
Характеристика внутриклеточной и внеклеточной среды возбудимой клетки кратко
Обновлено: 02.07.2024
Строение клеточной мембраны нейрона. Мембранный потенциал покоя
Внеклеточная и внутриклеточная жидкости представляют собой водно-солевые растворы, в которых множество растворимых молекул диссоциируют на положительно и отрицательно заряженные атомы,— ионы. Ионы и молекулы водных растворов находятся в состоянии постоянного возбуждения и осуществляют процесс диффузии—движения из областей высокой концентрации в область меньшей концентрации. Диффузия ионов происходит не только по градиенту концентрации, но и под влиянием электрических градиентов. Положительно заряженные ионы, например ионы натрия (Na + ) и калия (К + ), называют катионами, поскольку в электрическом поле они движутся к катоду.
Отрицательно заряженные ионы, например ионы хлора (Cl - ), называют анионами, так как они движутся к аноду. Одноименные заряды (например, Na + и К + ) отталкиваются, а разноименные (например, Na + и Cl - ) — притягиваются.
Клеточную мембрану можно рассматривать в качестве электрического конденсатора, поскольку она состоит из двух противоположно заряженных слоев, между которыми расположен изолирующий липидный слой. Электрический потенциал тканевой жидкости на плазматической мембране стремится к нулю (0 мВ) за счет нейтрализующих влияний анионов Cl - на катионы Na + (и другие катионы), а в цитозоле — за счет влияний белков-анионов на катионы К + .
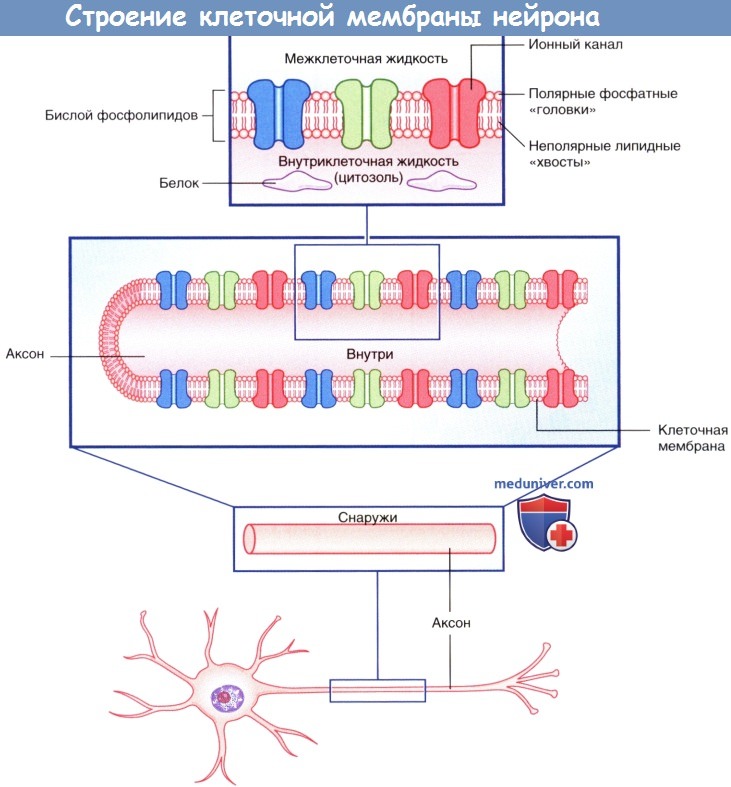
Строение клеточной мембраны нейрона.
Изображены мембранные белки, образующие ионные каналы.
а) Ионные каналы. Ионные каналы представляют собой трансмембранные белки, в центре которых находятся отверстия, по которым осуществляется транспорт ионов через клеточную мембрану. Большинство ионных каналов селективно пропускает определенные ионы, например, Na + , К + , Cl - . Выделяют несколько видов ионных каналов, из которых наиболее значимы первые три типа из следующих:
• Неуправляемые (независимые) ионные каналы постоянно находятся в открытом состоянии и осуществляют трансмембранный перенос ионов, создавая потенциал покоя на мембранах нейронов.
• Потенциалозависимые (потенциал-управляемые) ионные каналы содержат потенциалчувствительную цепь аминокислот, регулирующую открытие и закрытие отверстий ионных каналов в зависимости от изменений мембранного потенциала. Потенциал-управляемые ионные каналы играют важную роль в формировании потенциала действия.
• Энергозависимые транспортеры — ионные обменники (насосы) — обеспечивают постоянство концентраций ионов. Натрий-калиевый насос обеспечивает поддержание потенциала покоя.
• Хемоуправляемые (медиаторозависимые) ионные каналы нервной системы обеспечивают временное преобразование мембранного потенциала. В основном эти каналы представлены на постсинаптических мембранах. Активация хемоуправляемых ионных каналов может осуществляться молекулами медиаторов напрямую или опосредованно.
• Механочувствительные каналы активируются под действием физических раздражителей, вызывая деполяризацию мембраны и образование потенциалов действия, что обеспечивает восприятие определенных сигналов нервной системой. Каждый рецептор осуществляет преобразование специфических сигналов, например изменения длины или сократимости мышечных волокон, температурной и тактильной чувствительности кожи, хемочувствительности полости носа и рта или электромагнитных импульсов сетчатки.
На рисунке ниже изображены три неуправляемых ионных канала, обеспечивающих развитие потенциала покоя.
Если бы пассивная диффузия различных ионов через соответствующие им каналы осуществлялась свободно в одинаковом объеме, то не существовало бы разницы зарядов внутри и снаружи мембраны. В действительности проницаемость плазматической мембраны для ионов К + и Cl - намного выше, чем для ионов Na + , поскольку количество ионных каналов различается: калиевые каналы наиболее многочисленны, количество хлорных каналов вдвое меньше, а натриевые каналы встречают реже всего.
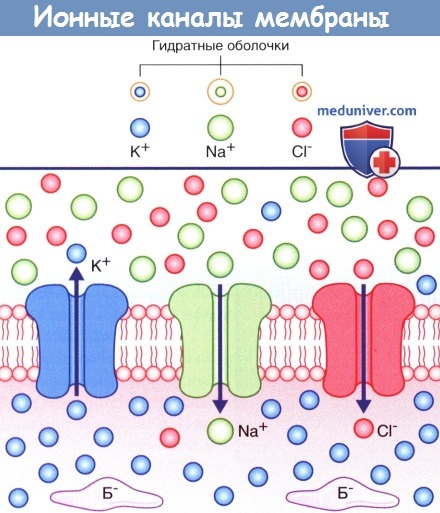
В состоянии покоя ионы Na + и Cl - сконцентрированы на внешней стороне плазматической мембраны за счет взаимного притяжения и образования толстой гидратной оболочки вокруг ионов Na + .
Ионы К + сконцентрированы на внутренней стороне плазматической мембраны за счет притяжения к белкам-анионам (Б - ).
Стрелками показано направление ионных градиентов концентрации.
б) Мембранный потенциал покоя. Мембранный потенциал невозбужденного нейрона (находящегося в состоянии покоя) формируется за счет разницы концентраций ионов Na + и К + во внеклеточной жидкости и цитозоле. Концентрация ионов К + в цитозоле выше, чем во внеклеточной жидкости в 20 раз, в то время как концентрация ионов Na + и Cl - во внеклеточной жидкости выше, чем в цитозоле в 10 и 3,8 раз, соответственно. Это обусловлено тем, что ионы К + под действием химических движущих сил переносятся из клетки в межклеточную жидкость, а ионы Na+ и Cl - — в обратном направлении.
На рисунке ниже показан вольтметр, электроды которого погружены в окружающую аксон внеклеточную жидкость. Один из электродов находится в стеклянной пипетке с тонким кончиком. В левой части рисунка, где оба электрода погружены во внеклеточную жидкость, разница электрического напряжения отсутствует, и вольтметр регистрирует нулевой показатель. В правой части рисунка пипетка проникает через плазматическую мембрану аксона в цитозоль, что позволяет оценить электрический заряд внутриклеточной жидкости. В данном случае разность электрических потенциалов составляет -70 мВ, однако у разных нейронов она может варьировать от -60 до -80 мВ.
Этот показатель отражает мембранный потенциал покоя, т.е. потенциал мембраны аксона, который не проводит электрические импульсы.
