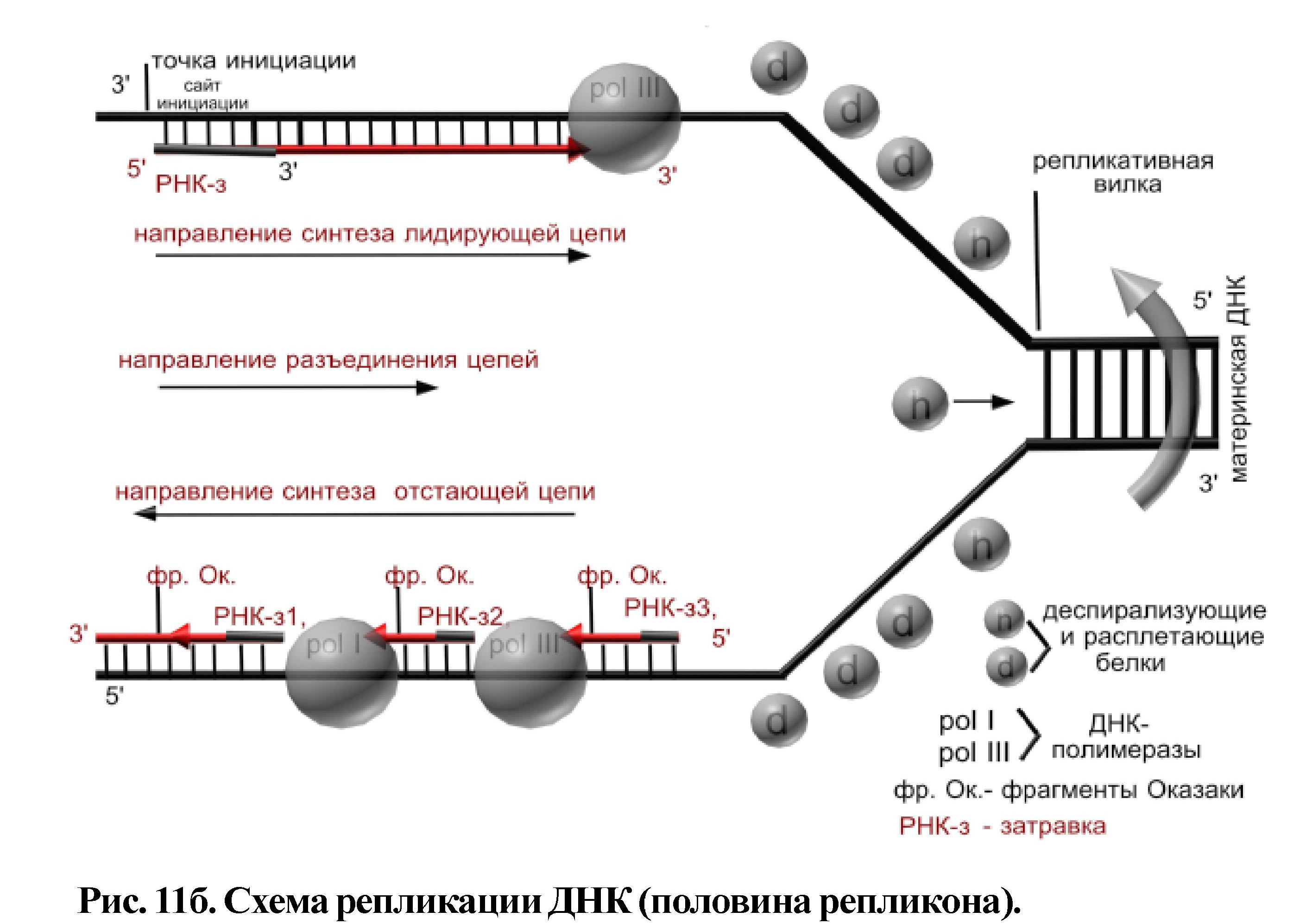
результате разрыва водородных связей обнаружится такой сайт; РНК-затравка синтезируется и от неё в сторону противоположную направлению разъединения ДНК синтезируется небольшой фрагмент дочерней цепи. После разъединения следующего участка молекулы ДНК, следующая молекула РНК-затравка находит свой сайт и синтезируется новый фрагмент дочерней цепи ДНК в направлении 5/ -3/ и т.д. Таким образом, эта цепь синтезируется небольшими фрагментами (фрагменты Оказаки) и отстаёт во времени. На другой половине репликона, где разъединение цепей ДНК идёт в другую сторону, также, в одном направлении дочерняя цепь синтезируется непрерывно, в другом - фрагментарно. Затем рестриктазы вырезают РНК-затравки (одну - из лидирующей цепи и от каждого фрагмента Оказаки на отстающей цепи), ДНК-полимераза достраивает молекулу ДНК на местах вырезанных РНК-затравок, а лигазы соединяют фрагменты в непрерывную цепь. В каждой новой молекуле ДНК одна цепь старая (материнская), а вторая - новая (дочерняя). Такой способ репликации называется полуконсервативным.
VI. Обратная транскрипция
Представление о направлении потока информации в клетке и последо-вательности процессов получило название центральной догмы молекулярной биологии. Передача генетической информации идёт в направлении

Однако, оказалось, что иногда информация может передаваться от РНК к ДНК. Это явление было изучено у вирусов, генетический аппарат которых представлен не ДНК, а РНК. Это группа ретровирусов, к которым относится вирус гриппа, СПИДа и др. Чтобы после внедрения таких вирусов в клетку хозяина их генетическая информация могла быть использована для синтеза вирусных белков, необходимо на вирусной РНК синтезировать ДНК, с последующим встраиванием ее в геном клетки. Этот процесс идёт под контролем фермента ревертазы (обратной транскриптазы) и называется обратной транскрипцией. Таким образом, направление потока генетической информации в клетке в окончательном виде выглядит так:

Открытие явления обратной транскрипции сыграло большую роль в развитии генной инженерии, микробиологии. С помощью ревертаз получают важные лекарственные препараты белковой природы (интерферон, гамма- глобулин и др.), вводя в микробную клетку мРНК человека с информацией о строении этих белков.
Читайте также: