
Противовирусный иммунитет иммунология доклад
Обновлено: 04.07.2024
Механизмы противовирусного иммунитета имеют особенности, определяемые природой вирусов, паразитизмом на уровне молекулярных и субмолекулярных структур. Внеклеточный вирус не проявляет своего патогенного действия, а иммунные реакции против внеклеточных вирионов сходны с реакциями на другие микроорганизмы.
Защитные реакции, направленные на внутриклеточные стадии развития вируса, действуют не на вирус, а на инфицированную вирусом клетку. Только таким путем происходит подавление репродукции вируса и освобождение от него.
Кардинальные особенности противовирусной защиты - защита клетки от проникшей в нее генетически чужеродной информации и подавление репродукции вируса.
Несмотря на особенности, иммунные реакции в отношении вируса подчиняются тем же закономерностям, что и иммунные реакции в отношении других антигенов.
Особенности инфекционного агента
У РНК-содержащих ретровирусов сначала происходит обратная транскрипция генома в ДНК, затем ее интеграция в клеточные хромосомы и лишь после этого транскрипция генов.
Хотя вирусы и не образуют типичных токсинов, однако и вирионы, и вирусные компоненты, накапливающиеся в пораженных тканях, выходя в кровоток, оказывают токсическое действие. Неменьшее токсическое действие оказывают и продукты распада клеток. В этом случае действие вирусной инфекции столь же неспецифично, как и действие патогенных организмов, убивающих клетки и вызывающих их аутолиз. Поступление токсинов в кровь вызывает ответную реакцию - лихорадку, воспаление, иммунный ответ. Лихорадка является преимущественно рефлекторным ответом на поступление в кровь и воздействие на ЦНС токсичных веществ.
Если лихорадка - общий ответ организма на вирусную инфекцию, то воспаление - это местная многокомпонентная реакция. При воспалении происходят инфильтрация пораженных тканей макрофагами, утилизация продуктов распада, репарация и регенерация. Одновременно развиваются реакции клеточного и гуморального иммунитета.
Механизмы противовирусной защиты можно разделить на факторы резистентности организма, в норме невосприимчивого к определенному виду вируса (обладающего видовой невосприимчивостью), факторы неспецифической защиты восприимчивого организма и факторы приобретенного иммунитета.
Противовирусный иммунитет. Вирусы и иммунная система.
Способность организма распознавать, обезвреживать и удалять чужеродный антиген (иммунореактивность) может значительно колебаться в зависимости от индивидуальных, возрастных и видовых особенностей организма. При встрече с вирусом иммунная система определяет вирусные антигены (белки, гликопротеины) как чужеродные, которые вызывают у нее ответы разного рода с целью удаления вируса и предотвращения реинфекции.
Механизмы, контролирующие различные типы иммунного ответа на вирусные антигены, весьма сложные. Лимфоциты, способные распознавать антигены, являются главными клетками иммунной системы. Их можно разделить на два основных класса: В- и Т-клетки. Как полагают, они происходят из стволовых клеток, находящихся в костном мозгу. Стволовые клетки, пройдя ряд последовательных превращений либо в тимусе, либо в фабрициевой сумке у птиц, либо в ее эквиваленте у млекопитающих, приобретают способность распознавать чужеродные антигены.
Т-клетки можно разделить на несколько субпопуляций, каждая из которых выполняет различные эффекторные или регуляторные функции. Например, цитотоксические Т-клетки (Тц) лизируют клетки, инфицированные вирусами. Регуляторные Т-клетки могут усиливать (хелперные Т-клетки, Тх) или угнетать (супрессорные Т-клетки, Тс) иммунные реакции, в которых участвуют различные популяции лимфоидных клеток. Эти субпопуляции Т-клеток отличаются друг от друга по фенотипической экспрессии определенных поверхностных белков. Имеются данные, подтверждающие существование нескольких функционально отличных субпопуляций В-клеток, различающихся реактивностью на особые антигены. Функция В-клеток проявляется непосредственно или при участии Т-клеток.
Известно, что антитела могут угнетать выработку эффекторных Т-клеток и осуществлять контроль по типу обратной связи.
Антигены являются специфическими митогенами, вызывающими ответные реакции лимфоцитов. После антигенной стимуляции начинается деление и дифференцирование Т- и В-клеток, в результате чего возникают популяции антигенреактивных клеток памяти, также появляются зрелые клетки, осуществляющие эффекторные функции, например, секрецию антител или лизис инфицированных клеток.
Выраженность приобретенного иммунитета определяется Ir (immune response) генами иммунного ответа, расположенными в главном комплексе гис-тосовместимости. Сила иммунного ответа зависит от структурного соответствия вирусного антигена и продуктов этих генов.
В норме иммунная система тонко сбалансирована и обеспечивает индукцию различных типов иммунного ответа и образование Т- и В-клеток памяти. Формирование иммунологической памяти является основной целью иммунизации. Однако, несмотря на развитие иммунного ответа, ряд вирусов вызывает персис-тентные инфекции и использует механизмы, позволяющие избегать иммунного ответа организма.
В процессе эволюции у вирусов выработались различные способности, позволяющие в той или иной степени избегать ответных реакций со стороны иммунной системы. Эти механизмы специфичны для каждой отдельной группы вирусов. Для защиты от вирусной инфекции организм, в свою очередь, использует различные типы иммунитета, и поэтому необходимо как можно полнее изучить разновидности иммунного ответа, требуемые для защиты от разных вирусов. Иммунный ответ организма млекопитающих определяется сложным взаимодействием различных компонентов. Всестороннее изучение его природы -важнейшая предпосылка понимания механизмов действия вакцин и одно из условий их дальнейшего совершенствования. Однако для выработки соответствующих положений иммунизации необходимо точно знать защитные звенья иммунного ответа на каждый конкретный вирус и его иммуногенные компоненты.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
В основе некоторых существенных различий механизмов защиты организма от вирусов и бактерий лежат особенности биологии этих возбудителей.
В защите организма от вирусов участвуют все системы иммунитета, однако противовирусный иммунитет имеет существенные специфические черты. Они определяются тем, что в первую очередь на проникновение вируса в организм реагируют не системы комплемента и макрофагов, а системы интерферонов и Т-киллерных клеток. Другая особенность формирования иммунитета связана с тем, что вирусы оказывают слабое антигенное воздействие на В-лимфоциты и для их активирования, пролиферации и дифференцировки необходимо участие Т-хелперов и соответственно представление последним процессированного вирусного антигена (пептидных фрагментов) при участии молекул МНС класса II. Поэтому роль макрофагов и других антигенпредставляющих клеток заключается не столько в самом фагоцитозе, сколько в процессировании и представлении антигена.
Однако интерферонов и ингибиторов оказалось недостаточно для защиты от вирусов, поэтому природа создала против вирусов другой, очень мощный механизм защиты на уровне организма. Он представлен прежде всего Т-цитотоксическими лимфоцитами и другими киллерными клетками. Эти клетки распознают все чужеродные антигены, в том числе и вирусные, предсталяемые им молекулами МНС класса I. Главное биологическое значение Т-киллерных клеток и заключается в обнаружении и уничтожении любых клеток, инфицированных чужеродными антигенами.
Синтез антител связан, в свою очередь, с системой фагоцитов, В– и Т-лимфоцитов и МНС. В-лимфоциты с помощью иммуноглобулиновых рецепторов распознают соответствующий антиген и отвечают на него синтезом рецепторов, необходимых для распознавания сигналов от Т-хелперов. Активированные Т-хелперы синтезируют и секретируют факторы активации, пролиферации и дифференциации В-лимфоцитов. В результате их действия из активированных В-лимфоцитов формируются клоны антителообразующих клеток и клеток памяти (соответственно возникают и клоны клеток памяти Т-лимфоцитов).
Защитная роль антител в противовирусном иммунитете состоит главным образом в том, что они, взаимодействуя с вирусными рецепторами, исключают возможность адсорбции вирусов на мембране клеток и таким образом нейтрализуют их активность, делают невозможным проникновение вируса в клетку. Только таким путем, т. е. исключая возможность проникновения вируса в клетку, антитела обеспечивают формирование приобретенного иммунитета. Приобретенный противовирусный иммунитет при наличии постоянных клонов клеток памяти может сохраняться пожизненно. Помимо способности нейтрализовать вирусы, антитела выполняют большую роль в освобождении организма от вирусов и вирусных антигенов. Связываясь с ними, антитела образуют иммунные комплексы, которые выводят эти антигены из организма. Эффективность антител в формировании приобретенного иммунитета против вирусных инфекций подтверждена многолетней практикой специфической профилактики полиомиелита, кори, желтой лихорадки, других инфекций и полной ликвидацией на Земле натуральной оспы.
Вместе с тем сами по себе вирусы могут стать причиной вторичных иммунодефицитов. Типичным примером такого вирусного иммунодефицита является ВИЧ-инфекция (см. главу 57).

У РНК-содержащих ретровирусов сначала происходит обратная транскрипция генома в ДНК, затем ее интеграция в клеточные хромосомы и лишь после этого транскрипция генов.
Цитопатические эффекты при вирусных инфекциях разнообразны, они определяются как вирусом, так и клеткой и сводятся к разрушению клетки (цитолитический эффект), сосуществованию вируса и клетки без гибели последней (латентная и персистирующая инфекция) и трансформации клетки.
Вовлеченность организма в инфекционный процесс зависит от ряда обстоятельств - количества погибших клеток, токсичности вирусов и продуктов распада клеток, от реакций организма, начиная от рефлекторных и заканчивая иммунными. Количество погибших клеток влияет на тяжесть инфекционного процесса. Например, будут ли поражены при гриппе только клетки носа и трахеи или вирус поразит клетки эпителия альвеол, зависит тяжесть и исход болезни.
Хотя вирусы и не образуют типичных токсинов, однако и вирионы, и вирусные компоненты, накапливающиеся в пораженных тканях, выходя в кровоток, оказывают токсическое действие. Неменьшее токсическое действие оказывают и продукты распада клеток. В этом случае действие вирусной инфекции столь же неспецифично, как и действие патогенных организмов, убивающих клетки и вызывающих их аутолиз. Поступление токсинов в кровь вызывает ответную реакцию - лихорадку, воспаление, иммунный ответ. Лихорадка является преимущественно рефлекторным ответом на поступление в кровь и воздействие на ЦНС токсичных веществ.
Если лихорадка - общий ответ организма на вирусную инфекцию, то воспаление - это местная многокомпонентная реакция. При воспалении происходят инфильтрация пораженных тканей макрофагами, утилизация продуктов распада, репарация и регенерация. Одновременно развиваются реакции клеточного и гуморального иммунитета. На ранних стадиях инфекции действуют неспецифические киллеры и антитела класса IgM. Затем вступают в действие основные факторы гуморального и клеточного иммунитета. Однако гораздо раньше, уже в первые часы после заражения, начинает действовать система интерферона, представляющая семейство секреторных белков, вырабатываемых клетками организма в ответ на вирусы и другие стимулы. Описанные явления относятся к так называемой острой репродуктивной вирусной инфекции. Взаимодействие вируса и клеток может происходить, как отмечалось выше, без гибели последних. В этом случае говорят о латентной, т.е. бессимптомной или персистирующей хронической вирусной инфекции. Дальнейшая экспрессия вируса, образование вирусспецифических белков и вирионов вызывает синтез антител, на этой стадии латентная инфекция переходит в персистирующую и появляются первые признаки болезни.
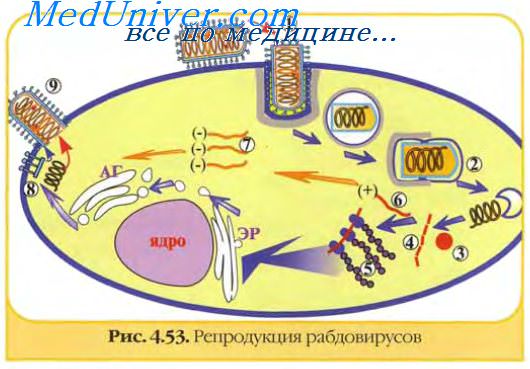
Репродукция вируса в клетках сопровождается развитием цитопатических процессов, специфичных для разных вирусов и для разных типов инфекционных процессов. Цитопатические процессы при вирусных инфекциях разнообразны, они определяются как вирусом, так и клетками, причем специфика их больше "задается" клеткой, нежели вирусом, и сводится в основном к разрушению клеток, сосуществованию вируса и клеток без гибели последних и трансформация клеток. Несмотря на значительные различия цитоцидного действия разных вирусов, в общем, они сходны. Подавление синтеза клеточных макромолекул - нуклеиновых кислот и белков, а также истощение энергетических ресурсов клетки ведут к необратимым процессам, заканчивающимся гибелью пораженной клетки. Повреждение клеток вирусами, их отмирание и распад переносят вирусную инфекцию с клеточного уровня на уровень организма в целом.
При встрече организма с вирусной инфекцией продукция интерферона (растворимого фактора, вырабатываемого вирус-инфицированными клетками, способного индуцировать антивирусный статус в неинфицированных клетках) становится наиболее быстрой реакцией на заражение, формируя защитный барьер на пути вирусов намного раньше специфических защитных реакций иммунитета, стимулируя клеточную резистентность, - делая клетки непригодными для размножения вирусов.
Продукция и секреция цитокинов относятся к самым ранним событиям, сопутствующим взаимодействию микроорганизмов с макрофагами. Этот ранний неспецифический ответ на инфекцию важен по нескольким причинам: он развивается очень быстро, поскольку не связан с необходимостью накопления клона клеток, отвечающих на конкретный антиген; ранний цитокиновый ответ влияет на последующий специфический иммунный ответ.
Интерферон активирует макрофаги, которые затем синтезируют интерферон-гамма, ИЛ-1, 2, 4, 6, ФНО, в результате макрофаги приобретают способность лизировать вирус-инфицированные клетки.
Интерферон-гамма является специализированным индуктором активации макрофагов, который способен индуцировать экспрессию более 100 разных генов в геноме макрофага.
Продуцентами этой молекулы являются активированные Т-лимфоциты (Тh1-тип) и естественные киллеры (NK-клетки). Интерферон-гамма индуцирует и стимулирует продукцию провоспалительных цитокинов (ФНО, ИЛ-1, 6), экспрессию на мембранах макрофагов, антигенов МНС II; гамма-интерферон резко усиливает антимикробную и противовоспалительную активность путем повышения продукции клетками супероксидных радикалов, а усиление иммунного фагоцитоза и антителоопосредованной цитотоксичности макрофагов под влиянием гамма-интерферона связано с усилением экспрессии Fc-рецепторов для JgG. Активирующее действие интерферона-гамма на макрофаги опосредовано индукцией секреции этими клетками ФНО -альфа. Этот пик наблюдается совместно с ФНО-альфа. Максимум продукции ИЛ-4 наступает через 24-48 ч с момента активации клеток. При этом ИЛ-4 рассматривается как цитокин, ограничивающий иммуновоспалительные реакции и снижающий ответ организма на инфекцию, угнетая при этом экспрессию гамма-интерферона. Интерферон-гамма ин витро усиливает фагоцитарную активность нейтрофилов, что обусловлено усилением экспрессии Fc-рецепторов и поверхностных белков семейства интегринов на нейтрофилы. Это позволяет нейтрофилам осуществлять цитотоксические функции и фагоцитоз. В качестве основных эффекторных клеток воспалительного процесса, они обеспечивают элиминацию инфекта из организма.
Взаимодействие цитокина с клеткой определяется универсальной биологической системой, специфическим механизмом которой является рецепторный аппарат, связанный с восприятием метаболического кода. Для проявления биологической активности цитокина необходимо присутствие на поверхности чувствительных клеток специфических рецепторов, которые могут экспрессироваться параллельно с синтезом цитокина. Рецепторы цитокинов представляют собой комплексы, состоящие из двух и более рецепторных молекул, которые объединяются на мембране клетки-мишени и образуют высокоаффинный рецепторный комплекс. Большинство рецепторов состоит из отдельных молекул, связывающих цитокины, которые ассоциируются после связывания лиганда с сигналпередающим рецепторным компонентом; часть рецепторов существует как растворимые изоформы, способные связывать и растворять цитокины, а часть функционирует как многокомпонентные блоки; механизм комплексирования субъединиц рецепторов объясняет плейотропные и дублирующие эффекты цитокинов, имеющих большое структурное сходство. Рецепторы ИЛ-10 имеют гомологию рецепторов интерферона, и подобно ИЛ-10 индуцирует экспрессию в моноцитах гена Fc- рецептора. Для полного функционирования цитокиновой системы необходимы повышение уровня цитокина в ответ на инфект и экспрессия нормального количества рецепторов к ним на клетках. Изменение рецепторов после их связывания с цитокином заключается в интернализации комплексов цитокин - рецептор внутрь клетки. На поверхности клеток рецептор появляется заново, постепенно синтезируясь в течение 24-36 ч (время появления рецепторов интерферон-альфа). В этот период клетки остаются чувствительными к последующим дозам цитокина, чем объясняется эффективность введения препаратов интерферона и их индукторов три раза в неделю.
При антигенной стимуляции клеток трансдукция сигнала с активированного рецептора на генетический аппарат осуществляется с помощью внутриклеточных регуляторных систем, компоненты которых (белки мембран, ферментов, хроматина) связываются с чувствительными к ним последовательностями ДНК. После связывания цитокина (интерферон) с поверхностными клеточными мембранными рецепторами происходит активация ферментов протеинкиназы-С (ПКС), тирозинкиназы, ц-АМФзависимой протеинкиназы, серин-треонинкиназы. Интерферон-альфа активирует tyk 2 и jak 1-киназы, а интерферон-гамма активирует jak 1 и 2-киназы. Далее факторы транскрипции перемещаются в ядро клетки и связывают гены раннего ответа.
Первый ответ клеток на цитокин - это быстрая индукция генов раннего ответа ("immediate early" генов), в число которых и входит ген интерферон-гамма. Стимуляция экспрессии этих генов важна для выхода клеток из Go-стадии и перехода в Gi-стадию и дальнейшей прогрессии клеточного цикла. Их индукция происходит после активации рецепторов роста на клеточной мембране и активации протеин-киназной системы. Гены раннего ответа являются ключевыми регуляторами клеточной пролиферации и дифференцировки, кодируют белки, регулирующие репликацию ДНК.
Таким образом, при активации клеток происходит стимуляция генов раннего ответа, что ассоциируется с изменением фаз клеточного цикла. Основная протективная роль в иммунном ответе, направленном против внутриклеточных паразитов (грибы, простейшие, вирусы, микобактерии туберкулеза), принадлежит клеточным механизмам. Способность перечисленных возбудителей переживать и размножаться внутри клеток делает их защищенными от действия антител и системы комплемента. Резистентность к антимикробным факторам макрофагов позволяет им длительно переживать внутри этих клеток. Для элиминации возбудителя необходим специфический клеточно-опосредованный ответ. Его специфичность определяется антигенраспознающими СД8+-Т-лимфоцитами, которые пролиферируют, активируются и формируют клон эффекторных цитотоксических лимфоцитов. Решающий момент специфического иммунного ответа - это ответ СД4+Т-лимфоцитов с хелперной направленностью на распознавание антигена. На этом этапе определяется форма иммунного ответа: либо с преобладанием гуморального иммунитета, либо с преобладанием клеточных реакций (ГЗТ). Направление дифференцировки СД4 + -лимфоцитов, от которого зависит форма специфического иммунного ответа, контролируется цитокинами, образующимися в ходе воспалительной реакции. Так, в присутствии ИЛ-12 и интерферон-гамма СД4 + -лимфоциты дифференцируются в воспалительные Тh1-клетки, начинают продуцировать и секретировать интерлейкин-2, интерферон-гамма, ФНО и определяют клеточный характер специфического иммунного ответа. Присутствие ИЛ-12 обеспечивается его продукцией макрофагами, а интерферон-гамма - естественными киллерами, активированными в раннюю фазу ответа на внутриклеточно паразитирующие бактерии и вирусы. В отличие от этого, в присутствии ИЛ-4 СД4 + -лимфоциты дифференцируются в хелперы Тh 2, которые начинают продуцировать и секретировать ИЛ-4, ИЛ-5, ИЛ-6 и запускают гуморальный иммунный ответ, т.е. синтез специфических антител - иммуноглобулинов. Воспалительные Тh 1-лимфоциты нужны для борьбы с внутриклеточными паразитами, а Тh 2 хелперы нужны для элективной защиты от внеклеточных паразитов.
Вирусная инфекция может вызывать быстрое подавление экспрессии ряда клеточных генов (из которых наиболее изучены интерфероновые гены и гены, кодирующие дс-РНК-зависимые ферменты -2,5-ОАС и ПК-дс), принимающих участие в антивирусном действии. Специальные исследования механизма антивирусного действия интерферонов и дс-РНК в клеточных и бесклеточных системах показали ключевую роль в этом процессе вышеуказанных ферментов. ПК-дс, взаимодействуя с дс-РНК, фосфорилируется и в активной форме фосфорилирует регуляторные факторы транскрипции и трансляции, из которых наиболее изучен инициирующий фактор трансляции (eIF2).
ПК-дс выполняет регуляторную роль в системе клеточной пролиферации на уровне факторов трансляции и активации ряда генов цитокинов. Вероятно, существует связь между подавлением транскрипции мРНК и ПК-дс, угнетением общего синтеза клеточного белка при вирусных инфекциях и накоплением в ядрах клеток белка нуклеокапсида и белка NSP2. Фрагментация клеточных хромосом, наблюдающаяся на ранних сроках вирусной инфекции, может быть одной из причин подавления экспрессии генов, участвующих в противовирусном ответе.
Есть основания предполагать участие белков NSP2 в регуляции активности генов цитокинов - низкомолекулярных белковых регуляторных веществ, продуцируемых клетками и способных модулировать их функциональную активность. Нарушения в системе цитокинов приводят к нарушению кооперативных взаимодействий иммунокомпетентных клеток и нарушению иммунного гомеостаза.
В последние годы показано, что ИЛ- 12, относящийся к провоспалительным цитокинам, является ключевым для усиления клеточно-опосредованного иммунного ответа и инициации эффективной защиты против вирусов.
Средства терапии гриппа и ОРЗ можно разделить на этиотропные, иммунокорригирующие, патогенетические и симптоматические. Приоритет принадлежит этиотропным препаратам, действие которых направлено непосредственно на возбудитель инфекции. Все препараты этиотропного действия целесообразно рассматривать с учетом их точек приложения в цикле репродукции вирусов гриппа и других ОРЗ.
Применение химиопрепаратов для профилактики и лечения гриппа и ОРЗ относится к базовой терапии и является общепризнанным мировым стандартом. Многолетние клинические исследования достоверно выявили их высокую лечебно-профилактическую значимость. Химиотерапевтические средства представлены тремя основными группами: это блокаторы М2-каналов (амантадин, ремантадин); ингибиторы нейраминидазы (занамивир, озельтамивир) и ингибиторы протеаз (амбен, аминокапроновая кислота, трасилол). Препараты оказывают прямое антивирусное действие, нарушая различные фазы репликативного цикла вирусов. Несколько особняком стоит группа вирулицидных препаратов, применяемых местно для предотвращения адсорбции и проникновения вирионов в клетки.
Читайте также: