
Группа несовершенные грибы кратко
Обновлено: 05.07.2024
Дейтеромицеты, пли несовершенные грибы, наряду с аскомицетами и базидиомицетами представляют один из крупнейших классов грибов (в нем около 30% всех известных видов). Этот класс объединяет грибы с септированным мицелием, весь жизненный цикл которых обычно проходит в гаплоидной стадии, без смены ядерных фаз. Они размножаются только бесполым путем — конидиями, а половые (совершенные) стадии у них отсутствуют.
Дейтеромицеты существенно отличаются от других классов грибов, представители которых имеют обычно общих предков. Это заведомо гетерогенная группа, виды которой связаны но происхождению с разными группами из двух классов — аскомицетов и базидиомицетов. Некоторые микологи, подчеркивая это отличие несовершенных грибов, называют их формальным классом.
Хотя в системе несовершенных грибов мы пользуемся теми же таксономическими категориями, как и для других групп грибов, в них вкладывается иной смысл — это искусственные, формальные группы, выделяемые на основе чисто внешнего сходства оргапизмов, а не их родства. Например, род дейтеромицетов не соответствует истинному роду (группе близкородственных видов). Он представляет искусственную группу, объединяющую виды со сходными конидиальными спороношениями, называемую часто, в отличие от родов других организмов, формальным родом.
Мы знаем, что часто один род аскомицетов включает виды с разными конидиальными стадиями. Так, у рода микосферелла известны конидиальные спороношения из родов рамулярия (Ramularia), церкоспора (Cercospora), септория (Septoria) и другие, а у рода нектрия конидиальные спороношения из родов акремониум (Acremonium), фузариум (Fusarium), туберкулярия (Tubercularia), цилиндрокарпон (Суlindrocarpon).
С другой стороны, конидиальные спороношения различных родов аскомицетов, даже относящихся к разным порядкам, часто бывают очень похожи. В результате этого в один род дейтеромицетов нередко входят виды, связанные по происхождению с представителями нетолько разных родов, но и порядков и даже классов высших грибов. Род акремониум, например, связан с совершенными стадиями эмерицеллопсис (порядок эуроциевых), хетомиум (порядок сферейных), нектрия (порядок гипокрейных) и кордицепс (порядок спорыньевых), а конидиальные стадии типа эдоцефалюм (Oedocephalum) известны как у дискомицетов, так и у некоторых базидиомицетов. Связи с представителями разных классов установлены и у некоторых родов аспорогенных дрожжей, например кандида (Candida).
Вегетативное тело дейтеромицетов — хорошо развитый, ветвящийся, гаплоидный мицелий, состоящий- обычно из многоядерных клеток. В мицелии всегда присутствуют септы (перегородки), обычно с простыми порами, как у аскомицетов (рис. 51). Можно предположить, что имеются дейтеромицеты и с септами типа, характерного для базидиомицетов, но они пока не изучены методом электронной микроскопии. У аспорогенных дрожжей мицелия нет, а вегетативное тело представлено почкующимися клетками.

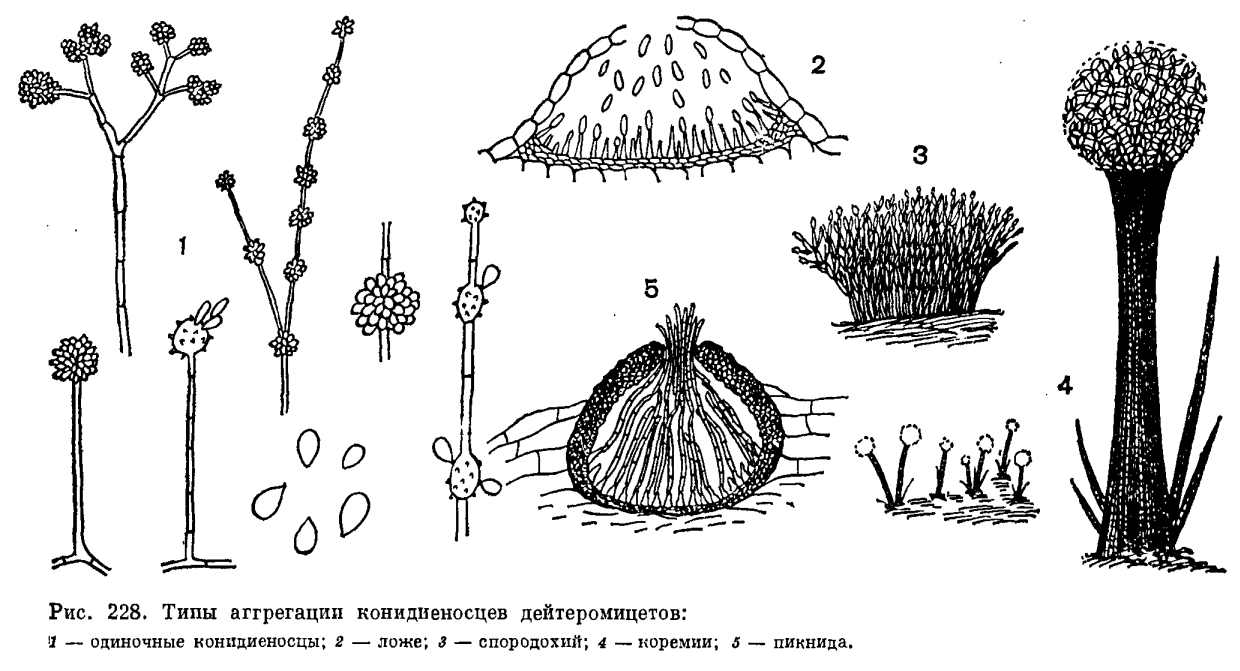
У большинства дейтеромицетов размножение происходит при помощи конидий. Лишь у немногих из них конидиальное спороношение отсутствует. Такие грибы часто образуют склероции (Sclerotium, Rhizoctonia), а иногда встречаются только в виде стерильных мицелиев. Конидии (споры бесполого размножения) образуются на гаплоидном мицелии, на многоклеточных, реже одноклеточных конидиеносцах, представляющих ветви мицелия, обычно поднимающиеся над ним. Они могут быть малодифференцированными, не отличающимися от вегетативных гиф мицелия, но чаще хорошо развиты. Увеличение продукции конидий достигается путем ветвления конидиеносцев различным образом (мутовчатое, моноподиальное, симподиальное, дихотомическое, рис. 228), образования их расширений или вздутий, несущих группы спор и расположенных на вершине конидиеносца (Aspergillus, Oedocephalum, рис. 228) или интеркалярно (Arthrobotrys, Gonatobotrys, рис. 228), а также образованием конидий в длинных акропетальных или базипетальных цепочках (рис. 230).
Многие дейтеромицеты образуют конидиеносцы, объединенные в группы на мицелии или внутри пикнид. Простейшая форма такой аггрегации конидиеносцев — коремии. Многочисленные конидиеносцы развиваются тесно сближенным пучком, обычно склеиваясь своими боками, а иногда срастаясь. В результате этого образуется компактная колонка, на вершине которой на веточках конидиеносцев синхронно развиваются конидии (рис. 228).
Коремии характерны для семейства стильбелловых (Stilbellaceae) порядка гифомицетов, а также для некоторых пенициллов. У многих дейтеромицетов конидиеносцы образуют слой на поверхности выпуклого сплетения гиф или стромы в виде подушечки; Этот тип конидиального спороношения называют спородохиями (рис. 228). Если конидиальные спороношения такого типа имеют слизистую или желеобразную консистенцию, а в основании более рыхлое сплетение гиф мицелия, их называют пиониотами. Конидии, образующиеся в пионнотах, погружены в слизь.
Ложа (ацервулы) по характеру объединения конидиеносцев напоминают спородохии, но конидиеносцы в них образуют тесный слой не на выпуклой строме, а на более или менее плоском сплетении гиф. Ложа часто встречаются у дейтеромицетов — паразитов растений и представляют тесно скученные многочисленные конидиеносцы, сначала развивающиеся в ткани хозяина под кутикулой или эпидермисом, а затем прорывающие их и выступающие наружу (рис. 228).

Конидии дейтеромицетов разнообразны по строению. Они могут быть одноклеточными или с различным числом перегородок, иногда муральными (рис. 229). Обычно они шаровидные или эллипсоидные, но у некоторых дейтеромицетов известны нитевидные, звездчатые или спирально закрученные конидии (рис. 229). У водных гифомицетов они обычно три- или тетрарадиальные, реже сигмоидные (рис. 245). Окраска конидий светлая или темная, буроватокоричневая, вследствие присутствия меланинов.

По характеру развития конидии дейтеромицетов относятся к двум группам — талломной и бластической. Талломные конидии (артроспоры и алевриоспоры) развиваются в результате превращения элемента мицелия. Увеличение в размерах и дифференциация таких конидий происходит после их отделения перегородкой (септой) от конидиогенной клетки; таким образом, конидии этого типа происходят из целой клетки. Артроспоры образуются в результате фрагментации конидиеносца или гифы (Geotrichum, Oidiodendron, рис. 230).
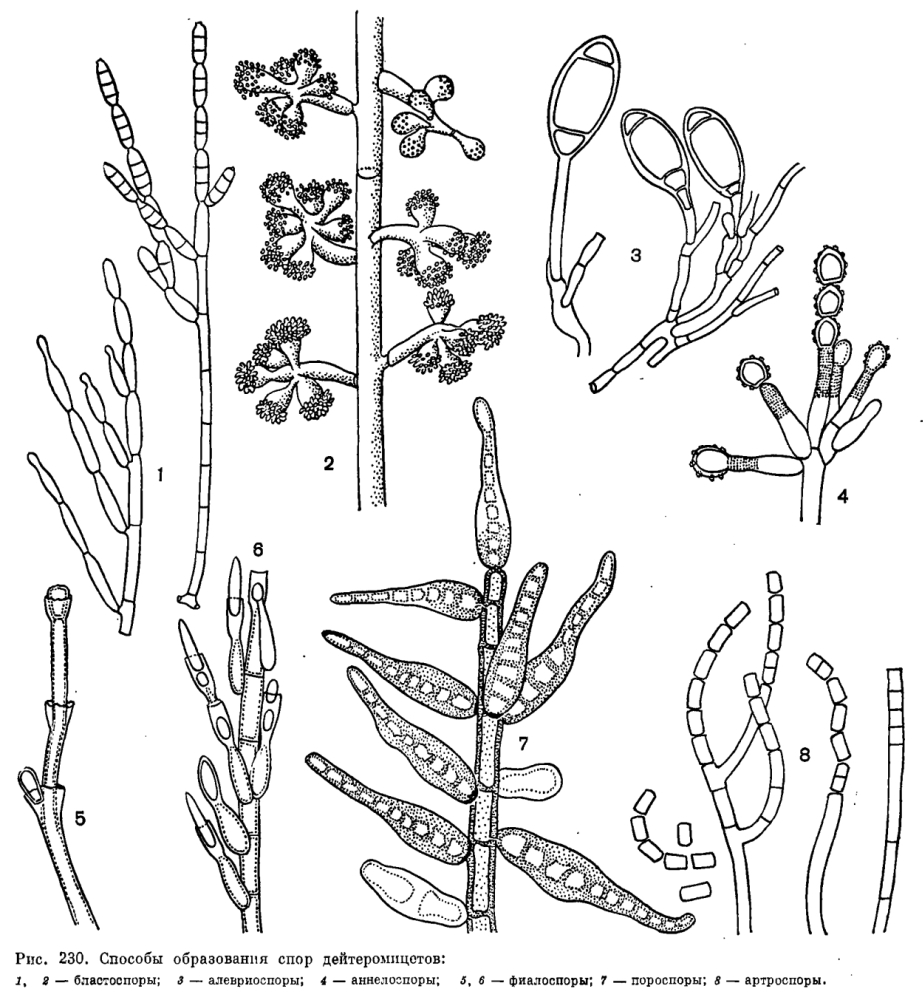
Алевриоспоры образуются из отделившейся перегородкой части конидиогенной клетки, посла разрастания превращающейся в зрелую конидию. Они образуются одиночно на вершине ветвей конидиеносца или гиф и часто имеют крупные размеры и утолщенную клеточную стенку (рис. 230). Иногда они развиваются в коротких цепочках. К этому же типу конидий относятся и аннелоконидии или аннелоспоры, часто образующие довольно длинные цепочки. Первая конидия такой цепочки образуется как концевая алевриоспора, затем конидиеносец прорастает через рубец от споры и конец его превращается в следующую алевриоспору. При многократном прорастании конидиеносца через рубцы спор образуется цепочка алевриоспор. Конидиогенная клетка конидиеносца в результате этого процесса удлиняется, и рубцы, остающиеся после отделения алевриоспор, заметны на ней в виде колец. Конидиогенные клетки такого типа названы аннелофорами, а сами конидии — аннелоконидиями. Они известны у многих грибов, например у аннелофоры африканской (Annelophora africana, или скопуляриопсиса короткостеблевого (Scopulariopsis brevicaulis, рис. 230).
Бластический способ образования конидий характеризуется заметным увеличением в размерах зачатка конидии до его отделения перегородкой от конидиогенной клетки. Конидии здесь развиваются таким образом из части клетки, в отличие от конидий первого типа. На основе участия стенки конидиогенной клет ки в формировании стенки конидии разграничивают три типа бластогенных конидий.
У бластоспор, или бластоконидий, все слои стенки конидиогенной клетки участвуют в образовании степки конидии. Бластоспора развивается как вздутие конца конидиогенной клетки, затем отшнуровывающееся от нее и отделяющееся перегородкой. Этот процесс напомипаот почкование у дрожжей. Бластоспоры образуются в акропетальных цепочках (например, у родов Cladosporium, Septonema, рис. 230) или одиночно, часто на небольших зубчиках (например, у рода Artb.robotrys, рис. 246).
В формировании клеточной стенки пороспор, или пороконидий, принимают участие только внутренние слои клеточной стенки конидиогенной клетки. Они образуются путем почкования через поры в стенках конидиеносцев. Пороспоры обычно толстостенные, развиваются по одной на вершине и по бокам конидиеносца. Конидии этого типа образуются у представителей рода гельминтоспорий (Helminthosporium), а также у некоторых других гифомицетов (рис. 230).
При образовании третьего типа бластогенных конидий — фиалоспор, или фиалоконидий,— клеточная стенка конидии формируется заново, стенка конидиогенной клетки не участвует в ее образовании. Фиалоспоры образуются на фиалидах — конидиогенных клетках, обычно утолщенных у основания и слегка оттянутых в верхней части, у многих грибов с ясно заметным воротничком (например, у родов Phialophora, Catenularia, рис. 230). У других грибов воротничок фиалиды заметен мало или вообще отсутствует, как у пенициллов и аспергиллов (рис. 231).
У несовершенных грибов с разными типами конидий хорошо прослеживается параллелизм в усложнении структуры конидиеносцев и их аггрегации, ведущих к увеличению количества спор. Например, среди грибов с фиалоспорами можно обнаружить виды с простыми конидиеносцами (Acremonium), обильно ветвящимися конидиеносцами (пенициллы), коремиями (Stilbella), спородохиями (Tubercularia), ложами (Colletotrichum, МугоШесшт),.пикнидами (Cytospora).
Параллельный ряд усложняющихся форм конидиального аппарата представляют грибы с алевриоспорами. У грибов с пороспорами и артроспорами известны формы только с одиночными конидиеносцами.
Конидии несовершенных грибов, имеющие сухую поверхность, распространяются обычно воздушными течениями. В воздухе можно обнаружить в больших количествах споры грибов из родов алътернария (Alternaria), кладоспорий (Cladosporium), пенициллов и многих других. Если чашку Петри с питательной средой хотя бы ненадолго оставить открытой, в ней уже через 3—4 дня появляются многочисленные колонии этих грибов. Их можно обнаружить даже на высоте более 7000 м над землей. У представителей несовершенных грибов, споры которых погружены в слизь (Acremonium, Verticillium, Graphium), в распространении их участвует вода (часто капли дождя) или мелкие животные (например, жуки-короеды, переносящие конидии графиума — конидиальной стадии возбудителя голландской болезни вязов). При низкой влажности воздуха и высыхании слизи такие споры переносятся также токами воздуха.
Освобождение конидий у несовершенных грибов обычно происходит пассивно. Активное их освобождение наблюдают очень редко. У грибов из родов дейгтониелла (Deightoniella) и курвулярия (Curvularia) конидиогенная клетка теряет воду в результате испарения, в ней развивается отрицательное давление и стенки клетки искривляются. Если процесс потери воды продолжается, оставшиеся ее молекулы сцепляются между собой и со стенками клетки. Вода в клетке находится поэтому в состоянии повышенного поверхностного натяжения, пока не достигается точка, в которой оно снимается путем перехода воды в газообразную фазу. При этом втянутая клеточная стенка конидиогенной клетки внезапно расправляется и сообщает толчок соседней с ней клетке — конидии, которая отбрасывается в воздух (Д. С. Мередит).
Размножение несовершенных грибов происходит только бесполым путем, поэтому можно было бы предположить, что все особи, происшедшие из конидий, образовавшихся из одной колонии, и последующие их поколения будут развиваться практически в виде чистых линий и изменяться очень мало, только за счет немногочисленных спонтанных мутаций. Но в природе у несовершенных грибов этого никогда не наблюдается. Они представляют, напротив, одну из наиболее вариабильных и подвижных в экологическом отношении групп грибов, именно поэтому весьма широко распространенную в природе и заселяющую самые разнообразные субстраты. Это объясняется тем, что у несовершенных грибов очень часто мицелий гетерокариотичен, т. е. содержит генетически различные ядра.
Гетерокариоз известен в различных таксономических группах грибов — у оомицетов, аскомицетов, зигомицетов, но у несовершенных грибов он представляет собой основной механизм изменчивости. В процессе развития гетерокариотического мицелия число ядер того или иного типа может варьировать в зависимости от изменения условий среды, обеспечивая таким образом адаптацию гриба к этим изменениям. Так, пеницилл распростертый (Penicillium expansum) в природе всегда встречается в гетерокариотическом состоянии. Его гомокариотические изоляты (с генетически однородными ядрами) обычно растут хуже, чем исходный гетерокарионт. Содержание ядер разных генетических типов у этого гриба регулируется условиями среды, например составом питательных сред: при изменении содержания в среде яблочной пульпы число ядер одного из генетических типов (А) колеблется от 13 до 58%. Такие же колебания числа ядер разных типов в зависимости от условий обитания обнаружены и у некоторых фузариумов.
Образование гетерокариотического мицелия у несовершенных грибов может происходить несколькими путями. У одних из них на гетерокариотическом мицелии образуются конидии, содержащие генетически различные ядра. Из таких конидий снова развивается гетерокариотический мицелий. У аспергилла угольного (Aspergillus carbonarius), например, конидии содержат по 2—5 ядер. Гетерокарионты между диким типом с черными конидиями и мутантом со светло-коричневыми конидиями образуют споры промежуточной окраски, которые, в свою очередь, дают колонии также смешанного типа. Если конидии на гетерокариотическом мицелии образуются из одноядерных конидиогенных клеток, они гомокариотичны, т. е. содержат ядра только одного генетического типа. У другого вида аспергиллов — A. versicolor — гетерокарионты между диким штаммом с зелеными конидиями и мутантом с белыми конидиями образуют головки с цепочками белых и зеленых конидий. Из таких конидий вырастает гомокариотический мицелий. Гетерокариотичность у него может восстанавливаться в результате образования анастомозов между гифами с генетически различными ядрами, а также в результате мутаций в отдельных ядрах мицелия.
В некоторых случаях гетерокариоз может быть основой не только изменчивости и адаптации гриба в результате изменения в его мицелии числа генетически различных ядер, но и рекомбинации признаков. Однако рекомбинация у несовершенных грибов происходит не при мейозе, а при митозе в изредка образующихся в гетерокариотическом мицелии диплоидных ядрах. Такие ядра могут гаплоидизироваться в результате утраты ими хромосом.
Процесс рекомбинации такого типа был назван парасексуальным процессом (Дж. Понтекорво). Он слагается из нескольких этапов: слияния ядер гетерокариона и образования диплоидного гетерозиготного ядра; размножения таких гетерозиготных ядер в гетерокариотическом мицелии вместе с гаплоидными; митотической рекомбинации (при митозе) во время размножения диплоидных ядер; вегетативной гаплоидизации диплоидных ядер в результате утраты ими хромосом. Парасексуальный процесс известен сейчас у многих несовершенных грибов, однако его наблюдают у них только в условиях лаборатории; какова же его роль в естественных условиях обитания, пока не установлено.
Несовершенные грибы широко распространены в природе во всех районах земного шара. Многие представители этой группы обитают как сапрофиты в почве, составляя большую часть почвенных грибов. Они в изобилии встречаются на различных растительных субстратах, реже на субстратах животного происхождения. Эти группы несовершенных грибов принимают активное участие в разложении органических остатков и в почвообразовательном процессе. Некоторые сапрофитные несовершенные грибы вызывают плесневение пищевых продуктов и различных промышленных изделий.
Многочисленна группа несовершенных грибов, паразитирующих на высших растениях. Они вызывают многочисленные серьезные болезни сельскохозяйственных культур, приносящие большой экономический ущерб. Некоторые представители этого класса вызывают заболевания у животных и человека. Развиваясь на зерне и других продуктах питания, отдельные несовершенные грибы выделяют в них токсины, которые могут вызвать тяжелые отравления при использовании таких продуктов в пищу человеком или кормлении ими животных.
Среди несовершенных грибов известны многочисленные продуценты биологически активных веществ, используемые при производстве антибиотиков (пенициллина, гризеофульвина, фумагиллина, трихотецина), различных ферментов и органических кислот. Несовершенные грибы, паразитирующие на насекомых-вредителях и грибах, патогенных для растений, а также хищные грибы, уничтожающие фитонематод, используют для разработки биологических методов защиты растений от вредителей и болезней.
Широко используемая сейчас всеми микологами система несовершенных грибов, разработанная П. А. Саккардо в 1880 г., заведомо искусственна и служит только более или менее удобной схемой для их упорядочения и определения. Несовершенные грибы подразделяются в ней на три порядка на основе строения конидиального аппарата. Порядок гифомицеты (Hyphomycetales) объединяет несовершенные грибы, образующие одиночные конидиеносцы или конидиеносцы, соединенные в коремии и спородохий. К порядку меланкониевых (Melanconiales) относятся несовершенные грибы, образующие ложа, а к порядку сферопсидных (Sphaeropsidales) — образующие пикниды.
Некоторые микологи (С. Дж. Xьюз, К. В. Субраманиан, К. Тубаки, С. Нильссон, Дж. Л. Баррон и другие) делят несовершенные грибы на разное число групп по типу спорогенеза, основными из которых являются: бластоспоровые (Blastosporae), алевриоспоровые (Aleuriosporae), фиалоспоровые (Phialosporae), пороспоровые (Рогоsporae) и артроспоровые (Arthrosporae).
Однако новая система несовершенных грибов находится пока в процессе разработки, она охватывает далеко не все известные сейчас несовершенные грибы. Морфогенез конидий хорошо изучен только у гифомицетов, но он известен лишь у отдельных представителей двух других порядков.
В настоящей книге мы пользуемся системой П. А. Саккардо, охватывающей все многообразие несовершенных грибов.
Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров . 1974 .

Тип Анаморфные грибы (Anamorphic fungi) – таксон объединяет виды грибов с вегетативным телом в виде септированного мицелия или отдельных клеток (дрожжи), размножающиеся бесполым путем (бесполое размножение), делением или почкованием. До недавнего времени Анаморфные грибы или Дейтеромицеты позиционировали как группу грибов, утратившую в ходе эволюции половую стадию (телеоморфу). В настоящее время под этим названием объединяют все анаморфы высших грибов, и виды у которых телеоморфная стадия отсутствует или неустановленна [4] [1] .
Фактически Анаморфные грибы или Дейтеромицеты являются полифелитической группой грибов, то есть объединяют различные подгруппы, в отношении которых доказано близкое родство с другими группами грибов, не входящими в нее. Объединение Анаморфные грибы основано на поверхностном сходстве (септированный мицелий, конидии, конидиеносцы) и не включает наиболее близкого общего предка для видов грибов, входящих в данный таксон. Есть мнение, что тип Анаморфные грибы, не имеет права на существование в современной систематике и при более глубоких исследованиях, входящих в него видов, будет расформирован. Однако, на сегодняшний день, такое действие не считается целесообразным [4] [1] .
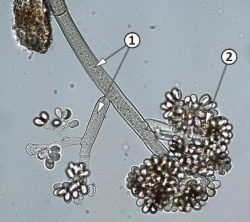
Конидиеносцы и конидии Botrytis cinerea

Систематика несовершенных грибов
Существует несколько различных подходов к систематике Анаморфных грибов (Дейтеромицетов):
На сегодняшний день реально пользуются, особенно на практике, системами, построенными на внешних признаках. Параллельно не забывают об их формальности или искусственности. В связи с этим группы называют: формальный класс, формальный порядок, а само объединение Анаморфные грибы (Anamorphic fungi) рассматривается, как формальный тип [1] .
Таким образом, чаще всего на практике используется разделение Анаморфных грибов на три формальные класса по строению конидиального спороношения:
- Класс Гифомицеты(Hyphomycetes) – включает виды, характеризующиеся простыми или ветвящимися конидиеносцами, развивающимися на мицелии одиночно, в коремиях или спородохиях;
- Класс Целомицеты(Coelomycetes) – включает виды с конидиеносцами, собранными в группы.
- Класс Агономицеты(Agonomycetes) – включает виды, не образующие спороношения. В цикле развития данных грибов установлены только мицелий и склероции[1] .
Особенности номенклатуры
Внедрение в фитопатологию современных методов исследования привело к тому, что у многих анаморфных видов обнаруживаются родственные связи с уже известными телеоморфами, принадлежащими к типу Аскомицеты (Ascomycota) или типу Базидиомикота (Basidiomycota). В некоторых случаях такие виды рекомендуют относить к типу Аскомицеты. В частности, виды рода Penicillium и рода Aspergillus [2] .
В большинстве случаев Международный кодекс ботанической номенклатуры разрешает использовать устоявшееся название по анаморфе наряду с основным названием гриба по его телеоморфе. Самостоятельные видовые названия конидиальной стадии допускаются и для представителей типа Базидиомикота (Basidiomycota). Кроме того, во многих источниках конидиальные стадии (анаморфы) Аскомицетов (Ascomycota) и Базидиомикот (Basidiomycota) относят к типу Анаморфные грибы [2] .
Исключение представляют виды Класса Эуаскомицеты (Euascomycetidae), имеющие в цикле развития анаморфную стадию и телеоморфную стадию. Их анаморфы часто не имеют собственного названия и относятся к типу Аскомицеты (Ascomycota) [3] .
В частности, вид Blumeria graminis, вызывающий мучнистую росу на зерновых культурах, имеет в цикле развития как телеоморфу, так и анаморфу. По систематическому положению обои стадии относятся к типу Аскомицеты (Ascomycota), класс Эуаскомицеты (Euascomycetes), порядок Эризифовые (Мучнисторосяные) (Erysiphales), семейство Эризифовые (Erysiphaceae), род Erysiphe [3] .

Царство грибов отличается поразительным многообразием. Всем известны съедобные его представители – настоящее украшение стола. Многие знают о существовании ядовитых грибов, которые нередко так удачно маскируются, что отличить их бывает непросто. Наконец, на слуху простые по строению организмы, например мукора, то есть плесень. Но что такое несовершенные грибы, за что получили свое нелестное название? Наша статья поможет разобраться в особенностях их строения, классификации и значении.
Определение
В царстве грибов выделяется особый класс несовершенных грибов, именующиеся дейтеромицетами, к которому относятся высшие организмы. Для них присущ лишь один вид размножения – бесполое спороношение. К таковым относятся те организмы, что не имеют возможности образовывать базидий либо сумку. Класс довольно обширен, включает более 25 тысяч видов. При этом отдельные представители обладают даже половым спороношением, но оно не играет особой роли в их развитии.

Значение в живой природе подобных организмов довольно специфично – они являются возбудителями различных заболеваний растений.
Отличительные особенности
Группа несовершенных грибов имеет ряд признаков, по которым их можно отличить от иных представителей царства. Особенности такие:
- Размножение происходит особыми спорами – конидиями, которые формируются на участках мицелия, отличающихся обособленностью. Эти ответвления принято называть конидиеносцами.
- Размножение может походить путем рассечения гиф на отдельные клетки.
- Половые процессы у таких грибов, как правило, отсутствуют.
Исследования отдела несовершенных грибов доказали, что многие представители являются одной из стадий развития сумчатых либо базидиальных грибов.
Чаще всего ведут паразитический образ жизни, поражая растения, реже – животных. Разнообразие представителей несовершенных грибов, а также недостаток материала для исследования привели к путанице. Так, в науке прошлого было принято дейтеромицеты относить к отделу царства Грибы, однако теперь большинство ученых поддерживает позицию, что это не таксономическая группа.

Специфика строения
Примеры несовершенных грибов позволяют выявить особенности, присущие строению группы:
- Тело составляют многоклеточные гифы, расчлененные. Они могут быть окрашенными или прозрачными.
- Споры для размножения – конидии – располагаются на конидиеносцах или пикнидах.
- Мицелий чаще всего многоядерный, с большим количеством септ (перегородок).
- Конидии отличаются богатством форм: чаще всего они представляют собой шар или эллипс, но встречаются и в форме нити, спирали, даже звездочки. Цвет таких спор светлый, бывает коричневатый, темный.
- Освобождение от конидий – сам процесс их попадания в воздух – носит пассивный характер.
Конидиеносцы могут быть одиночными (такое наблюдается у простых по строению организмов), но чаще всего собраны в группы, пучки. В зависимости от их расположения выделяют несколько типов конидиального аппарата (ложе – обилие параллельных, не соединенных друг с другом отростков, коремия – сросшиеся пучки конидиеносцев).
Самый сложный тип конидиального спороношения – пикнида, находящиеся в ней конидии расположены в слизи и на поверхность могут выйти лишь при набухании.
Многообразие
К несовершенным относится большое число грибов:
- Те, которым присуще конидиальное бесполое спороношение.
- Размножающиеся вегетативно, то есть не имеющие спороношения (стерильный мицелий).
- Дрожжевые организмы, для которых присуще размножение почкованием.
- Даже такие организмы, которые имеют возможность совершенного размножения, но использующие ее редко по причине малой значимости в процессе развития.
В природе примеров несовершенных грибов существует огромное количество. Это могут быть паразиты (таковых большинство) или сапрофиты, многие особи представляют собой возбудителей серьезных заболеваний растительных культур. Ниже будет представлена их классификация, краткая характеристика и конкретные примеры.

Классификация
Несовершенные грибы можно подразделить на три порядка:
- Пикнидиальные (сферопсидные).
- Гифальные (гифомицеты).
- Меланкониевые.
Для каждого порядка характерны определенные черты строения.
Характеристика пикнидиальных
Эти фитопатогенные грибы весьма разнообразны и могут вызывать такие поражения растений, как:
- гниль овощей, плодов, семян;
- пятнистость листьев;
- рак ветвей, стволов;
- некрозы.
Узнать о наличии возбудителя можно по появлению большого количества пикнид – черных точек и бугорков на частях растения. Опасны эти несовершенные грибы для свеклы, капусты, лимонов, моркови, картофеля, тыкв, кукурузы, томатов. Огромные вред приносят возбудители черного рака яблони.

Коротко о гифомицетах
Среди несовершенных грибов более всего распространены гифальные, которые весьма разнличные по форме и строению. Мицелий таких организмов отличается хорошим развитием, обычно имеет перегородки. Может располагаться в субстрате или находиться на его поверхности. Конидии появляются как результат рассечения мицелия, могут также формироваться на конидиеносцах.
Они могут вызывать такие заболевания культур:
- гниение;
- плесень;
- увядание;
- образование пятен.
Так, именно такие несовершенные грибы являются возбудителями следующих заболеваний:
- фузариозное увядание льна;
- сухая гниль картофеля (клубней); семечковых;
- серая гниль отдельных овощей, плодов и ягод;
- зеленая и голубая плесень плодов цитрусов;
- ооспороз (парша картофеля, гниль томатов).
Нередко именно эта группа организмов повреждает корма, предназначенные для длительного хранения.

Однако не стоит полагать, что несовершенные грибы приносят лишь вред, среди них имеется несколько полезных видов. Использование их многообразно:
- Изготовление антибиотиков, органических кислот.
- Применение в борьбе с насекомыми-вредителями. применяются для уничтожения фитогельминтов.
Как видим, данный порядок и его представители способны приносить человечеству, как пользу, так и вред. Среди этих организмов многие являются сапрофитами.
Информация о меланкониевых грибах
К порядку Меланкониевые относится небольшое число несовершенных грибов, сходных между собой по особенностям строение и развития. Отличительная особенность представителей – конидии расположены в пикнидиях. У растений такие грибы вызывают заболевания, относимые к антракнозу – на плодах и семенах культур образуются небольшие по размеру язвы, стебли растрескиваются, на листьях появляется пятнистость.
Приведем примеры опасных заболеваний, которые наносят урон сельскохозяйственной деятельности человека:
- антракноз тыквенных;
- поражение фасоли;
- льна;
- винограда;
- смородины.
- бурая пятнистость земляничных листьев.
Отдельные виды обладают очень высокой вредоносностью.

Значение в природе
Мы рассмотрели, что представляют собой несовершенные грибы. А какова их роль в природе? Эти живые организмы помогают уничтожить остатки погибших растений, то есть осуществляют функции редуцентов.
Однако подавляющая часть таких грибов вредна: они не только паразитируют на культурных растениях, но и могут оставлять токсичные вещества в продуктах питания, которые становятся средой их жизни. Человек, употребив такой продукт, может отравиться.
Итак, несовершенные грибы – весьма условное понятие, ведь к ним относится огромное количество видов, отличающихся бесполым способом размножения. В группу включено большое число представителей, часть из которых является паразитами и наносит огромный ущерб сельскому хозяйству, уничтожая зерновые, овощные, плодовые культуры и корма для скота, предназначенные для хранения. Многие из грибов человек научился применять во благо, используя их для производства ферментов, антибиотиков и ядов для защиты урожая от вредоносных насекомых.
Из всего многообразия известных науке грибов в особняком в царстве Грибы стоит класс Несовершенные грибы или Дейтеромицеты. Это класс высших организмов, размножение которых осуществляется только при помощи бесполого спороношения. Некоторые ученые-микологи считают их формальным классом грибов.
Описание и общая характеристика
Дейтеромицеты известны также под названием Митоспоровые или Аноморфные. Этот биологический класс, относящийся к отделу плесневых грибов, насчитывает более 25 тыс. видов Он включает представителей, главной особенностью которых является неспособность образовывать сумки или базидии при размножении, в отличие от совершенных. Они размножаются с помощью пропагул, которые представляют собой конидии.
Реже эти грибы образуются из недифференцированного мицелия. Многие из них – фитопатогенные (провоцируют заболевания растений). Причем у отдельных имеются примеры полового спороношения, но ведущей является конидиальная стадия, а сумчатая приобретает приоритетное значение в развитии. Некоторые из этого класса находятся на промежуточной стадии развития аскомицетовых и базидиомицетовых грибов.
Вегетативное тело митоспоровых представляют собой разветвленный мицелий гаплоидного типа, имеющий перегородки – септы, с простыми порами. Состоит он из многоядерных клеток, образующих гифы (грибные нити). Сам процесс размножения происходит в изолированных групповых конидиеносцах либо на пикнидах – специфических образованиях плодовых тел.
Классификация конидиеносцев подразумевает две разные формы: одиночные и групповые.
Лучшим способом размножения считают пикниды – биологические контейнеры, внутри которых формируются конидии. Вверху у них имеются маленькие отверстия, с помощью которых споры попадают наружу.
Группы дейтеромицетов и их распространение
Несовершенные грибы имеют следующую классификацию по типу конидиального спороношения:
- Гифомицетовые, или гифомицеты: самый массовый порядок, относящийся к этому отделу. Включает много групп, родов и видов, которые непохожи по форме, строению и цвету конидиеносцев и конидий. Главной общей характеристикой этого порядка является то, что конидиеносцы имеют мицелиальный тип развития и на субстрате определяются одиночно или с наличием коремий и спородохий. По этой причине их спороношение выглядит как порошащийся налет. К указанному порядку относится много представителей фитопатогенных видов, вредящих сельскохозяйственным насаждениям. Болезни, вызванные ими, приводят к плесени, гнили, увяданию и гибели. Так, гриб рода Фузариум служит причиной сухой гнили на клубнях картофеля, серой гнили на овощных и плодово-ягодных растениях, поражения пятнами свекольной ботвы, побегов ячменя, плесени хлопковых коробочек, плодов цитрусовых, семян и луковиц сельхозрастений. Деление порядка на семейства основано на характере агрегации конидиеносцев, а также окраске их и конидий.
- Меланкониальные: однородная группа, включающая небольшое количество представителей, которые относятся к классу Несовершенные грибы. Их конидии образуются в специальных ложах укороченных конидиеносцев. Они проявляются на поверхности пораженной питательной среды в виде подушечек, плоской либо выступающей формы. Провоцируют язвы на поверхности семян, трещины стебля и пятна на листьях. Из-за наличия слизи распространение конидий при помощи воды, животных и насекомых, особенно тех, которые используют в пищу эту слизь.
- Сферопсидные, или пикнидиальные: отличаются тем, что их конидии, сформированные в пикнидах, могут погружаться в субстрат. У многих представителей этого порядка споровместилища (пикниды) в виде мелких, едва заметных невооруженным глазом шариков – откуда и происходит название порядка. Вызывают много заболеваний сельскохозяйственных культур, проявляющихся гнилями, пятнистостями, некрозными поражениями частей стебля и цитоспорозом. Общий признак – появление пикнид на пораженных участках в виде выпуклостей и черных точек.
Также к классу Дейтеромицет относятся микофильные и энтомопатогенные несовершенные грибы
Значение в жизни человека

Плесневые грибы применяются для изготовления антибиотиков
Несовершенные грибы встречаются в почве, растительных остатках либо в субстратах животного происхождения. Хотя большинство их представителей являются обычными сапрофитами, они способны вызывать болезни и паразитировать на животных и даже людях, доставляя немало хлопот.
Пикнидиальные грибы часто паразитируют на лишайниках, водорослях и вызывают заболевания культурных растений. Но другой немаловажной их особенностью является способность повреждать промышленные неорганические материалы: лакокрасочные изделия, штукатурку, бетон, поселяться в пресноводных и морских водоемах. Существуют патогенные и токсичные виды, провоцирующие заболевания кожного покрова (например, стригущий лишай), а также способные привести к тяжёлой интоксикации организма, развиваясь на пищевых продуктах.
Однако несовершенные грибы имеют и полезные свойства. Так, аспергиллы играют важную роль в производстве органических кислот: лимонной, щавелевой и прочих.
Энтомопатогенные несовершенные грибы имеют большое значение в ограничении численности вредителей цитрусовых.
Их используют в целях производства ферментов для пищевой отрасли и текстильной промышленности, а также для борьбы с насекомыми-вредителями (биологический метод). Они играют важную роль в почвообразовательном процессе и переработке органических остатков. Также применяют для изготовления антибиотиков.
Заключение
Несовершенные грибы, в большинстве случаев, ухудшают качество продукции, но человек научился часть их использовать себе во благо. Исследования в этой области не прекращаются и позволяют открывать все новые сферы применения и узнавать новое об этих грибах.
Читайте также: