
Нервная регуляция деятельности мышц кратко
Обновлено: 02.07.2024
Функция поддержания мышечного тонуса обеспечивается по принципу обратной связи на различных уровнях регуляции организма Периферическая регуляция осуществляется с участием гамма-петли, в состав которой входят супраспинальные моторные пути, вставочные нейроны, нисходящая ретикулярная система, альфа- и гамма-нейроны.
Существует два типа гамма-волокон в передних рогах спинного мозга. Гамма-1-волокна обеспечивают поддержание динамического мышечного тонуса, т.е. тонуса, необходимого для реализации процесса движения. Гамма-2-волокна регулируют статическую иннервацию мышц, т.е. осанку, позу человека. Центральная регуляция функций гамма-петли осуществляется ретикулярной формацией через ретикулоспинальные пути. Основная роль в поддержании и изменении мышечного тонуса отводится функциональному состоянию сегментарной дуги рефлекса растяжения (миотатического, или проприоцептивного рефлекса). Рассмотрим его подробнее.
Рецепторным элементом его является инкапсулированное мышечное веретено. Каждая мышца содержит большое количество этих рецепторов. Мышечное веретено состоит из интрафузальных мышечных волокон (тонких) и ядерной сумки, оплетенной спиралевидной сетью тонких нервных волокон, представляющих собой первичные чувствительные окончания (анулоспинальная нить). На некоторых интрафузальных волокнах имеются также и вторичные, гроздевидные чувствительные окончания. При растяжении интрафузальных мышечных волокон первичные чувствительные окончания усиливают исходящую из них импульсацию, которая через быстропроводящие гамма-1-волокна проводится к альфа-большим мотонейронам спинного мозга. Оттуда, через также быстропроводящие альфа-1-эфферентные волокна, импульс идет к экстрафузальным белым мышечным волокнам, которые обеспечивают быстрое (фазическое) сокращение мышцы. От вторичных чувствительных окончаний, реагирующих на тонус мышцы, афферентная импульсация проводится по тонким гамма-2-волокнам через систему вставочных нейронов к альфа-малым мотонейронам, которые иннервируют тонические экстрафузальные мышечные волокна (красные), обеспечивающие поддержание тонуса и позы.
Интрафузальные волокна иннервируются гамма-нейронами передних рогов спинного мозга. Возбуждение гамма-нейронов, передаваясь по гамма-волокнам к мышечному веретену, сопровождается сокращением полярных отделов интрафузальных волокон и растяжением их экваториальной части, при этом изменяется исходная чувствительность рецепторов к растяжению (происходит снижение порога возбудимости рецепторов растяжения, и усиливается тоническое напряжение мышцы).
Гамма-нейроны находятся под влиянием центральных (супрасегментарных) воздействий, передающихся по волокнам, которые идут от мотонейронов оральных отделов головного мозга в составе пирамидного, ретикулоспинального, вестибулоспинального трактов.
При этом если роль пирамидной системы заключается преимущественно в регуляции фазических (т.е. быстрых, целенаправленных) компонентов произвольных движений, то экстрапирамидная система обеспечивает их плавность, т.е. преимущественно регулирует тоническую иннервацию мышечного аппарата. Так, по мнению J. Noth (1991), спастичность развивается после супраспинального или спинального поражения нисходящих двигательных систем при обязательном вовлечении в процесс кортикоспинального тракта [2] .
В регуляции мышечного тонуса принимают участие и тормозные механизмы, без которых невозможно реципрокное взаимодействие мышц-антагонистов, а значит, невозможно и совершение целенаправленных движений. Они реализуются с помощью рецепторов Гольджи, расположенных в сухожилиях мышц, и вставочных клеток Реншоу, находящихся в передних рогах спинного мозга. Сухожильные рецепторы Гольджи при растяжении или значительном напряжении мышцы посылают афферентные импульсы по быстропроводящим волокнам 1б-типа в спинной мозг и оказывают тормозящее воздействие на мотонейроны передних рогов. Вставочные клетки Реншоу активизируются через коллатерали при возбуждении альфа-мотонейронов, и действуют по принципу отрицательной обратной связи, способствуя торможению их активности. Таким образом, нейрогенные механизмы регуляции мышечного тонуса многообразны и сложны.
При поражении пирамидного пути растормаживается гамма-петля, и любое раздражение путем растяжения мышцы приводит к постоянному патологическому повышению мышечного тонуса. При этом поражение центрального мотонейрона приводит к снижению тормозных влияний на мотонейроны в целом, что повышает их возбудимость, а так же на вставочные нейроны спинного мозга, что способствует увеличению числа импульсов, достигающих альфа-мотонейронов в ответ на растяжение мышцы [4].
В качестве других причин спастичности можно указать структурные изменения на уровне сегментарного аппарата спинного мозга, возникающие вследствие поражения центрального мотонейрона: укорочение дендритов альфа-мотонейронов и коллатеральный спрауттинг (разрастание) афферентных волокон, входящих в состав задних корешков.
Возникают так же и вторичные изменения в мышцах, сухожилиях и суставах. Поэтому страдают механико-эластические характеристики мышечной и соединительной ткани, которые определяют мышечный тонус, что еще больше усиливает двигательные расстройства.
Следствием этого является спастичность мышц, гиперрефлексия, появление патологических рефлексов, а также первоочередная утрата наиболее тонких произвольных движений [5].
Наиболее значимым компонентом мышечного спазма является боль. Болевая импульсация активирует альфа- и гамма-мотонейроны передних рогов, что усиливает спастическое сокращение мышцы, иннервируемой данным сегментом спинного мозга. В то же время, мышечный спазм, возникающий при сенсомоторном рефлексе, усиливает стимуляцию ноцицепторов мышцы. Так, по механизму отрицательной обратной связи формируется замкнутый порочный круг: спазм – боль – спазм – боль [3].
Интерес представляют также данные о центральных холинергических механизмах регуляции мышечного тонуса. Показано, что клетки Реншоу активируются ацетилхолином как через коллатерали мотонейрона, так и через ретикулоспинальную систему.
M.Schieppati и соавт., (1989) установили, что фармакологическая активация центральных холинергических систем значительно снижает возбудимость альфа-мотонейронов путем повышения активности клеток Реншоу.
В последние годы исследователи регуляции мышечного тонуса придают огромное значение роли нисходящих адренергических супраспинальных путей, начинающихся в области голубого пятна. Анатомически эти образования тесно связаны со спинальными структурами, особенно с передними рогами спинного мозга. Норадреналин, высвобождаемый с терминалей бульбоспинальных волокон, активизирует адренорецепторы, располагающиеся во вставочных нейронах, первичных афферентных терминалях и мотонейронах и воздействует одновременно на альфа- и бета-адренорецепторы в спинном мозге (D.Jones et al., 1982). К ядерным образованиям ретикулярной формации ствола подходят многочисленные аксоны болевой чувствительности. На основе информации, поступающей в ретикулярную формацию ствола головного мозга, выстраиваются соматические и висцеральные рефлексы. От ядерных образований ретикулярной формации формируются связи с таламусом, гипоталамусом, базальными ядрами и лимбической системой, которые обеспечивают реализацию нейроэндокринных и аффективных проявлений боли, что особенно важно при хронических болевых синдромах [2].
В итоге формирующийся порочный круг включает в себя мышечный спазм, боль, локальную ишемию, дегенеративные изменения, которые самоподдерживают друг друга, усиливая первопричину патологических изменений.
Следует учитывать, что чем больше компонентов этого порочного круга становятся мишенями при лечении, тем выше вероятность его успеха. Поэтому современными требованиями к миорелаксирующей терапии являются: мощность миорелаксирующего действия, его селективность, наличие противосудорожного и антиклонического эффектов, мощность анальгетического действия, а так же безопасность и наличие широкого терапевтического диапазона доз препарата.
Согласно современным представлениям, большинство миорелаксантов воздействуют на трансмиттеры или нейромодуляторы ЦНС. Воздействие может включать супрессию возбуждающих медиаторов (аспартат и глутамат) и/или усиление тормозных процессов (ГАМК, глицин). Далее

Рецепторным элементом миотатического рефлекса является инкапсулированное мышечное веретено. Каждая мышца содержит большое количество этих рецепторов. Мышечное веретено состоит из интрафузальных мышечных волокон (в сравнении с обычными экстрафузальными мышечными волокнами они значительно тоньше) и ядерной сумки, оплетенной спиралевидной сетью тонких нервных волокон, представляющих собой первичные чувствительные окончания. На некоторых интрафузальных волокнах имеются еще и вторичные, гроздевидные чувствительные окончания. При растяжении интрафузальных волокон первичные чувствительные окончания усиливают исходящую из них импульсацию, которая через быстро проводящие афферентные волокна типа Iа проводятся к альфа-большим мотонейронам спинного мозга. Оттуда через также быстропроводящие альфа-1-эфферентные волокна импульс идет к экстрафузальным белым мышечным волокнам, которые обеспечивают быстрое (фазическое) сокращение мышцы. От вторичных чувствительных окончаний, реагирующих на тонус мышцы, афферентная импульсация проводится по тонким волокнам II типа через систему вставочных нейронов к альфа-малым мотонейронам, которые иннервируют тонические экстрафузальные мышечные волокна (красные мышечные волокна), обеспечивающие поддержание тонуса и позы.
В регуляции мышечного тонуса принимают участие и тормозные механизмы, которые реализуются с помощью рецепторов Гольджи, расположенных в сухожилиях мыщц, и вставочных клеток Реншоу, находящихся в передних рогах спинного мозга. Сухожильные рецепторы Гольджи при растяжении или значительном напряжении мышцы посылают афферентные импульсы, которые проводятся по быстропроводящим волокнам 1b типа в спинной мозг и оказывают тормозящее воздействие на мотонейроны передних рогов. Вставочные нейроны Реншоу активизируются через коллатерали при возбуждении альфа-мотонейронов и действуют по принципу отрицательной обратной связи, тормозя активность последних. Таким образом, нейрогенные механизмы регуляции мышечного тонуса многообразны и сложны.
Высшие двигательные центры коры (двигательная зона лобной доли) свои импульсы отправляют глубоко в головной мозг, в мозжечок, а также к исполнительным нейронам спинного мозга. В целом именно кора головного мозга создает программы для сложных движений, импульсы от нее идут в спинной мозг, который отдает команды для конкретных движений.
Существует и гуморальная регуляция мышц — посредством биологически активных веществ, содержащихся в крови, например, ионов кальция. Причиной сокращения внутри мышечного волокна белковых нитей актина и миозина выступает химическое взаимодействие между этими белками в присутствии АТФ и ионов кальция. Ионы в клетке накапливаются в полости гладкой ЭПС.
Работа мышц
1. Сокращение мышц. В каждом своем волокне скелетные мышцы изолированно передают возбуждение. Сила сокращения мышцы зависит от того, сколько мышечных волокон сократилось в данный момент.
Утомление — падение работоспособности мышцы. Его темпы зависят от характера деятельности, величины нагрузки, ритмичности движений. Активная работа мышц с высокой частотой сокращений снижает силу этих сокращений. Чем шустрее вы будете работать какой-то мышцей, тем быстрее в ней станет падать сила. При отдыхе работоспособность возвращается, идет восстановление.
Активный отдых и работа мышц
1. Однако физиолог И.М. Сеченов обнаружил, что восстановление происходит быстрее, если чередовать нагрузки на разные группы мышц — то есть при активном отдыхе.
2. Наиболее оптимальны средние нагрузки и средняя же скорость сокращения мышц.
3. Выделяют статическую и динамическую работу скелетной мышцы. Статическая работа мышц — поддержание позы, удержание груза без изменения положения тела.
4. При статической работе утомление развивается быстрее. Попробуйте взять тяжелую гантель и удерживать вытянутую руку с грузом на уровне плеча — уж через пару десятков секунд вы почувствуете боль в мышцах.
5. При динамической же работе, то есть разнообразных движениях руки с гантелью, утомление наступает медленнее. Кстати, долгое сидение за компьютером (статическая работа) очень плохо влияет на мышцы спины — они затекают, деревенеют. Отдыхом для них будет несколько энергичных упражнений.
Как выглядит гигиена физического труда, его правильная организация? Необходимо менять нагрузку на мышцы, давать им восстановиться, нагружая другие — это и есть активный отдых. Нужно чаще, и лучше регулярно, делать гимнастику, выбирать правильный ритм физической работы.
Гладкие мышцы
Строение гладких мышц отлично от скелетных — они не имеют поперечных полос. Находятся в составе стенок желудка, кишечника, мочевого пузыря, матки, большинства кровеносных сосудов. Сокращаются медленно — десятки секунд. Но энергии тратят меньше, выделяют меньше продуктов обмена. Сокращаться могут долго, утомление почти не наблюдается. При этом тип сокращений — непроизвольный, человек не может как-то воздействовать на гладкую мускулатуру. Клетки этой ткани сохраняют способность к делению.
Хочешь сдать экзамен на отлично? Жми сюда - видео подготовка к ОГЭ по биологии
Нервно-мышечные соединения в гладких мышцах. Гуморальная регуляция сокращений гладких мышц
Волокна скелетной мышцы стимулируются исключительно нервными сигналами, но сокращения гладкой мышцы могут вызываться множеством типов сигналов: нервными, гормональными, растяжением мышцы и некоторыми другими. Основная причина различий состоит в том, что гладкомышечная мембрана содержит много типов рецепторных белков, способных запустить сократительный процесс. Кроме того, есть и другие рецепторные белки, тормозящие сокращение гладкой мышцы, что также отличает ее от скелетной мышцы. Обсудим сначала нервную регуляцию гладкомышечного сокращения, а затем гормональный контроль и другие способы управления.
Физиологическая анатомия нервно-мышечных соединений гладких мышц. Сложно организованные нервно-мышечные соединения, обнаруживаемые на волокнах скелетных мышц, в гладких мышцах не встречаются. Вместо этого вегетативные нервные волокна, иннервирующие гладкие мышцы, обычно диффузно разветвляются на наружной поверхности пласта мышечных волокон. В большинстве случаев эти волокна не входят в непосредственный контакт с клеточными мембранами гладкомышечных волокон, а формируют так называемые диффузные соединения, которые секретируют медиаторы в матрикс, покрывающий гладкую мышцу, часто на расстоянии от нескольких нанометров до нескольких микрометров от мышечных клеток; затем медиатор диффундирует к клеткам.
Более того, при наличии многих слоев мышечных клеток нервные волокна часто иннервируют только наружный слой, и возбуждение распространяется от этого наружного слоя к внутренним слоям путем проведения потенциалов действия по мышечной массе или путем дополнительной диффузии медиатора.
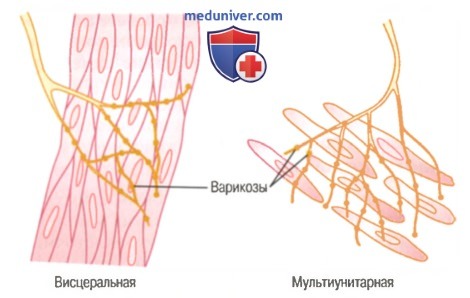
Иннервация гладких мышц.
Аксоны, иннервирующие гладкомышечные волокна, не имеют типичных ветвящихся окончаний, характерных для двигательной концевой пластинки волокон скелетной мышцы. Вместо этого большинство тонких концевых частей аксонов имеют множество варикозных расширений (варикозов), распределенных вдоль их осей. В этих местах связь между шванновскими клетками, окутывающими аксоны, прерывается, и медиатор может секретироваться через стенки варикозов. В варикозах находятся везикулы, которые подобно везикулам в концевой пластинке скелетных мышц содержат медиатор. Но в противоположность везикулам в соединениях скелетных мышц, которые всегда содержат ацетилхолин, везикулы окончаний вегетативных нервных волокон содержат ацетилхолин в одних волокнах и норадреналин — в других, а иногда и другие вещества.
В некоторых случаях, особенно в гладких мышцах мультиунитарного типа, варикозы отдалены от мембраны мышечных клеток на 20-30 нм, что равно ширине синаптической щели в соединении скелетной мышцы. Такие соединения называют контактными, и они функционируют во многом так же, как нервно-мышечные соединения скелетных мышц. Скорость сокращения этих гладкомышечных волокон значительно больше, чем у волокон, стимулируемых диффузными соединениями.
Возбуждающие и тормозящие медиаторы, секретируемые в нервно-мышечных соединениях гладких мышц. Самыми важными медиаторами, которые секретируются вегетативными нервами, иннервирующими гладкие мышцы, являются ацетилхолин и норадреналин, однако они никогда не выделяются одними и теми же нервными волокнами. Ацетилхолин для гладких мышц одних органов является возбуждающим медиатором, а на гладкие мышцы других органов действует как тормозящий агент. Если ацетилхолин возбуждает мышечное волокно, норадреналин обычно тормозит его. И наоборот, если ацетилхолин тормозит волокно, норадреналин, как правило, его возбуждает.
Но почему возникают такие разные реакции? Ответ заключается в том, что ацетилхолин и норадреналин возбуждают или тормозят гладкую мышцу, связываясь сначала с рецепторным белком на поверхности мембраны мышечной клетки. Некоторые из этих рецепторных белков являются возбуждающими рецепторами, тогда как другие — тормозящими рецепторами. Следовательно, тип рецептора определяет, как будет реагировать гладкая мышца — торможением или возбуждением, а также какой из двух медиаторов (ацетилхолин или норадреналин) будет проявлять возбуждающее или тормозящее действие.
Видео физиология гуморальной регуляции и ее отличие от гормональной - профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Читайте также:
- Что такое операторы в информатике кратко
- К каким последствиям привело бы осуществление этого проекта кратко
- Что значит выражение работать на совесть 5 класс кратко
- Исследовательская деятельность в физической культуре в доу
- Объясните заголовок разделенная италия подумайте какие черты традиционного общества кратко