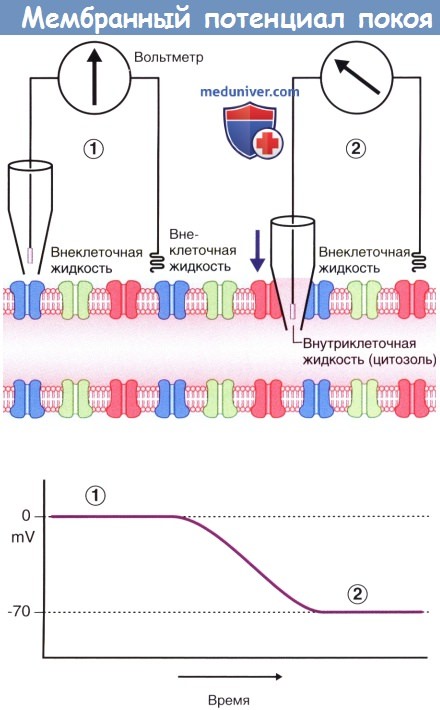
Мембранный потенциал покоя.
(1) Оба электрода вольтметра погружены во внеклеточную жидкость, окружающую аксон. Конец левого электрода находится в стеклянной пипетке.
Разница электрических потенциалов не регистрируется; на графике отмечены нулевые показатели.
(2) При опускании пипетки (направление указано стрелкой) производят пункцию плазматической мембраны для забора внутриклеточной жидкости.
Регистрируется разница электрических потенциалов величиной -70 мВ.
в) Проницаемость мембраны в состоянии покоя: 1. Ионы К + . На уровне ионного канала перемещение ионов К + осуществляется под электрическими влияниями притягивающего действия белков-анионов на внутренней стороне мембраны и отталкивающего действия ионов Na + на ее наружной стороне; при отсутствии этих факторов концентрация ионов К + на разных сторонах плазматической мембраны была бы одинаковой.
Концентрационный градиент на внешней стороне плазматической мембраны и электрический градиент потенциала на внутренней ее стороне уравновешиваются, когда мембранный потенциал достигает определенной величины, которую называют равновесным потенциалом для ионов К + (Ek). Его величина может быть рассчитана из уравнения Нернста, которое позволяет представить электрический градиент потенциала иона через его концентрационный градиент на основании законов термодинамики:
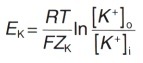
где Ek — равновесный потенциал для ионов К + (мВ)
R — газовая постоянная (8,31 Дж/(моль/°К)
Т — абсолютная температура в градусах Кельвина (310 К = 37 °С) F — число Фарадея (96500 Кл/моль)
Zk — валентность ионов К + (+1)
ln — натуральный логарифм
[К + ]0 — концентрация ионов K + с наружной стороны мембраны
[К + ]i — концентрация ионов К + с внутренней стороны мембраны.
Переведем натуральный логарифм в десятичный и рассчитаем значение показателя дроби:
Рассчитаем эти же значения для ионов Na + и Cl - :

Для расчета мембранного потенциала покоя применяют уравнение Гольдмана, учитывающее проницаемость мембраны для трех основных ионов и их приблизительное количество.
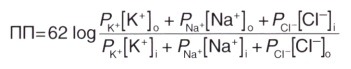
где ПП — потенциал покоя
62 — RT/Fx2,3 (постоянная для перевода натурального логарифма в десятичный)
Р — константа проницаемости мембраны для каждого из ионов (соответствует приблизительному количеству ионных каналов) о, i—концентрации ионов на внешней и внутренней сторонах мембраны, соответственно; концентрация отрицательно заряженных ионов Cl - внутри клетки расположена в числителе, а снаружи — в знаменателе дроби, поскольку log (X/Y) = log (Y/X).
Квадратными скобками обозначены концентрации ионов.
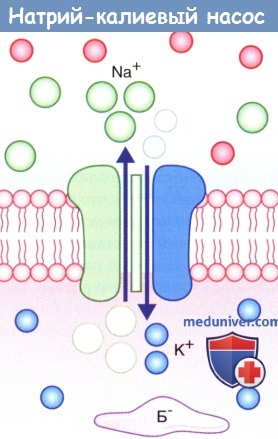
Натрий-калиевый насос.
На рисунке показан одновременный перенос трех ионов Na + из клетки в обмен на два иона К + в клетку.
Б - — белок-анион.
2. Натрий-калиевый насос. Под влиянием градиента концентрации происходит непрерывное перемещение ионов Na + внутрь клетки, а ионов К + — наружу, что нарушает постоянство потенциала покоя. Поддержание этого равновесия обеспечивает натрий-калиевый насос, корректирующий пассивное движение ионов. Натрий-калиевый насос представляет собой канал, способный перемещать ионы Na + наружу, а ионы К + — внутрь клетки. Во время работы насоса происходит перенос трех ионов Na + из клетки на каждые два иона К + в клетку.
Перенос обоих катионов осуществляется против градиента концентрации за счет энергии превращения АТФ в АДФ ферментом АТФазой. Активация этого фермента происходит при повышении концентрации ионов Na + в цитозоле.
Аксональная дегенерация при рассеянном склерозе возникает вследствие нарушения работы натрий-калиевого насоса в демиелинизированной аксолемме, что вызывает увеличение концентрации ионов Na + , которое в свою очередь приводит к повышению высвобождения ионов Са 2+ из внутриклеточного пула.

Обзор
Потенциал покоя. В мембране открыты калиевые ионные каналы постоянного тока, закрыты натриевые каналы, работает насос-обменник (Na + /K + -АТФаза).
Автор
Редакторы

Давайте разбираться подробнее.
Зачем нам нужно знать, что такое потенциал покоя и как он возникает?
На эти вопросы можно ответить только в том случае, если узнать, как клетка создаёт себе разность электрических потенциалов (потенциал покоя) на мембране.
Совершенно очевидно, что для понимания того, как работает нервная система, необходимо вначале разобраться, как работает её отдельная нервная клетка — нейрон. Главное, что лежит в основе работы нейрона — это перемещение электрических зарядов через его мембрану и появление вследствие этого на мембране электрических потенциалов. Можно сказать, что нейрон, готовясь к своей нервной работе, вначале запасает энергию в электрической форме, а затем использует ее в процессе проведения и передачи нервного возбуждения.
Таким образом, наш самый первый шаг к изучению работы нервной системы — это понять, каким образом появляется электрический потенциал на мембране нервных клеток. Этим мы и займёмся, и назовём этот процесс формированием потенциала покоя.
В норме, когда нервная клетка находится в физиологическом покое и готова к работе, у неё уже произошло перераспределение электрических зарядов между внутренней и наружной сторонами мембраны. За счёт этого возникло электрическое поле, и на мембране появился электрический потенциал — мембранный потенциал покоя.
Таким образом, мембрана оказывается поляризованной. Это означает, что она имеет разный электрический потенциал наружной и внутренней поверхностей. Разность между этими потенциалами вполне возможно зарегистрировать.
В этом можно убедиться, если ввести внутрь клетки микроэлектрод, соединённый с регистрирующей установкой. Как только электрод попадает внутрь клетки, он мгновенно приобретает некоторый постоянный электроотрицательный потенциал по отношению к электроду, расположенному в окружающей клетку жидкости. Величина внутриклеточного электрического потенциала у нервных клеток и волокон, например, гигантских нервных волокон кальмара, в покое составляет около −70 мВ. Эту величину называют мембранным потенциалом покоя (МПП). Во всех точках аксоплазмы этот потенциал практически одинаков.
Ноздрачёв А.Д. и др. Начала физиологии [5].
Ещё немного физики. Макроскопические физические тела, как правило, электрически нейтральны, т.е. в них в равных количествах содержатся как положительные, так и отрицательные заряды. Зарядить тело можно, создав в нем избыток заряженных частиц одного вида, например, трением о другое тело, в котором при этом образуется избыток зарядов противоположного вида. Учитывая наличие элементарного заряда (e), полный электрический заряд любого тела можно представить как q = ±N×e, где N — целое число.
Потенциал электростатического поля φ определяется как отношение потенциальной энергии W пробного заряда q к величине этого заряда: φ = W/q, откуда следует, что потенциал численно равен потенциальной энергии, которой обладает в данной точке поля единичный положительный заряд. Единицей измерения потенциала служит вольт (1 В) [4]. В некоторых случаях потенциал электрического поля нагляднее определяется как физическая величина, численно равная работе внешних сил против сил электрического поля E при перемещении единичного положительного точечного заряда из бесконечности в данную точку. Последнее определение удобно записать следующим образом:

В электрофизиологии кроме потенциала покоя рассматриваются и другие электрические потенциалы: локальные постсинаптические и рецепторные потенциалы (возбуждающие и тормозные), электротонические и следовые потенциалы, миниатюрные потенциалы концевой пластинки, концентрационный потенциал и потенциал действия [5].
Потенциал покоя — это разность электрических потенциалов, имеющихся на внутренней и наружной сторонах мембраны, когда клетка находится в состоянии физиологического покоя. Его величина измеряется изнутри клетки, она отрицательна и составляет в среднем −70 мВ (милливольт), хотя в разных клетках может быть различной: от −35 мВ до −90 мВ.
Важно учитывать, что в нервной системе электрические заряды представлены не электронами, как в обычных металлических проводах, а ионами — химическими частицами, имеющими электрический заряд. И вообще в водных растворах в виде электрического тока перемещаются не электроны, а ионы. Поэтому все электрические токи в клетках и окружающей их среде — это ионные токи.
Итак, изнутри клетка в покое заряжена отрицательно, а снаружи — положительно. Это свойственно всем живым клеткам, за исключением, разве что, эритроцитов, которые, наоборот, заряжены отрицательно снаружи. Если говорить конкретнее, то получается, что снаружи вокруг клетки будут преобладать положительные ионы (катионы Na + и K + ), а внутри — отрицательные ионы (анионы органических кислот, не способные свободно перемещаться через мембрану, как Na + и K + ).
Теперь нам всего лишь осталось объяснить, каким же образом всё получилось именно так. Хотя, конечно, неприятно сознавать, что все наши клетки кроме эритроцитов только снаружи выглядят положительными, а внутри они — отрицательные.
Сущность формирования потенциала покоя
Попробуем разобраться, откуда берётся электрический заряд нервных клеток, хотя их никто не трёт, как это делают физики в своих опытах с электрическими зарядами.
Здесь исследователя и студента поджидает одна из логических ловушек: внутренняя отрицательность клетки возникает не из-за появления лишних отрицательных частиц (анионов), а, наоборот, из-за потери некоторого количества положительных частиц (катионов)!
Так куда же деваются из клетки положительно заряженные частицы? Напомню, что это покинувшие клетку и скопившиеся снаружи ионы натрия — Na + — и калия — K + .
Главный секрет появления отрицательности внутри клетки
Сразу откроем этот секрет и скажем, что клетка лишается части своих положительных частиц и заряжается отрицательно за счёт двух процессов:
Эти два процесса нам и надо объяснить.
Первый этап создания внутренней отрицательности: обмен Na + на K +
Но ведь при обмене одного положительного заряда (Na + ) на другой такой же положительный заряд (K + ) никакого дефицита положительных зарядов в клетке возникать не может! Правильно. Но, тем не менее, из-за этого обмена в клетке остаётся очень мало ионов натрия, потому что они почти все ушли наружу. И в то же время клетка переполняется ионами калия, которые в неё накачали молекулярные насосы. Если бы мы могли попробовать на вкус цитоплазму клетки, мы бы заметили, что в результате работы насосов-обменников она превратилась из солёной в горько-солёно-кислую, потому что солёный вкус хлорида натрия сменился сложным вкусом довольно-таки концентрированного раствора хлорида калия. В клетке концентрация калия достигает 0,4 моль/л. Растворы хлорида калия в пределах 0,009–0,02 моль/л имеют сладкий вкус, 0,03–0,04 — горький, 0,05–0,1 — горько-солёный, а начиная с 0,2 и выше — сложный вкус, состоящий из солёного, горького и кислого [8].
Кстати, интересно, что клетка не рождается с готовым потенциалом покоя. Ей его ещё надо создать. Например, при дифференцировке и слиянии миобластов потенциал их мембраны изменяется от −10 до −70 мВ, т.е. их мембрана становится более отрицательной — поляризуется в процессе дифференцировки. А в экспериментах на мультипотентных мезенхимальных стромальных клетках костного мозга человека искусственная деполяризация, противодействующая потенциалу покоя и уменьшающая отрицательность клеток, даже ингибировала (угнетала) дифференцировку клеток [1].
- любовь клетки к калию (поэтому клетка насильно затаскивает его к себе);
- любовь калия к свободе (поэтому калий покидает захватившую его клетку).
Механизм насыщения клетки калием мы уже объяснили (это работа насосов-обменников), а механизм ухода калия из клетки объясним ниже, когда перейдём к описанию второго этапа создания внутриклеточной отрицательности. Итак, результат деятельности мембранных ионных насосов-обменников на первом этапе формирования потенциала покоя таков:
- Дефицит натрия (Na + ) в клетке.
- Избыток калия (K + ) в клетке.
- Появление на мембране слабого электрического потенциала (−10 мВ).
Можно сказать так: на первом этапе ионные насосы мембраны создают разность концентраций ионов, или градиент (перепад) концентрации, между внутриклеточной и внеклеточной средой.
Второй этап создания отрицательности: утечка ионов K + из клетки
Итак, что начинается в клетке после того, как с ионами поработают её мембранные натрий-калиевые насосы-обменники?
Из-за образовавшегося дефицита натрия внутри клетки этот ион при каждом удобном случае норовит устремиться внутрь: растворённые вещества всегда стремятся выровнять свою концентрацию во всём объёме раствора. Но это у натрия получается плохо, поскольку ионные натриевые каналы обычно закрыты и открываются только при определённых условиях: под воздействием специальных веществ (трансмиттеров) или при уменьшении отрицательности в клетке (деполяризации мембраны).
В то же время в клетке имеется избыток ионов калия по сравнению с наружной средой — потому что насосы мембраны насильно накачали его в клетку. И он, тоже стремясь уравнять свою концентрацию внутри и снаружи, норовит, напротив, выйти из клетки. И это у него получается!
Ионы калия K + покидают клетку под действием химического градиента их концентрации по разные стороны мембраны (мембрана значительно более проницаема для K + , чем для Na + ) и уносят с собой положительные заряды. Из-за этого внутри клетки нарастает отрицательность.
К + -каналы утечки постоянно открыты при нормальных значениях мембранного потенциала покоя и проявляют взрывы активности при сдвигах мембранного потенциала, которые длятся несколько минут и наблюдаются при всех значениях потенциала. Усиление К + -токов утечки ведёт к гиперполяризации мембраны, тогда как их подавление — к деполяризации. . Однако, существование канального механизма, ответственного за токи утечки, долгое время оставалось под вопросом. Только сейчас стало ясно, что калиевая утечка — это ток через специальные калиевые каналы.
Зефиров А.Л. и Ситдикова Г.Ф. Ионные каналы возбудимой клетки (структура, функция, патология) [7].
От химического — к электрическому
А теперь — ещё раз самое главное. Мы должны осознанно перейти от движения химических частиц к движению электрических зарядов.
Для этой составной части потенциала покоя есть даже специальное название — концентрационный потенциал [5]. Концентрационный потенциал — это часть потенциала покоя, созданная дефицитом положительных зарядов внутри клетки, образовавшимся за счёт утечки из неё положительных ионов калия.
Ну, а теперь немного физики, химии и математики для любителей точности.
Электрические силы связаны с химическими по уравнению Гольдмана. Его частным случаем является более простое уравнение Нернста, по формуле которого можно рассчитать трансмембранную диффузионную разность потенциалов на основе различной концентрации ионов одного вида по разные стороны мембраны. Так, зная концентрацию ионов калия снаружи и внутри клетки, можно рассчитать калиевый равновесный потенциал EK:

где Ек — равновесный потенциал, R — газовая постоянная, Т — абсолютная температура, F — постоянная Фарадея, К + внеш и K + внутр — концентрации ионов К + снаружи и внутри клетки, соответственно. По формуле видно, что для расчёта потенциала между собой сравниваются концентрации ионов одного вида — K + .
Более точно итоговая величина суммарного диффузионного потенциала, который создаётся утечкой нескольких видов ионов, рассчитывается по формуле Гольдмана-Ходжкина-Катца. В ней учтено, что потенциал покоя зависит от трех факторов: (1) полярности электрического заряда каждого иона; (2) проницаемости мембраны Р для каждого иона; (3) [концентраций соответствующих ионов] внутри (внутр) и снаружи мембраны (внеш). Для мембраны аксона кальмара в покое отношение проводимостей РK : PNa :PCl = 1 : 0,04 : 0,45 [5].

Заключение
Итак, поте нциал покоя состоит из двух частей:
Вот теперь мембранный потенциал покоя окончательно сформирован.
Читайте также: