
Какую группу вымерших растений считают предками голосеменных кратко
Обновлено: 05.07.2024
Прочитайте текст и составьте план рассказа о древних голосеменных растениях.
Значительная часть древних голосеменных вымерла. Видовое разнообразие современных голосеменных всего около 700 видов.
Семенные папоротники — вымершая группа голосеменных. Они были похожи на настоящие папоротники как по строению перистых листьев, так и по внешнему облику, за что и получили своё название. Но размножались они с помощью семян. Предполагают, что от семенных папоротников произошли примитивные группы голосеменных, такие, как саговниковые, гинкговые и вымершие беннеттитовые.
Саговниковые появились ещё в каменноугольном периоде. Их ошибочно называют саговыми пальмами. Растут они в тропических и субтропических районах земного шара. Это внешне похожие на пальмы растения, иногда достигающие 20 м в высоту. У некоторых видов наземный стебель не формируется совсем и перистые листья непосредственно отходят от корневища (рис. 47 ).
На верхушках стеблей развиваются огромные, иногда метровые, разнополые шишки (по−научному шишки голосеменных называют стробилами). Оплодотворение происходит с помощью жгутиковых сперматозоидов; яйцеклетка огромна (до 6 мм). Жгутики помогают сперматозоидам двигаться в воде в направлении яйцеклетки. Сохранение столь примитивного признака показывает, что нет резкой грани между половым размножением папоротниковидных и голосеменных. У большинства саговников семена крупные, сочные и употребляются местными жителями в пищу.
Единственный современный представитель класса Гинкговые − реликтовое растение гинкго двулопастный (рис. 48 ).
Это растение называют живым ископаемым, так как его ближайшие родичи вымерли десятки миллионов лет назад. В дикорастущем состоянии он встречается только в горах Западного Китая. Гинкго — листопадное дерево, достигающее более 30 м в высоту, с диаметром ствола более 3 м.
Гинкго очень долговечен, доживает до 2000 лет. Растение это двудомное, то есть у него имеются мужские и женские экземпляры. Яйцеклетка оплодотворяется сперматозоидами.
Довольно крупные семена гинкго съедобны и, несмотря на неприятный запах, высоко ценятся в восточной кухне.
Беннеттитовые представляют особый интерес, так как они могли быть предками цветковых. Большинство беннеттитовых имели обоеполые шишки (стробилы), по типу строения напоминающие цветок наиболее примитивных из ныне живущих покрытосеменных. По внешнему виду они были сходны с саговниками.
Рис. 47 . Саговник
Рис. 48 . Гинкго
Решение
План:
1 . Голосеменные.
2 . Семенные папоротники.
а. внешний облик
3 . Примитивные группы голосеменных.
а. Саговниковые.
б. Гинкговые.
в. Беннеттитовые.
4 . Саговниковые.
а. Среда обитания.
б. Внешний вид.
в. Размножение.
г. Значимость для человека.
5 . Гинкговые.
а. Среда обитания.
б. Внешний вид.
в. Размножение.
г. Значимость для человека.
6 . Беннеттитовые.
а. Вымерший вид.
б. Предки цветковых.
в. Внешний вид.
Возникновению голосеменных растений предшествовало несколько важных эволюционных событий, которые произошли в течение девонского периода. В области репродуктивной сферы это было возникновение разносноровости. Разноспоровые формы известны уже начиная со среднего девона (около 400 млн. лет назад). У шалёрии (Chaleuria cirrosa) из среднего девона Нью-Брансуика (Канада) одни спорангии содержат либо преимущественно крупные споры, либо преимущественно маленькие или смесь тех и других. В позднем девоне появляются растения с еще более ясно выраженной разноспоровостью. В области вегетативной сферы это было возникновение камбия и появление древесных форм. Хорошо сохранившаяся вторичная ксилема, свидетельствующая о камбиальной активности, также известна уже из отложений среднего девона (эйфельский век). Следовательно, камбиальная активность возникла немногим менее 400 млн. лет назад. Несколько позднее (живетский век среднего девона) возникают первые известные нам древовидные формы. Одним из первых был аневрофит германский (Aneurophyton germanicum), достигавший более 10 м высоты. К концу среднего девона появляется другое древесное растение — знаменитый археоптерис (Archaeopteris latifolia) — крупное дерево, вероятно, высотой более 30 м, с кроной ветвей, несущих спирально расположенные листья (рис. 94).
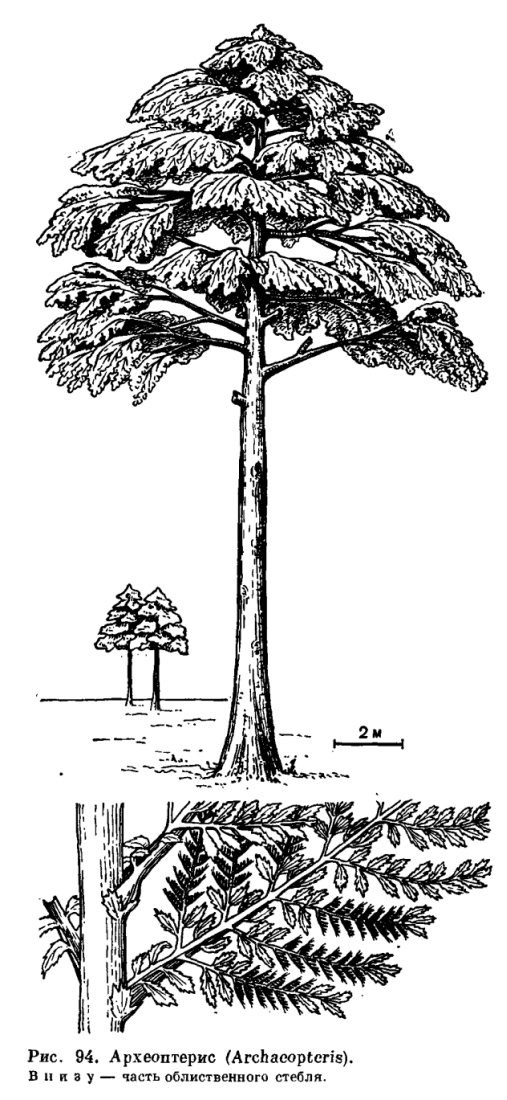
Из факта нахождения голосеменных в верхнем девоне следует, что предков голосеменных следует искать среди палеозойских растений, возраст которых не моложе позднего девона. Тем самым из числа возможных предков голосеменных выпадают почти все типичные папоротники, от которых нередко пытались вывести голосеменные. В качестве их возможных предков остается рассматривать только древнейших представителей отдела папоротниковидных. Именно среди них мы находим разноспоровые древовидные формы с вторичной древесиной, которые могли дать начало голосеменным.
Нередко высказывается предположение, что голосеменные могли произойти от одного из наиболее высокоорганизованных девонских папоротниковидных, а именно от рода археоптерис или ближайших к нему форм. Но у археоптериса уже имеется сердцевина, вторичная ксилема у него сильно развита и толстая, а трахеиды точечные (с округлыми окаймленными порами). По уровню организации своих вегетативных органов археоптерис мало отличается не только от некоторых пермокарбоновых хвойных, но даже от некоторых современных форм. Можно было бы легко представить себе археоптерис в качестве исходной формы, если бы среди голосеменных не было семейств с гораздо более примитивной организацией. Конечно, можно было бы допустить, что хвойные и родствепные им группы имеют самостоятельное происхождение, и тогда их происхождение от растений типа археоптериса не вызвало бы сомнений. Но все говорит о том, что голосеменные представляют собой естественную группу и имеют, следовательно, общее происхождение. Это доказывается не только строением такого сложного органа, как семязачаток, но также строением вегетативных органов. Так, по мнению Ч. Бека (1970), строение стелы семенного папоротника лигиноптериса (Lyginopteris oldhamia) во всех основных чертах, включая явное отсутствие листовых лакун, идентично со строением стелы хвойных. Но если голосеменные действительно имели общее происхождение, то они могли произойти только от предка с протостелической организацией стелы (как у некоторых древнейших семенных папоротников), со слабо развитой вторичной ксилемой и с лестничными трахеидами (сохранившимися даже у некоторых ныне живущих голосеменных). Ни археоптерис и ни аневрофит с их точечными трахеидами не могли быть такой предковой формой. Скорее всего это были какие-то более древние и примитивные протостелические, но в то же время разноспоровые папоротниковидные с лестничными трахеидами во вторичной ксилеме.
Таким образом, мы неизбежно приходим к выводу, что голосеменные произошли не от настоящих (типичных) папоротников, но от одной из боковых разносноровых ветвей древнейших папоротниковидных растений. Это еще раз подтверждает сформулированное известным палеонтологом Э. Коном (1904) правило, согласно которому в процессе эволюции новые группы организмов обычно происходят не от высших представителей предкового таксона, а от сравнительно наименее специализированных его представителей.
Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров . 1974 .
Свидетельство и скидка на обучение каждому участнику
Зарегистрироваться 15–17 марта 2022 г.
Описание презентации по отдельным слайдам:
Голосеменные, или Acrogymnospermae — группа родственных растений, имеющих общих предков, схожих в характере размножения , строении семязачатка, мужского и женского гаметофитов, опыления и оплодотворения. Именно семена отличают Голосеменные растения от и других споровых.
Класс Семенные папоротники
Класс Беннеттитовые
5
Систематика голосеменных
Вымершие
Живущие
Класс Саговниковые
Класс Гинкговые
Класс Гнетовые
Класс Хвойные
Происхождение Голосеменных:
Голосеменные появились в девонском периоде палеозойской эры и были первыми на Земле семенными растениями. Полагают, что именно с ними связано вымирание менее организованных папоротникообразных, размножение которых всецело зависело от присутствия воды. Наибольшего расцвета голосеменные достигли в мезозойской эре, где они доминировали в растительных сообществах. Появление и широкое распространение значительно более прогрессивных цветковых прервало гегемонию голосеменных и привело к вымиранию большинства из них.
Возникновению голосеменных растений предшествовало несколько важных эволюционных событий, которые произошли в течение девонского периода.
В позднем девоне появляются растения с еще более ясно выраженной разноспоровостью. В области вегетативной сферы это было возникновение камбия и появление древесных форм. Хорошо сохранившаяся вторичная ксилема, свидетельствующая о камбиальной активности, также известна уже из отложений среднего девона (эйфельский век).
К концу среднего девона появляется другое древесное растение - знаменитый археоптерис (Archaeopteris latifolia) - крупное дерево, вероятно, высотой более 30 м, с кроной ветвей, несущих спирально расположенные листья (рис. 94). Таким образом появились необходимые предпосылки для возникновения голосеменных. Это важнейшее в эволюции растительного мира событие произошло уже в девонском периоде. Лучшим доказательством явилась находка в 1968 г. Дж. Петтитом и Ч. Беком в верхнедевонских отложениях Северной Америки хорошо сохранившегося семени, названного археоспермой (Archaeosperma).
Спорофитное
(неполовое поколение, На спорофите образуются споры, которые прорастают и дают начало гаметофиту)
Гаметофитное
(представлено
мохообразными, это половое поколение, на котором образуются половые органы — антеридии и архегонии)
Направление эволюции Голосеменных:
Появление семени – крупнейший ароморфоз
Наличие запасных веществ в семени
Все фазы развития на спорофите
Имеется диф-ный зародыш
Наличие семенной кожуры
Эволюция семенных растений
Соотношение гаметофитов и спорофитов растений
Эволюции семян предшествовала эволюция сосудистого камбия и сосудистых тканей. В некоторых случаях камбий возникал самостоятельно (древние плауновидные и хвощевидные), клетки в нём могли подвергаться радиальному продольному делению, что позволяло ткани камбия нарастать в окружности по мере накопления древесины. В отличие от хвощей и плаунов, новый тип камбия смог и до сих пор приводит к массивности деревьев, таких как гикори, секвойя, дуб. Видимо, этот камбий возник только один раз, в группе растений, которая дала начало всем древесным породам.
Цикл размножения голосеменных:
1 – женская шишка; 2 – семенная чешуя с двумя семязачатками; 3 – ; 4 – мегаспоры; 5 – нуцеллус семязачатка; 6 – интегумент, один листочек; 7 – нуцеллус, образующий перисперм; 8 – архегоний; 9 – первичный ; 10 – мужская шишка; 11 – микроспорофилл с двумя микроспорангиями; 12 – антеридиальная клетка; 13 – сифоногенная клетка, клетка трубки; 14 – ядро спермагенной клетки; 15 – ядро клетки-ножки; 16 – клетка трубки; 17 – зародыш семени; 18 – первичный эндосперм; 19 – семенная кожура; 20 – шишка, которой один год; 21 – шишка, возраст которой – два года.
Более совершенные циклы развития имеют сосновые, у которых мужские гаметы без жгутиков (спермии) и пыльцевая трубка доставляет их к яйцеклетке, расположенной в сохраняющихся еще архегониях. Семя начинает развиваться только после оплодотворения и имеет период покоя. Эндосперм семени также образован гаплоидной тканью женского гаметофита.
Эволюция мегаспорангиев выразилась в формировании семяпочки, где собственно спорангием является нуцеллус, но микроспоры улавливает интегумент с пыльцевходом на верхушке, где выделяется капля липкой жидкости. По этому проходу пыльцевая трубка доставляет к архегониям мужские половые клетки.
С появлением пыльцевой трубки и интегумента с микропиле развитие гаметофита и сам половой процесс стали протекать в закрытом виде, ни одна гаплоидная клетка мужского и женского гаметофитов с внешней средой не сообщается. Именно эти 2 структуры создали условия для появления семени. Также не сообщается с внешней средой до полного своего развития и зародыш спорофита.
Сближение процессов бесполого и полового размножения становятся еще ярче выраженным: формирование микро- и мегаспор представляет собой лишь начальные этапы полового размножения.
Семенные папоротники — полностью вымершие растения. Они существовали с позднего девона до раннего мела. Сами голосеменные пришли на смену вымирающим древовидным папоротникообразным и семенным папоротникам.

Если вас не устраивает ответ или его нет, то попробуйте воспользоваться поиском на сайте и найти похожие ответы по предмету школьной программы: биология.
На сегодняшний день (06.03.2022) наш сайт содержит 16351 вопросов, по теме: биология. Возможно среди них вы найдете подходящий ответ на свой вопрос.
Поможем написать работу на аналогичную тему
Нажимая на кнопку "Ответить на вопрос", я даю согласие на обработку персональных данных
Последние опубликованные вопросы
![]()

Индивидуальные работы по различным предметам
Сделайте индивидуальный заказ на нашем сервисе. Там эксперты помогают с учебой без посредников Разместите задание – сайт бесплатно отправит его исполнителя, и они предложат цены.
Цены ниже, чем в агентствах и у конкурентов
Вы работаете с экспертами напрямую. Поэтому стоимость работ приятно вас удивит
Бесплатные доработки и консультации
Исполнитель внесет нужные правки в работу по вашему требованию без доплат. Корректировки в максимально короткие сроки
Если работа вас не устроит – мы вернем 100% суммы заказа
Техподдержка 7 дней в неделю
Наши менеджеры всегда на связи и оперативно решат любую проблему
Строгий отбор экспертов
Требуются доработки?
Они включены в стоимость работы
Работы выполняют эксперты в своём деле. Они ценят свою репутацию, поэтому результат выполненной работы гарантирован
Голосеменные растения являются группой растительных организмов, не обладающих специфическими способами защиты семян.
Классификация голосеменных выглядит следующим образом:
- царство — Растения;
- подцарство — Высшие растения;
- отдел — Голосеменные растения.
К голосеменным растениям причисляют:
- гинкговые;
- гнетовые;
- хвойные;
- саговниковые.
Голосеменные — это около тысячи видов, восемьдесят родов и четырнадцать семейств.
У таких растений наблюдаются определенные адаптации, благодаря которым они вышли на новый эволюционный уровень. Это:
- редуцированный гаметофит, находящийся внутри спорофита;
- неподвижность мужских гамет, переносимых при помощи ветра, насекомых и пыльцевых зерен. Их оплодотворение происходит без воды;
- развивающийся внутри семени зародыш получает защиту и систематическую пищу в виде питательных веществ;
- наличие устьиц на эпидерме наземных органов и эпидерме листьев. Это обеспечивает хороший газообмен между растением и атмосферой.
Появление семенного размножения и уменьшение роли гаплоидной стадии развития — важнейшие ароморфозы в ходе эволюции растений.
Развитие семян голосеменных происходит на поверхности спорофилла. Оно защищается в результате смыкания спорофилла.
Когда возникли голосеменные растения
Поздний карбон — а это примерно 319 млн. лет назад — время появления голосеменных. После они распространились по всем широтам, в том числе они стали обосновываться в достаточно сухих местах.
Для голосеменных свойственно размножение при помощи семени, а в их жизненном цикле преобладает диплоидный спорофит. Эта группа растений имеет женские и мужские половые органы (антеридии и архегонии).
Также у них есть еще одна особенность: длительное функционирование камбия, который образует ксилему или древесину, флоэму и луб.
Камбий представляет собой образовательную ткань голосеменного растения, способную дать начало любому типу растительной ткани.
Также у них имеется главный корень, стержневая корневая система, благодаря которой развиваются древесные формы. Для голосеменных также характерная такая жизненная форма как кустарник.
Нарастание побегов у голосеменных осуществляется моноподиально.
Древние папоротниковидные (вымерли в начале каменноугольного периода) — предки первых голосеменных растений. Расцвет голосеменных приходится на мезозойскую эру, которая характеризуется горообразованием, поднятием материков и иссушением климата. Но начиная с середины мелового периода на смену голосеменным постепенно приходят покрытосеменные.
Современные голосеменные растения представляют собой отдел, включающий свыше 700 видов — это растения, завоевавшие всю планету. Распространение голосеменных пришлось и на умеренные широты Северного полушария. Здесь они сформировали лесное пространство — тайгу.
Современные голосеменные включают 4 класса (гинкговые, саговниковые, гнетовые и хвойные). При этом гинкговые — это всего один вид: Ginkgo biloba. Особо стоит выделить саговниковые, являющиеся остатками вымерших групп, которые процветали в мезозое.
А вот у гнетовых довольно неясное происхождение: палеоботанических данных о них сохранилось немного. Зато класс Хвойные представлен широко и полно. Его видовое разнообразие превосходит все остальные группы голосеменных. Хвойные — это свыше 500 растительных видов. Большая часть таких растений — вечнозеленые. Их возраст говорит о древнем происхождении.
Североамериканская сосна — почетный долгожитель.
Эта группа растений находится где-то между папоротниками и покрытосеменными. В отличие от папоротников, у этой группы растений есть семязачаток, а в отличие от покрытосеменных — у них нет плодолистиков.
Листья голосеменных разнообразны по морфологическим и анатомическим признакам. У большинства видов листья игловидные, у некоторых растений листья довольно большие.
Время появления голосеменных точно установить трудно, их предков обычно ищут среди господствовавших в палеозое растений. Возможными их предками многие считают древнейших представителей отдела папоротниковидные. Среди них были разноспоровые древовидные формы с вторичной древесиной — они вполне могли дать начало голосеменным.
Также голосеменные могли появиться из группы высокоорганизованных папоротников девона — из рода археоптерис. Такие растения имеют сердцевину, хорошо развитую вторичную ксилему, а трахеиды характеризуются точечной формой с окаймленными порами. Уровень организации вегетативных органов ничем не отличает археоптериса от некоторых пермокарбоновых хвойных и определенных современных форм.
Некоторые ученые обозначают самостоятельность происхождения хвойных и родственных им видов. Однако поспорить с этой точкой зрения легко, если приводить в качестве аргумента наличие общего плана строения семязачатка у всех голосеменных.
На единство происхождения голосеменных указывают особенности строения вегетативных органов.
Также считают, что голосеменные могли иметь предка, имеющего протостелическую организацию стелы и слабо развитую вторичную ксилему. Для трахеид таких растений, вероятно, была характерна лестничная форма.
Поэтому можно говорить о том, что предками голосеменных были не настоящие папоротники, а одна из боковых разноспоровых ветвей папоротниковидных живых организмов. На логичность и стройность этого факта указывает на то, что в ходе эволюции новые растительные группы происходят не от высшего таксона предков, а от менее специализированных представителей растительной группы.
Читайте также: