
Почему к началу митоза хромосомы состоят из двух хроматид кратко
Обновлено: 05.07.2024
Хромосомы клетки. Митоз и его стадии
Наряду с ДНК в состав хромосом входит много белка, большая часть которого представлена мелкими положительно заряженными молекулами гистонов. Они образуют множество небольших, похожих на катушки структур, которые, располагаясь одна за другой, обвиваются короткими сегментами ДНК.
Среди основных компонентов хромосом есть и негистоновые белки, которые, с одной стороны, являются структурными белками хромосом, а с другой — активаторами, ингибиторами или ферментами в составе регуляторных генетических систем.
Репликация хромосом в полном объеме начинается через несколько минут после завершения репликации ДНК. В течение этого времени вновь синтезированные цепи ДНК объединяются с белками. Две вновь образованные хромосомы до самого конца митоза остаются прикрепленными друг к другу в участке, близком к их центру и называемом центромерой. Такие разделившиеся, но не разошедшиеся хромосомы называют хроматидами.
Процесс деления материнской клетки на две дочерние называют митозом. Вслед за репликацией хромосом с образованием двух хроматид в течение 1-2 ч автоматически начинается митоз.
Одно из самых первых изменений в цитоплазме, связанных с митозом, происходит на поздних стадиях интерфазы и затрагивает центриоли.Центриоли так же, как ДНК и хромосомы, удваиваются во время интерфазы; обычно это происходит незадолго до репликации ДНК. Центриоль длиной около 0,4 мкм и диаметром около 0,15 мкм состоит из девяти параллельных триплетов— трубочек, собранных в виде цилиндра. Центриоли каждой пары лежат под прямым углом друг к другу. Пару центриолеи вместе с прилегающим к ней веществом называют центросомой.
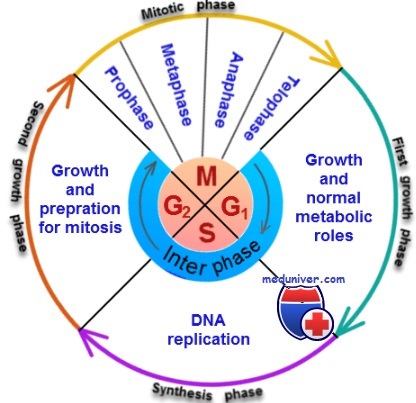
Фазы митоза клетки
Незадолго до начала митоза обе пары центриолей начинают перемещаться в цитоплазме, отдаляясь друг от друга. Это движение обусловлено полимеризацией белка микротрубочек, которые начинают расти от одной пары центриолеи к другой и за счет этого расталкивать их к противоположным полюсам клетки. В то же время от каждой пары центриолеи начинают расти другие микротрубочки, которые увеличиваются в длину и отходят от них радиально в виде лучей, образуя на каждом полюсе клетки так называемую астросферу. Отдельные ее лучи проникают через ядерную оболочку, способствуя таким образом разделению каждой пары хроматид во время митоза. Группу микротрубочек между двумя парами центриолеи называют веретеном деления, а весь набор микротрубочек вместе с центриолями — митотическим аппаратом.
Профаза. По мере образования веретена в ядре начинается конденсация хромосом (в интерфазе они состоят из двух слабосвязанных цепей), которые благодаря этому становятся хорошо различимы.
Прометафаза. Идущие от астросферы микротрубочки разрушают ядерную оболочку . В то же время другие микротрубочки, отходящие от астросферы, прикрепляются к центромерам, которые пока еще соединяют все хроматиды попарно, и начинают тянуть обе хроматиды каждой пары к разным полюсам клетки.
Анафаза. Во время этой фазы две хроматиды каждой пары отрываются друг от друга в области центромеры. Все 46 пар хроматид разделяются и образуют два самостоятельных набора из 46 дочерних хромосом. Каждый набор хромосом движется к противоположным астросферам, а полюсы делящейся клетки в это время расходятся все дальше.
Телофаза. В этой фазе два набора дочерних хромосом полностью расходятся, митотический аппарат постепенно разрушается, вокруг каждого набора хромосом за счет мембраны эндоплазматического ретикулума формируется новая ядерная оболочка. Вскоре после этого между двумя новыми ядрами появляется перетяжка, делящая клетку на две дочерних. Деление обусловлено образованием кольца из микрофиламентов актина и, возможно, миозина (два сократительных мышечных белка) в области перетяжки между дочерними клетками, которое отшнуровывает их друг от друга.

Все клетки человека проходят через цикл деления (клеточный цикл). Известны два типа деления клетки: митотческое и мейотическое, которые представлены на рисунках 6 и 7. Митоз – деление клетки, за счет которого достигается воспроизведение соматических клеток в организме. Мейоз – деление клеток, в ходе которого получаются клетки с редуцированным гаплоидным (n) набором хромосом (половые клетки). Среднее время клеточного цикла у млекопитающих составляет 17–18 часов. Клеточный цикл разделен на четыре основных стадии: G1, S (фаза синтеза ДНК), G2 и митоз (М). Первые три фазы представляют собой интерфазный период или интерфазу. Клетки, которые не делятся, находятся на стадии покоя G0.
Во время стадии G1 (примерно 9 часов) хромосомы имеют вид одиночных хроматид, клетка метаболически активна и в ней происходит синтез белков. Если клетка, находящаяся на G1 стадии, не подвергается последующему делению, то это состояние соответствует G0. Фаза синтеза (или S фаза) длится около 5 часов и характеризуется процессом репликации хромосомной ДНК. На этой стадии хромосомы состоят из двух идентичных сестринских хроматид. G2 стадия длится примерно 3 часа. Во время этой стадии клетка готовится к процессу деления. Завершение G2 соответствует концу интерфазы. Митоз длится не более 1–2 часов и является процессом образования двух генетически идентичных дочерних клеток. В свою очередь, митоз также делится на 4 стадии: профаза, метафаза, анафаза и телофаза (рис. 6).
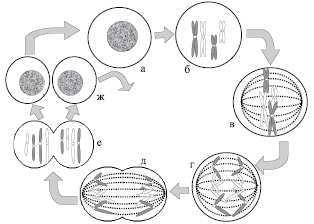
Рис. 6. Митоз. Схематическое изображение двух пар хромосом во время митоза: а – интерфаза; б – профаза; в – метафаза; г – анафаза; д – телофаза; е – цитокинез; ж – интерфазы разделившихся клеток
Профаза – стадия, во время которой происходит постепенная конденсация (уплотнение) и спирализация хромосом, в результате чего, они имеют вид дискретных структур. Во время профазы образуется веретено деления (двуполюсное веретено, состоящее из пучков микротрубочек, которые тянутся от одного полюса к другому).
Метафаза – стадия, характеризующаяся тем, что полностью осуществляется присоединение хромосом к нитям веретена, и хромосомы собираются в экваториальной плоскости клетки, находящейся на одинаковом расстоянии от обоих полюсов веретена. На этой стадии хромосомы достигают максимальной конденсации.
Цитогенетики выделяют начало метафазы как прометафазу, на которой довольно часто проводят лабораторные исследования. Прометафаза – короткий промежуточный период между профазой и метафазой, в ходе которой исчезает ядерная мембрана и появляются нити веретена деления, к которым с помощью кинетохоров присоединяются хромосомы.
Анафаза – стадия разделения сестринских хроматид и их расхождения к противоположным полюсам веретена.
Телофаза – последняя стадия митоза. Она начинается, когда все сестринские хроматиды доходят до полюсов веретена. В ходе данной фазы митоза происходит восстановление клеточного ядра и внутриядерных структур. За ней, как правило, следует цитокинез – разделение двуядерной клетки на две с одним ядром в каждой (рис. 6).
Мейоз – деление клеток, в ходе которого получаются клетки с редуцированным гаплоидным набором хромосом (половые клетки). Данный процесс включает в себя две фазы клеточного деления: мейоз 1 и мейоз 2. В ходе мейоза происходит редукция диплоидного хромосомного набора (2n) до гаплоидного (n).
Мейоз 1, в свою очередь также делится на несколько стадий: профаза I, метафаза I, анафаза I и телофаза I (рис. 7). Профаза I является комплексной стадией, которая, в свою очередь, делится на несколько этапов:
лептотена: 46 хромосом состоят из 2-х хроматид и начинают коденсироваться; эта стадия характеризует начало мейоза;
зиготена: гомологичные хромосомы спариваются своими участками – этот этап деления называется синапсом; в результате образуется тройственная структура – синаптонемальный комплекс;
пахитена: стадия завершения синапса, спаренные гомологи, состоящие из 4-х хроматид, формируют бивалент; во время этой стадии происходит кроссинговер – обмен последовательностями ДНК хромосом между несестринскими хроматидами бивалентов. Результатом этого процесса является рекомбинация генетического материала между гомологичными хромосомами, создающая новые комбинации генов в дочерних клетках;
диплотена: на этой стадии хромосомы отталкиваются друг от друга до тех пор, пока гомологи не будут соединены только участками, подверженными кроссинговеру. Такие участки называются хиазмами;
диакинез: хромосомы претерпевают наибольшее сжатие во время этой последней стадии профазы I.
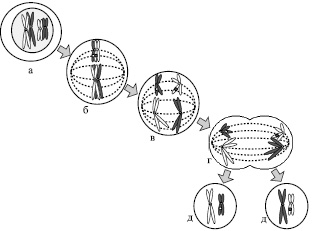
Рис. 7. Мейоз I. Схематическое изображение двух пар хромосом во время мейоза I: а – профаза I; б – метафаза I; в – анафаза I; г – телофаза I; д – клетки, образующиеся в результате первого мейоза
Метафаза I характеризуется исчезновением ядерной мембраны и образованием мейотического веретена деления. Биваленты выравниваются по экваториальной плоскости клетки и их центромеры случайным образом ориентируются к противоположным полюсам. Во время анафазы I биваленты разделяются и расходятся к противоположным полюсам. В ходе телофазы I каждая хромосома из двух гаплоидных наборов достигает противоположных полюсов, и образуются две дочерние клетки, в каждой из которых по 23 хромосомы, состоящие из 2-х хроматид.
Мейоз 2 практически идентичен митотическому делению за исключением того, что в данном случае делящиеся клетки имеют гаплоидный хромосомный набор. Хромосомы выравниваются по экваториальной плоскости клетки на стадии метафазы II, хроматиды разделяются и расходятся к противоположным полюсам на стадии анафазы II, цитокинез происходит на стадии телофазы II. В результате митотического деления (мейоза 1 и 2), как правило, образуются 4 дочерние клетки с гаплоидным набором хромосом, каждая из которых генетически отличается друг от друга за счет процесса кроссинговера и случайного расхождения гомологичных хромосом.
Подготовка клетки к митозу происходит в интерфазу: удваивается ДНК, накапливается АТФ, синтезируются белки веретена деления, удваиваются центриоли.
В схемах деления гаплоидный набор хромосом обозначают буквой n , а молекул ДНК (т. е. хроматид ) — буквой с . Перед буквами указывают число гаплоидных наборов:
в клетках человека гаплоидный набор составляют \(23\) хромосомы. Значит, запись 2n2с обозначает \(46\) хромосом и \(46\) хроматид, а 2n4с — 46 хромосом и 92 хроматиды и т. д.



В результате митоза образуются генетически одинаковые дочерние клетки с тем же набором хромосом, что был у материнской клетки. Сохраняется преемственность в ряду клеточных поколений.

Чтобы жизненный цикл эукариот, обладающих половым процессом, мог продолжаться, должен был развиться механизм, посредством которого из диплоидных клеток снова могли образовываться гаплоидные. Таким механизмом стал мейоз – особый вид клеточного деления, при котором число хромосом в дочерних клетках уменьшается вдвое по сравнению с родительской клеткой.
В статье Ю.Ф.Богданова, крупнейшего специалиста по эволюции мейоза, рассматриваются современные представления о происхождении механизмов мейотического деления у эукариот.

В ходе митоза (а также в ходе второго деления мейоза) хромосомы вступают в метафазу поодиночке, и нити веретена деления присоединяются к каждой хромосоме с двух сторон.
Именно благодаря указанным особенностям первого деления мейоза и обеспечивается уменьшение числа хромосом: к полюсам клетки расходятся не сестринские хроматиды, а гомологичные хромосомы, по одной из каждой пары (см. схему).
Эти ключевые особенности первого деления мейоза развились на основе одного и того же ароморфоза – возникновения так называемого синаптонемного комплекса (СК).
Одним из этих исходных блоков стал уже имевшийся к тому времени у эукариот механизм митоза, другим – механизм репарации (починки повреждений) ДНК, основанный на гомологичной рекомбинации (см.: Глазер В.М. Гомологичная генетическая рекомбинация ). Суть процесса в том, что поврежденный участок одной молекулы ДНК заменяется его неповрежденной копией, взятой из другой (гомологичной) молекулы ДНК.

Упрощенная схема одного из вариантов гомологичной рекомбинации (обмена участками между гомологичными молекулами ДНК) с образованием соединения Холлидея. Показаны две двухцепочечные молекулы ДНК. Тут важно не перепутать: каждая молекула ДНК состоит из двух параллельных цепей нуклеотидов (двойная спираль); в состав одной хроматиды входит одна молекула ДНК; каждая хромосома (вплоть до анафазы II) состоит из двух хроматид. Соответственно, каждая пара гомологичных хромосом (например, в метафазе I) состоит из ДВУХ хромосом, ЧЕТЫРЕХ хроматид, ЧЕТЫРЕХ молекул ДНК, ВОСЬМИ нуклеотидных цепочек. Рекомбинация происходит между хроматидами отцовской и материнской хромосом. На этом рисунке показана рекомбинация у одного из вирусов, у эукариот во время мейоза все несколько иначе и сложнее
Починка разрывов ДНК – одна из основ мейоза
Механизмы репарации и рекомбинации ДНК – очень древние, они должны были возникнуть еще на заре жизни.
Жизненный цикл древних эукариот (как и многих прокариот), вероятно, состоял из двух фаз и двух соответствующих типов метаболизма: 1) вегетативная фаза (условия благоприятны, клетки размножаются митозом); 2) фаза споруляции (условия стали неблагоприятными, клетки превращаются в споры). У низших эукариот – например, у дрожжей, - перед споруляцией происходит мейоз, и из получившихся гаплоидных клеток образуются споры.
Становление мейоза в ходе эволюции, так же как и сам мейоз в жизненном цикле современных организмов, начинается с репарации повреждений (разрывов) ДНК. Разрывы эти в начале мейоза возникают не сами собой – их создает специальный фермент, эндонуклеаза SPO11. Этот фермент является модификацией другого белка – ДНК-топоизомеразы VI, унаследованной эукариотами от своих прокариотических предков – архей.
Белковые оси хромосом
В клетках эукариот, делящихся путем митоза, сестринские хроматиды (образовавшиеся в результате репликации исходной молекулы ДНК) соединены посредством белков-когезинов. Чтобы хроматиды могли разойтись к полюсам клетки (в анафазе митоза), когезины должны гидролизоваться и исчезнуть из межхроматидного пространства.
Синаптонемные комплексы. Ароморфоз.
Происхождение белков СК


Сравнительный анализ структуры белков СК показал, что они возникли независимо у предков современных эукариот, давших начало разным крупным таксонам (растений, грибов, животных).
Почему в первом делении мейоза сестринские хроматиды не расходятся?
С фактами и теориями, касающимися происхождения мейоза, можно ознакомиться в статьях Ю.Ф.Богданова и его коллег:
Читайте также: