
Бикарбонатная буферная система кратко
Обновлено: 07.07.2024
Бикарбонатная буферная система — буферная система, состоящая из угольной кислоты (H 2 CO 3) в качестве донора протонов (H +) и бикарбоната (HCO-3) в качестве их акцептора. Бикарбонатная буферная система играет роль в поддержании постоянства pH жидкостей организма животных, в частности, она является важнейшей буферной системой крови, тканевой жидкости и лимфы, также нужна для забуференном внутриклеточной жидкости.
Механизм действия
Бикарбонатная система несколько сложнее других буферные системы, состоящие из пары слабая кислота / сопряженная основа, поскольку один из ее компонентов — угольная кислота — образуется в оборотной реакции с выделением углекислого газа, растворенного в плазме крови. А концентрация последнего в свою очередь зависит от парциального давления CO 2 в газовой фазе, с которой кровь контактирует, то есть в альвеолярном воздухе. Таким образом, pH бикарбонатной буферной системы зависит от трех равновесных процессов:
Когда концентрация ионов H + в крови увеличивается, например, вследствие выделения молочной кислоты мышцами, равновесие уравнения 1 смещается в сторону образования угольной кислоты, это в свою очередь приводит к повышению содержания растворенного углекислого газа (уравнение 2) и, наконец, к увеличению парциального давления CO 2 в легких (уравнение 3). Лишний оксид углерода выдыхаемого воздуха. Когда pH крови повышается, например, вследствие образования NH 3 при катаболизма белков, происходит обратный процесс: больше угольной кислоты распадается до бикарбоната, и, соответственно, больше углекислого газа растворяется в плазме.
Частота и глубина дыхания регулируется центром в стволе головного мозга, который получает информацию о концентрации углекислого газа в крови и ее кислотность. Снижение pH и увеличения парциального давления p (CO 2) стимулирует ускорение газообмена в легких.
Вычисление pH
Зона буферирования для пары слабая кислота / сопряженная основа, то есть диапазон pH, в котором такая буферная система может эффективно работать, рассчитывается как pK a ± 1. Негативный логарифм константы кислотной диссоциации для угольной кислоты pK a = 3,57 при 37 ° C. Итак, при физиологических условиях концентрация H 2 CO 3 (примерно 1 мм) существенно ниже, чем концентрация HCO-3 (24-25 мм), поэтому такая система должна была быть очень эффективной в предотвращении снижению pH, но при выделении в кровь щелочных веществ ее буферная емкость должна быстро исчерпываться. Однако, поскольку кровь постоянно контактирует с большим резервной емкостью углекислого газа в воздухе легких, бикарбонатная система может эффективно противостоять и увеличению pH. Реальное наблюдаемое значение pK a в физиологических условиях для нее составляет 6,1. В клинической медицине для вычисления pH плазмы, исходя из концентрации растворенного углекислого газа, используют такую модификацию уравнения Гендерсона-Гассельбалха:
где p (CO 2) выражается в килопаскалях (обычно от 4,6 до 6,7 кПа), а коэффициент 0,23 отражает растворимость углекислого газа в воде.
В культуральных средах
Бикарбонатная буферная система широко используется в средах для культивирования эукариотических клеток, например DMEM и RPMI-1640. В отличие от органических буферных агентов, таких как MOPS и HEPES, бикарбонат не является токсичным и большинство клеток потребует его для роста, не в зависимости от его роли в поддержании устойчивого pH.
Кроме самого бикарбоната, который добавляется в среды в виде натриевой соли, бикарбонатная буферная система для эффективного функционирования требует достаточной концентрации углекислого газа. Хотя он и выделяется живыми клетками, этого количества недостаточно, чтобы поддерживать необходимое значение pH. Из-за этого клетки культивируют в CO 2 -инкубаторах с концентрацией углекислого газа 5-10%. Если среда, в которой используется бикарбонатная буферная система, длительное время контактирует с обычным воздухом (концентрация CO 2 0,04%) его pH постепенно растет.
Буферные системы – это соединения, противодействующие резким изменениям концентрации ионов Н + . Любая буферная система - это кислотно-основная пара: слабое основание (анион, А – ) и слабая кислота (Н-Анион, H-А). Они минимизируют сдвиги количества ионов Н + в растворе за счет их связывания с анионом и включения в плохо диссоциирующее соединение – в слабую кислоту. Поэтому общее количество ионов Н + изменяется не так заметно, как это могло бы быть.
Существует три буферные системы жидкостей организма – бикарбонатная, фосфатная, белковая (включая гемоглобиновую).Они вступают в действие моментально и через несколько минут их эффект достигает максимума возможного.
Фосфатная буферная система
Фосфатная буферная система составляет около 2% от всей буферной емкости крови и до 50% буферной емкости мочи. Она образована гидрофосфатом (HPO4 2– ) и дигидрофосфатом (H2PO4 – ). Дигидрофосфат слабо диссоциирует и ведет себя как слабая кислота, гидрофосфат обладает щелочными свойствами. В норме отношение HРO4 2– к H2РO4 – равно 4 : 1.
При взаимодействии кислот (ионов Н + ) с двузамещенным фосфатом (HPO4 2‑ ) образуется дигидрофосфат (H2PO4 – ):
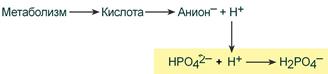
Удаление ионов H + фосфатным буфером
В результате концентрация ионов Н + понижается.
При поступлении в кровь оснований (избыток ОН – ‑групп) они нейтрализуются поступающими в плазму от H2PO4 – ионами Н + :
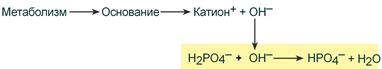
Удаление щелочных эквивалентов фосфатным буфером
Роль фосфатного буфера особенно высока во внутриклеточном пространстве и в просвете почечных канальцев. Кислотно-основная реакция мочи зависит только от содержания дигидрофосфата (H 2 PO 4 – ), т.к. бикарбонат натрия в почечных канальцах реабсорбируется.
Бикарбонатная буферная система
Эта система самая мощная, на ее долю приходится 65% всей буферной мощности крови. Она состоит из бикарбонат-иона (НСО3 – ) и угольной кислоты (Н2СО3). В норме отношение HCO3 – к H2CO3 равно 20 : 1.
При поступлении в кровь ионов H + (т.е. кислоты) ионы бикарбоната натрия взаимодействуют с ней и образуется угольная кислота:
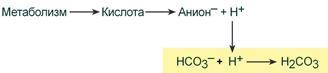
При работе бикарбонатной системы концентрация водородных ионов понижается, т.к. угольная кислота является очень слабой кислотой и плохо диссоциирует. При этом в крови не происходит параллельного значимого увеличения концентрации НСО3 – .
Если в кровь поступают вещества с щелочными свойствами, то они реагируют с угольной кислотой и образуют ионы бикарбоната:
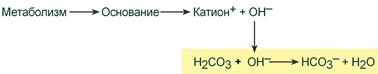
Работа бикарбонатного буфера неразрывно связана с дыхательной системой (с вентиляцией легких). В легочных артериолах при снижении плазменной концентрации СО2 и благодаря присутствию в эритроцитах фермента карбоангидразы угольная кислота быстро расщепляется с образованием CO2, удаляемого с выдыхаемым воздухом:
Кроме эритроцитов, значительная активность карбоангидразы отмечена в эпителии почечных канальцев, клетках слизистой оболочки желудка, коре надпочечников и клетках печени, в незначительных количествах – в центральной нервной системе, поджелудочной железе и других органах.
Белковая буферная система
Белки плазмы, в первую очередь альбумин, играют роль буфера благодаря своим амфотерным свойствам. Их вклад в буферизацию плазмы крови около 5%.
В кислой среде подавляется диссоциация СООН‑групп аминокислотных радикалов (в аспарагиновой и глутаминовой кислотах), а группы NH2 (в аргинине и лизине) связывают избыток Н + . При этом белок заряжается положительно.
В щелочной среде усиливается диссоциация COOH‑групп, поступающие в плазму ионы Н + связывают избыток ОН – ‑ионов и pH сохраняется. Белки в данном случае выступают как кислоты и заряжаются отрицательно.
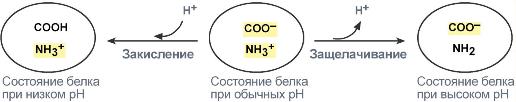
Изменение заряда буферных групп белка при различных рН
Гемоглобиновая буферная система
Высокой мощностью в крови обладает гемоглобиновый буфер, на него приходится до 28% всей буферной емкости крови. В качестве кислой части буфера выступает оксигенированный гемоглобин H‑HbO2 . Он имеет выраженные кислотные свойства и в 80 раз легче отдает ионы водорода, чем восстановленный Н‑Нb , выступающий как основание. Гемоглобиновый буфер можно рассматривать как часть белкового, но его особенностью является работа в теснейшем контакте с бикарбонатной системой.
Изменение кислотности гемоглобина происходит в тканях и в легких, и вызывается связыванием соответственно H + или О2. Непосредственный механизм действия буфера заключается в присоединении или отдаче иона H + остатком гистидина в глобиновой части молекулы (эффект Бора).
В тканях более кислый pH в норме является результатом накопления минеральных (угольной, серной, соляной) и органических кислот (молочной). При компенсации pH данным буфером ионы H + присоединяются к пришедшему оксигемоглобину (HbО2) и превращают его в H‑HbО2. Это моментально вызывает отдачу оксигемоглобином кислорода (эффект Бора) и он превращается в восстановленный H‑Hb.
В результате снижается количество кислот, в первую очередь Н2СО3, продуцируются ионы НСО3 ‑ и тканевое пространство подщелачивается.
В легких после удаления СО2 (угольной кислоты) происходит защелачивание крови. При этом присоединение О2 к дезоксигемоглобину H-Hb образует кислоту более сильную, чем угольная. Она отдает свои ионы Н + в среду, предотвращая повышение рН:
Работу гемоглобинового буфера рассматривают неотрывно от бикарбонатного буфера:
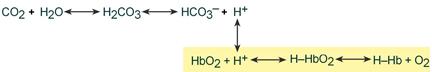
Эффективность гемоглобинового буфера напрямую зависит от активности дыхательной системы (Газообмен в легких и тканях).
Эта система состоит из бикарбонат-иона (НСО3–) и угольной кислоты (Н2СО3), буферная мощность составляет 65% от общей буферной емкости крови.
В норме отношение HCO3– к H2CO3 равно 20 : 1.
Работа этой системы неразрывно и тесно связана с легкими.
При поступлении в кровь более сильной кислоты, чем угольная, ионы бикарбоната натрия взаимодействуют с ней, происходит реакция обмена и образуется соответствующая соль и угольная кислота.
В результате, благодаря связыванию введенной в систему кислоты, концентрация ионов водорода значительно понижается.
NaНСО3 + Н-Анион > H2CO3 + Na+ + Анион–
При поступлении оснований они реагируют с угольной кислотой и образуют соли бикарбонатов:
H2CO3 + Катион-ОН > Катион+ + HCO3– + Н2О
Возникающий при этом дефицит угольной кислоты компенсируется уменьшением выделения CO2 легкими (и в этот момент мы сами не замечаем как задерживаем дыхание на какое-то время.А еще лучше в этом плане работает регулярная практика задержки дыхания впранаяме- кумбхака. ALG)
При накоплении угольной кислоты в крови не происходит параллельного значимого увеличения концентрации H2CO3-, т.к. угольная кислота очень плохо диссоциирует (к вопросу о безопасности практики кумбхаки в параняме. Как только дыхание восстанавливается, излишек угольной кислоты тут же удаляется. ALG)
Благодаря работе бикарбонатного буфера концентрация водородных ионов понижается по двум причинам:
• угольная кислота является очень слабой кислотой и плохо диссоциирует
• в крови легких благодаря присутствию в эритроцитах фермента карбоангидразы, угольная кислота быстро расщепляется с образованием CO2, удаляемого с выдыхаемым воздухом:
(и тоже, получается, сто очков в плюс к дыхательной практике, ведь если легкие работают не на полный объем - как это у нас столь распространено - СО2 выделяется не в полном объеме, и эффективность работы этого важнейшего буфера существенно снижается. ALG)
Кроме эритроцитов, значительная активность карбоангидразы отмечена в эпителии почечных канальцев, клетках слизистой оболочки желудка, коре надпочечников и клетках печени, в незначительных количествах – в центральной нервной системе, поджелудочной железе и других органах.
Фосфатная буферная система
Фосфатная буферная система составляет около 1-2% от всей буферной емкости крови и до 50% буферной емкости мочи.
Она образована дигидрофосфатом (NaH2PO4) и гидрофосфатом (Na2HPO4) натрия.
Первое соединение слабо диссоциирует и ведет себя как слабая кислота, второе обладает щелочными свойствами.
В норме отношение HРO42– к H2РO4– равно 4 : 1.
При взаимодействии кислот (ионов водорода) с двузамещенным фосфатом натрия (Na2HPO4) натрий вытесняется, образуется натриевая соль дигидрофосфата (H2PO4–). В результате, благодаря связыванию введенной в систему кислоты, концентрация ионов водорода значительно понижается.
HPO42– + Н-Анион > H2PO4– + Анион–
При поступлении оснований избыток ОН– групп нейтрализуется имеющимися в среде Н+, а расход ионов Н+ восполняется повышением диссоциации NaH2PO4.
H2PO4– + Катион-ОН > Катион+ + HPO42– + Н2О
Основное значение фосфатный буфер имеет для регуляции pH интерстициальной жидкости и мочи.
В моче роль его состоит в сбережении бикарбоната натрия за счет дополнительного иона водорода (по сравнению с NaHCO3) в составе выводимого NaH2PO4:
Na2HPO4 + Н2СО3 > NaH2PO4 + NaНСО3
Кислотно-основная реакция мочи зависит только от содержания дигидрофосфата, т.к. бикарбонат натрия в почечных канальцах реабсорбируется.
Белковая буферная система
Буферная мощность этой системы составляет 5% от общей буферной емкости крови.
Белки плазмы, в первую очередь альбумин, играют роль буфера благодаря своим амфотерным свойствам.
В кислой среде подавляется диссоциация СООН-групп, а группы NH2 связывают избыток Н+, при этом белок заряжается положительно.
В щелочной среде усиливается диссоциация карбоксильных групп, образующиеся Н+ связывают избыток ОН–-ионов и pH сохраняется, белки выступают как кислоты и заряжаются отрицательно.
Гемоглобиновая буферная система
Наибольшей мощностью обладает гемоглобиновый буфер, который можно рассматривать как часть белкового. На него приходится до 30% всей буферной емкости крови.
В буферной системе гемоглобина существенную роль играет гистидин, который содержится в белке в большом количестве.
Изоэлектрическая точка гистидина равна 7,6, что позволяет гемоглобину легко принимать и легко отдавать ионы водорода при малейших сдвигах физиологической рН крови (в норме 7,35-7,45).
Данный буфер представлен несколькими подсистемами:

Пара ННb/ННbО2 является основной в работе гемоглобинового буфера.
Соединение ННbО2 является более сильной кислотой по сравнению с угольной кислотой, HHb - более слабая кислота, чем угольная. Установлено, что ННbО2 в 80 раз легче отдает ионы водорода, чем ННb.
Присоединение ионов водорода к остатку гистидина дезоксигемоглобина выглядит так:
Работа гемоглобинового буфера неразрывно связана с дыхательной системой (к вопросу о значении пранаямы! - ALG)
Эта система состоит из бикарбонат-иона (НСО3–) и угольной кислоты (Н2СО3), буферная мощность составляет 65% от общей буферной емкости крови.
В норме отношение HCO3– к H2CO3 равно 20 : 1.
Работа этой системы неразрывно и тесно связана с легкими.
При поступлении в кровь более сильной кислоты, чем угольная, ионы бикарбоната натрия взаимодействуют с ней, происходит реакция обмена и образуется соответствующая соль и угольная кислота.
В результате, благодаря связыванию введенной в систему кислоты, концентрация ионов водорода значительно понижается.
NaНСО3 + Н-Анион > H2CO3 + Na+ + Анион–
При поступлении оснований они реагируют с угольной кислотой и образуют соли бикарбонатов:
H2CO3 + Катион-ОН > Катион+ + HCO3– + Н2О
Возникающий при этом дефицит угольной кислоты компенсируется уменьшением выделения CO2 легкими (и в этот момент мы сами не замечаем как задерживаем дыхание на какое-то время.А еще лучше в этом плане работает регулярная практика задержки дыхания впранаяме- кумбхака. ALG)
При накоплении угольной кислоты в крови не происходит параллельного значимого увеличения концентрации H2CO3-, т.к. угольная кислота очень плохо диссоциирует (к вопросу о безопасности практики кумбхаки в параняме. Как только дыхание восстанавливается, излишек угольной кислоты тут же удаляется. ALG)
Благодаря работе бикарбонатного буфера концентрация водородных ионов понижается по двум причинам:
• угольная кислота является очень слабой кислотой и плохо диссоциирует
• в крови легких благодаря присутствию в эритроцитах фермента карбоангидразы, угольная кислота быстро расщепляется с образованием CO2, удаляемого с выдыхаемым воздухом:
(и тоже, получается, сто очков в плюс к дыхательной практике, ведь если легкие работают не на полный объем - как это у нас столь распространено - СО2 выделяется не в полном объеме, и эффективность работы этого важнейшего буфера существенно снижается. ALG)
Кроме эритроцитов, значительная активность карбоангидразы отмечена в эпителии почечных канальцев, клетках слизистой оболочки желудка, коре надпочечников и клетках печени, в незначительных количествах – в центральной нервной системе, поджелудочной железе и других органах.
Фосфатная буферная система
Фосфатная буферная система составляет около 1-2% от всей буферной емкости крови и до 50% буферной емкости мочи.
Она образована дигидрофосфатом (NaH2PO4) и гидрофосфатом (Na2HPO4) натрия.
Первое соединение слабо диссоциирует и ведет себя как слабая кислота, второе обладает щелочными свойствами.
В норме отношение HРO42– к H2РO4– равно 4 : 1.
При взаимодействии кислот (ионов водорода) с двузамещенным фосфатом натрия (Na2HPO4) натрий вытесняется, образуется натриевая соль дигидрофосфата (H2PO4–). В результате, благодаря связыванию введенной в систему кислоты, концентрация ионов водорода значительно понижается.
HPO42– + Н-Анион > H2PO4– + Анион–
При поступлении оснований избыток ОН– групп нейтрализуется имеющимися в среде Н+, а расход ионов Н+ восполняется повышением диссоциации NaH2PO4.
H2PO4– + Катион-ОН > Катион+ + HPO42– + Н2О
Основное значение фосфатный буфер имеет для регуляции pH интерстициальной жидкости и мочи.
В моче роль его состоит в сбережении бикарбоната натрия за счет дополнительного иона водорода (по сравнению с NaHCO3) в составе выводимого NaH2PO4:
Na2HPO4 + Н2СО3 > NaH2PO4 + NaНСО3
Кислотно-основная реакция мочи зависит только от содержания дигидрофосфата, т.к. бикарбонат натрия в почечных канальцах реабсорбируется.
Белковая буферная система
Буферная мощность этой системы составляет 5% от общей буферной емкости крови.
Белки плазмы, в первую очередь альбумин, играют роль буфера благодаря своим амфотерным свойствам.
В кислой среде подавляется диссоциация СООН-групп, а группы NH2 связывают избыток Н+, при этом белок заряжается положительно.
В щелочной среде усиливается диссоциация карбоксильных групп, образующиеся Н+ связывают избыток ОН–-ионов и pH сохраняется, белки выступают как кислоты и заряжаются отрицательно.
Гемоглобиновая буферная система
Наибольшей мощностью обладает гемоглобиновый буфер, который можно рассматривать как часть белкового. На него приходится до 30% всей буферной емкости крови.
В буферной системе гемоглобина существенную роль играет гистидин, который содержится в белке в большом количестве.
Изоэлектрическая точка гистидина равна 7,6, что позволяет гемоглобину легко принимать и легко отдавать ионы водорода при малейших сдвигах физиологической рН крови (в норме 7,35-7,45).
Данный буфер представлен несколькими подсистемами:
Пара ННb/ННbО2 является основной в работе гемоглобинового буфера.
Соединение ННbО2 является более сильной кислотой по сравнению с угольной кислотой, HHb - более слабая кислота, чем угольная. Установлено, что ННbО2 в 80 раз легче отдает ионы водорода, чем ННb.
Присоединение ионов водорода к остатку гистидина дезоксигемоглобина выглядит так:
Работа гемоглобинового буфера неразрывно связана с дыхательной системой (к вопросу о значении пранаямы! - ALG)
Установлено, что состоянию нормы соответствует определенный диапазон колебаний рН крови – от 7,37 до 7,44 со средней величиной 7,40 . Кровь представляет собой взвесь клеток в жидкой среде, поэтому ее кислотно-основное равновесие поддерживается совместным участием буферных систем плазмы и клеток крови. Важнейшими буферными системами крови являются бикарбонатная, фосфатная, белковая и наиболее мощная гемогло-биновая.
Бикарбонатная буферная система – мощная и, пожалуй, самая управляемая система внеклеточной жидкости и крови. На долю бикарбонатного буфера приходится около 10% всей буферной емкости крови. Бикарбонатная система представляет собой сопряженную кислотно-основную пару, состоящую из молекулы угольной кислоты Н2СО3, выполняющую роль донора протона, и бикарбонат-иона НСО3 – , выполняющего роль акцептора протона:
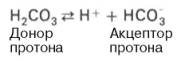
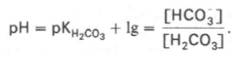
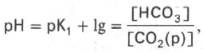
При нормальном значении рН крови (7,4) концентрация ионов бикарбоната НСО3 в плазме крови превышает концентрацию СО2 примерно в 20 раз. Бикарбонатная буферная система функционирует как эффективный регулятор в области рН 7,4.
Механизм действия данной системы заключается в том, что при выделении в кровь относительно больших количеств кислых продуктов водородные ионы Н + взаимодействуют с ионами бикарбоната НСО3 – , что приводит к образованию слабодиссоциирующей угольной кислоты Н2СО3. Последующее снижение концентрации Н2СО3 достигается в результате ускоренного выделения СО2 через легкие в результате их гипервентиляции (напомним, что концентрация Н2СО3 в плазме крови определяется давлением СО2 в альвеолярной газовой смеси).
Если в крови увеличивается количество оснований, то они, взаимодействуя со слабой угольной кислотой, образуют ионы бикарбоната и воду. При этом не происходит сколько-нибудь заметных сдвигов в величине рН. Кроме того, для сохранения нормального соотношения между компонентами буферной системы в этом случае подключаются физиологические механизмы регуляции кислотно-основного равновесия: происходит задержка в плазме крови некоторого количества СО2 в результате гиповентиляции легких . Как будет показано далее, данная буферная система тесно связана с гемоглобиновой системой.
Фосфатная буферная система представляет собой сопряженную кислотно-основную пару, состоящую из иона Н2РО4 – (донор протонов) и иона НРО4 2– (акцептор протонов):
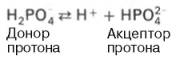
Роль кислоты в этой системе выполняет однозамещенный фосфат NaH2PO4, а роль соли двузамещенный фосфат – Na2HPO4.
Фосфатная буферная система составляет всего лишь 1% от буферной емкости крови. В других тканях эта система является одной из основных. Для фосфатной буферной системы справедливо следующее уравнение:
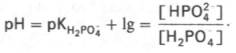
Во внеклеточной жидкости, в том числе в крови, соотношение [НРО4 2– ]: [Н2РО4 – ] составляет 4:1. Величина рКН2РО4– равна 6,86.
Буферное действие фосфатной системы основано на возможности связывания водородных ионов ионами НРО4 2– с образованием Н2РО4 – (Н + + + НРО4 2– —> Н2РО4 – ), а также ионов ОН – с ионами Н2РО4 – (ОН – + + Н2 Р О4 – —> HPO4 2– + H2O). Буферная пара (Н2РО4 – –НРО4 2– ) способна оказывать влияние при изменениях рН в интервале от 6,1 до 7,7 и может обеспечивать определенную буферную емкость внутриклеточной жидкости, величина рН которой в пределах 6,9–7,4. В крови максимальная емкость фосфатного буфера проявляется вблизи значения рН 7,2. Фосфатный буфер в крови находится в тесном взаимодействии с бикарбонатной буферной системой. Органические фосфаты также обладают буферными свойствами, но мощность их слабее, чем неорганического фосфатного буфера.
Белковая буферная система имеет меньшее значение для поддержания КОР в плазме крови, чем другие буферные системы.
Белки образуют буферную систему благодаря наличию кислотно-основных групп в молекуле белков: белок–Н + (кислота, донор протонов) и белок (сопряженное основание, акцептор протонов). Белковая буферная система плазмы крови эффективна в области значений рН 7,2–7,4.
Гемоглобиновая буферная система – самая мощная буферная система крови. Она в 9 раз мощнее бикарбонатного буфера; на ее долю приходится 75% от всей буферной емкости крови.
Участие гемоглобина в регуляции рН крови связано с его ролью в транспорте кислорода и углекислого газа. Константа диссоциации кислотных групп гемоглобина меняется в зависимости от его насыщения кислородом. При насыщении кислородом гемоглобин становится более сильной кислотой (ННbО2). Гемоглобин, отдавая кислород, превращается в очень слабую органическую кислоту (ННb).
Итак, гемоглобиновая буферная система состоит из неионизированного гемоглобина ННb (слабая органическая кислота, донор протонов) и калиевой соли гемоглобина КНb (сопряженное основание, акцептор протонов). Точно так же может быть рассмотрена оксигемоглобиновая буферная система. Система гемоглобина и система оксигемоглобина являются вза-имопревращающимися системами и существуют как единое целое. Буферные свойства гемоглобина прежде всего обусловлены возможностью взаимодействия кисло реагирующих соединений с калиевой солью гемоглобина с образованием эквивалентного количества соответствующей калийной соли кислоты и свободного гемоглобина:
Именно таким образом превращение калийной соли гемоглобина эритроцитов в свободный ННb с образованием эквивалентного количества бикарбоната обеспечивает поддержание рН крови в пределах физиологически допустимых величин, несмотря на поступление в венозную кровь огромного количества углекислого газа и других кисло реагирующих продуктов обмена.
Гемоглобин (ННb), попадая в капилляры легких, превращается в окси-гемоглобин (ННbО2), что приводит к некоторому подкислению крови, вытеснению части Н2СО3 из бикарбонатов и понижению щелочного резерва крови . Перечисленные буферные системы крови играют важную роль в регуляции кислотно-основного равновесия. Как отмечалось, в этом процессе, помимо буферных систем крови, активное участие принимают также система дыхания и мочевыделительная система.
Читайте также: