
Как работает транспортная система клетки кратко
Обновлено: 06.07.2024
Синтез белка всегда начинается в цитоплазме. Окончание синтеза происходит в цитоплазме либо на шероховатом эндоплазматическом ретикулуме (ШЭР).
Можно условно выделить два пути транспорта белка в клетке:
1. Из цитоплазмы в некоторые органеллы (ядро, пластиды, митохондрии)
2. Большой путь везикулярного транспорта из ШЭР через аппарат Гольджи (АГ) к другим органеллам (лизосомы, пероксисомы) и через секреторные везикулы во внеклеточную среду. Поскольку синтез всех белков начинается в цитоплазме, а конечная локализация каждого белка может быть различна внутри полипептида имеется система сигналов определяющая его транспортный путь. Первичный сигнал определяет путь из цитоплазмы (в ШЭР, в ядро, в митохондрию или в пластиду), вторичный сигнал определяет дальнейшее направление, например, внешняя или внутренняя мембрана митохондрии или матрикс; лизосома, пероксисома или секреторная гранула.
Сигнальные последовательности белков
Сигнальные последовательности имеют длину 3-80 аминокислот узнаются специфическими рецепторами на мембранах различных компартментов клетки.
Сигнальная последовательность ЭР - гидрофобный участок 5-15 аминокислот на N-конце полипептида.
Сигнал митохондриальных белков 20-80 аминокислот состоящий из спирали и торчащих концов - (+)-заряженного и гидрофобного. 5 (+)-заряженных аминокислот для транспортировки в ядро. Пероксисомные белки имеют последовательность на С-конце Ser-Lys-Leu-COOH.
Имеется класс сигнальных последовательностей которые не позволяют белку достигшему определенной локализации транспортироваться дальше. Например, мотив Lys-Asp-Glu-Leu-COOH (KDEL) не позволяет белкам покидать эндоплазматический ретикулум.
Одна из функций гладкого ЭР - удержание кальция готового для выпуска в цитозоль при стимуляции клетки. Кальретикулин - белок удерживающий ионы кальция. Первые 17 аминокислот включают 14 гидрофобных (синие) - сигнальная последовательность для проникновения в ЭР из цитозоля. Последние четыре аминокислоты KDEL удерживают белок в ЭР.
(NH2) MLL S VPLLL G LL G LAVA E PAV Y F K EQ FL DGDG W TSR WI ESKHKSD F GK FVL SSGK F
YGDEEKDKG L QTSQD A R F Y AL S A S F E PF SNKGQT LVV Q F T V KHEQN I DCGGGY V K LFP
NS L DQTD M HGDSEYN IMF G P D I CG P GTKK V H VIF NYKGKN VLI NKD I RCKDDE F TH L YT LIV R P
DNTYE V K I DNSQ V ESGS L EDD W D FLPP KK I KD P D A SK P ED W DER A K I DD P TDSK P
ED W DK P EH IP D P D A KK P ED W DEE M DGE W E PPVI QN P EYKGE W K P RQ I
DN P DYKGT WI H P E I DN P EYS P D P S I Y A YDN F
G VL G L D LW Q V KSGT IF DN FLI TNDE A Y A EE F GNET W G V TK AA EKQ M KDKQDEEQR L
KEEEEDKKRKEEEE A EDKEDDEDKDEDEEDEEDKEEDEEED VP GQ A KDE L (COOH)
]]> сервер для предсказания сигнальных последовательностей белков ]]>
Некоторые белки имеют различные локализации в клетки. Существует несколько путей транспортировки идентичных полипептидов в различные компартменты клетки [Karniely, 2005]:
1. Несколько сигнальных последовательностей в одном полипептиде преднозначенные для разных компартментов. Каталаза А дрожжей имеет две сигнальные последовательности - для митохондрий и пероксисом, причем количество фермента в этих органеллах зависит от состава среды. Некоторые цитохромы имеют два сигнала - митохондриальный и ЭР. Митохондриальный сигнал запускается после посттрансляционного фосфорилирования белка. Известно, что белок-предшественник амилоида болезни Альцгеймера также имеет два сигнала локализации - ЭР и митохондрий.
2. Одна сигнальная последовательность узнается различными рецепторами на поверхности компартментов. Например, некоторые белки митохондрий и хлоропластов имеют общую сигнальную последовательность, которая более гидрофобна чем специфические сигналы.
3. Сигнал может быть блокирован другим белком. Апуриновая/апиримидиновая эндонуклеаза 1 (Apn1) - основной фермент эксцизионной репарации репарации ДНК в ядре и митохондриях. С-конец имеет сигнал ядерной локализации (NLS), за которым идет сигнал митохондриальной локализации. белок Pir1 взаимодействует с С-концом Apn1 блокируя NLS.
4. Сигнал может быть блокирован специфическим сворачиванием белка. Аденилат-киназа дрожжей Aky2 локализуется в цитоплазме и в небольшом количестве в межмембранном пространстве митохондрий, имеет две сигнальные последовательности, активность которых зависит от конформации белка.
5. Сигнал может быть блокирован после модификации полипептида. Фосфорилированный цитохром CYP2B1, взаимодействует с цитозольным шапероном Hsp70, что приводит к конформационным изменениям и переключает одну сигнальную последовательность на другую.
6. Одна РНК может иметь два сайта инициации трансляции при этом образуются два белка - один с сигнальной последовательностью, другой без нее, что определит различную локализацию белков в клетке. В другом случае может образовываться две различные РНК кодирующие два идентичных белка, но у одного будет сигнальная последовательность, а у другого нет.
Транспорт в ядро
Транспорт в митохондрии и пластиды
Митохондрии и пластиды имеют собственную ДНК и самостоятельно синтезируют некоторые белки. Однако многие из основных белков митохондрий и пластид синтезируются в цитозоле.
Белки проникающие в митохондрии должны нести сигнал, определяющий локализацию - внутрення или наружная мембрана, или матрикс.
Белки преднозначенные для матрикса несут сигнал на N-конце, который узнается рецепторами на внешней мембране. Рецептор связан с комплексом переноса белка, который разворачивает белок и переносит его через мембрану. После переноса белка сигнальная последовательность отрезается и белок снова сворачивается.
Белки шапероны связываются с вновь синтезированным белком предотвращая его сворачивание.
Шаперонины связываются с белком после его транспортировки к месту доставки и способствуют правильному сворачиванию.
В ответ на различные стрессовые воздействия (например повышение температуры) в клетке синтезируются шапероны называемые белками теплового шока - hsp (heat-shock proteins), которые стабилизируют клеточные белки. Hsp обнаружены во всех клеточных компартментах эукариот и у бактерий.
Везукулярный транспорт
Из одной органеллы в другую перемещение происходит в везикуле или на ее поверхности в виде интегральных белков.
Донорый компартмент – органелла от которой отрывается мембрана в составе везикулы, акцепторный компартмент – принимает везикулу.
конститутивная секреция – происходит постоянно и не зависит от внешних сигналов.
регулируемая секреция – под ПМ происходит накопление пузырьков, которые сливаются с ПМ при наличии внешних сигналов – гормоны, нервы – и повышении конц. Ca2+ до 1мкм
ретроградный транспорт – возвращение рецепторных белков и липидов из АГ в Эр - восполнение мембраны ЭР.
антероградный транспорт – растворимые грузовые белки двигаются по секреторному пути ЭР→ пузырек?цис-Гольджи?пузырек?транс-Гольджи?пузырек?органелла или секреция
Окаймленные везикулы - покрыты белками, кот узнают и концентрируют специфич. м-ные белки и отделяют м-ну пузырька, формируют решетку и придают форму везикуле: клатриновые, COPI, COPII:
Клатриновые везикулы – ~0,1мкм, транспорт из АГ и ПМ,клатрин - 3типа, 3 большие и 3 малые субъединицы формирующие трискелетон – собирающиеся на поверхности м-ны со стороны цитоплазмы в пента- и гексагоны, кот спонтанно формируют сферу. Адаптин – связывает клатрин с м-ной и ловит различные трансм-ные белки в том числе грузовые рецепторы, кот. захватывают р-римые грузовые белки, кот попадают внутрь везикулы. Имеетя по крайней мере 4 типа адаптинов
динамин - GTP-аза, р-римый цитоплазматический белок, образует кольцо на отделяющейся клатриновой везикуле – регулирует кол-во клатрина отщепляющееся вместе с м-ной в составе везикулы, ассоциирует другие белки помогающие выпучить м-ну и белки модификаторы липидов, изменяющие локально липидный состав м-ны для выпучивания
После отделения везикулы от м-ны клатрин и адипин отделяют шапероны - ATP-азы hsp70 семейства. Ауксилин – прикрепляется к везикуле и активирует АТФ-азу. Т.к кайма формирующейся везикулы сущ. дольше чем кайма отделенной – имеется стабилизирующий механизм. Клатриновая оболочка обеспечивает значительную силу для изгибания м-ны, т.к. везикулы из внутриклеточных компартментов образуются на уже выпученной м-не
COP-I – транспорт от АГ и ЭР, 8субъединиц, GTP-белок – фактор рибозилирования АДФ –ARF – транспорт
COP-II – транспорт из АГ и ЭР, 5 субъединиц
Везикулы мб не только сферические, часто образуются трубчатые везикулы в которых высокое соотношение S/V
Образование клатриновых и COP везикул регулируется GTP-связывающими белками, которые могут находится в активном GTP- и неактивном GDP-состоянии
Два класса белков обменивают GDP-GTP: GEF-гуанин-нуклеотид-фактор обмена активирует белки заменяя GDF?GTF, GAP- белок активирующий GTP-азы – инактивирует GTP-связывающие белки меняя GTP?GDP.
GTP-азы необходимые для сборки окаймленных везикул перед сборкой пузырьков: мономерные GTP-связывающие белки (GTP-азы):
ARF-белки – необх для клатриновой и COP сборки на пов-ти м-ны АГ. Sar1 белок, необходим для COPII сборки на на ЭР м-не
тримерные (G белки).
GTP-азы находятся в цитозоле в неактивном состоянии, перед сборкой GEF встраивается в м-ну ЭР и связывает цитозольный SarI, кот обменивает GDF?GTP. В GTP состоянии SarI встраивается остатком жирной к-ты в м-ну ЭР. Ассоциирует белки об-ки и инициирует отпочковывание везикулы. GTP-азы попавшие в м-ну активируют фосфолипазу D, кот преобразует фосфолипиды в фосфотидную к-ту, что усиливает связывание оболочных белков. Вместе белок-белковые и белок-липидные взаимодействия изгибают м-ну
SNARE – белки – отвечают за слияние донорной и акцепторной м-н, более 20, каждая на специфич пов-ти м-ны, трансмембранные белки на пов-ти везикулы - v-SNAR, на пов-ти донора – t-SNAR. Взаимодействуя v- и t-SNAR обвиваются др на друга в транс-SNAR-комплекс, обеспечивающий слияние м-н. SNF-белок разрушает транс-SNAR-комплексы – цитозольный шаперон ATP-аза, использует адаптирующие белки для связывания с комплексом-SNAR
Rab-белки – мономерные GTP-азы, более 30, каждая органелла имеет хотя бы один Rab на м-не со стороны цитоплазмы, регулируют стыковку везикул и связывание v-SNAR-ов и t-SNAR-ов необходимых для слияния м-н. В состоянии GDP-не активны, нах в цитозоле, в состоянии GTP-активны и переходят на пов-ть м-ны органеллы или везикулы. В активном состоянии Rap связываются с м-ной липидным якорем и собирают другие белки участвующие в слиянии м-н
неактивный Rab-GDP связан с GDI – GDP-диссоциирующий ингибитор. Rab-GDP связывается с GEF-гуанин нуклеотид меняющий фактор, связанный с м-ной донорного компартмента – меняет GDP на GTP. Rab-GTP связывается с м-ной формирующейся везикулы и ассоциирует v-SNARE, которые в составе везикулы транспортируются к органелле и связываются с Rab-эффекторами и t-SNARE, связанными с м-ной акцепторного компартмента и обеспечивают слияние м-н
белок органелла
Rab1 ЭР и АГ
Rab2 цис-АГ
Rab3A синаптич везикулы, секрет гранулы
Rab4 ранние эндосомы
Rab5A ПМ, клатриновые везикулы
Rab5C ранние эндосомы
Rab6 промежуточный- и транс-АГ
Rab7 поздние эндосомы
Rab8 секреторные везикулы (базолатеральные)
Rab9 поздние эндосомы, trans-АГ
Слияние м-н происходит не только при везикулярном транспорте: слияние спермия с яйцом, слияние миобластов во время развития мышечной клетки.
Образование клатринового пузырька. Диаметр клатринового пузырька ~0,3 мкм
ТРАНСПОРТ ЧЕРЕЗ МЕМБРАНУ МАЛЫХ МОЛЕКУЛ
унипорт-независимый перенос, симпорт-одновременный, однонаправленный перенос, антипорт-одновременно антинаправленный транспорт | симпорт и антипорт - контранспорт - скорость процесса зависит от двух переносимых в-в
пассивный транспорт - идущий по градиенту концентрации и/или по электрическому градиенту - простая диффузия, канальный транспорт, облегченная диффузия
простая диффузия - липидная м-на обладает малой проницаемостью для ионов и большинства полярных молекул (исключение вода). Неполярные диффундируют Н2О, CO2, O2. Коэффициенты проницаемости для низкомолекулярных соединений коррелируют с отношением их растворимости в неполярных растворителях к растворимости в воде. Эта зависимость дает основание думать, что низкомолекулярные соединения проходят сквозь двуслойную липидную мембрану следующим образом: сначала они теряют окружающую их гидратную оболочку, затем растворяются в углеводородном внутреннем слое мембраны, после чего диффундируют через этот внутренний слой к другой стороне мембраны, где вновь растворяются в воде.
каналы - трансм-ные белки формирующие в м-не сквозные проходы - индиферентные (всегда открыты - каналы чувствительные к давлению), аллостерические (химически регулируемые - при наличии лиганда никотиновый ацетилхолиновый рец-р, рец-р -аминомаслянной к-ты, рец-р глицина) и потенциал-зависимые (при изменении м-ного потенциала Na+-канал, K+-канал, Ca2+-канал)
облегченная диффузия осущ белками переносчиками, кот специфич связывают и переносят молекулы через липидный бислой, путем изменения конформации
активный транспорт - требующий затрат энергии:
первично-активные -непосредственно использует энергию- сопряженные с ох-red р-циями (цитохром с-оксидаза), сопряженные с поглощ. hv (бактериородопсин), АТФ-азы (Na+/K+-АТФ-аза, Ca2+-АТФ-аза, Н+-АТФ-аза, K+-АТФ-аза, H+/K+-АТФ-аза
вторичноактивные по градиенту, созданному в процессе первично-активного транспорта - контранспорты – симпортеры (лактозопермеаза), антипортеры (белок полосы 3)
транспорт некоторых сахаров и аминокислот внутрь животных клеток обусловливается градиентом Na+ через плазматическую мембрану. Всасывание глюкозы в клетки кишечника и почек достигается с помощью системы симпорта, в которой глюкоза и ионы Na+ связываются с различными участками на белке-переносчике глюкозы (рис. 56). Na+ стремится войти в клетку по своему электрохимическому градиенту и как бы "тащит" глюкозу внутрь за собой. Чем выше градиент Na+, тем больше скорость всасывания
глюкозы. И наоборот, если концентрация Na+ во внеклеточной жидкости заметно уменьшается, транспорт глюкозы останавливается. Ионы Na+, проникающие в клетку вместе с глюкозой, выкачиваются обратно Na+,К+-АТФазой, которая, поддерживая градиент концентрации Na+, косвенным путем контролирует транспорт глюкозы. В плазматических мембранах многих животных клеток существует, по крайней мере, 5 различных белков переносчиков аминокислот, которые действуют как системы симпорта, перенося одновременно ионы Na+, причем каждый из этих белков специфичен для группы родственных аминокислот. У бактерий большинство систем активного транспорта, приводящихся в действие ионными градиентами, использует в качестве котранспортируемого иона Н+, а не Na+. В частности, активный транспорт большей части сахаров и аминокислот в бактериальные клетки обусловлен градиентом Н+ через плазматическую мембрану. Наиболее хорошо изученный пример такого рода - это трансмембранный белок переносчик лактозы (пермеаза, или М-белок), состоящий из одной полипептидной цепи (30 кДа) и осуществляющий Н+-зависимый симпорт: с каждой транспортируемой в клетку молекулой лактозы переносится один протон.
векторный транспорт - способ активного транспорта - молекулы, проникшие в клетку пассивно, химически модифицируются с затратой энергии, так что модифицированные молекулы уже не могут выйти обратно через тот же канал. Встречается у бактерий при транспорте сахаров: при переносе через плазматическую мембрану сахара фосфорилируются, становятся заряженными и не могут выйти обратно, поэтому они начинают накапливаться в клетке. При этом концентрация нефосфорилированных сахаров внутри клетки остается очень низкой, и сахара продолжают поступать в клетку по градиенту концентрации. Механизм сопряженного с транспортом фосфорилирования сахаров у бактерий довольно сложен: в нем участвуют, по крайней мере, четыре разных мембранных белка, а в качестве донора фосфатной группы используется не АТФ, а фосфоенолпируват
ТРАНСПОРТ ЧЕРЕЗ МЕМБРАНЫ МАКРОМОЛЕКУЛ
ТРАНСПОРТ БЕЛКОВ через мембраны
пориновый комплекс – 125000кДа транспорт ч-з ядерную м-ну цитозольных белков, сигнальные посл-ти связ с пк и переносятся с сохранением третичной стр-ры
белки-переносчик – транспорт из цитоплазмы в органеллы, белковые комплекся переносятлинейные пептиды ч-з биомембраны энергозависимым образом посредством связывания сигнальной посл-ти с рецептором, разворачивание и сворачивание белка контролируется шаперонами
• Существует два основных типа мембранных транспортных белков: каналы и переносчики
• Ионные каналы катализируют быстрый и избирательный транспорт ионов по электрохимическому градиенту
• Транспортеры и насосы представляют собой белки-переносчики, которым необходима энергия для транспорта веществ против электрохимического градиента
• В каждой клетке несколько различных мембранных транспортных белков действуют как единое целое
Некоторые из транспортных белков находятся в плазматической мембране, в то время как другие входят в состав мембраны клеточных органелл. Для поддержания определенного состава внутриклеточной среды и клеточных компартментов необходимо, чтобы транспортные белки обладали избирательностью по отношению к определенным метаболитам. В зависимости от характера процесса транспорта мембранные транспортные белки подразделяются на две группы: каналы и переносчики.
Канальные белки содержат поры, через которые при открытом канале с высокой скоростью проходят метаболиты. Белок-переносчик связывает метаболит, находящийся по одну сторону мембраны, после чего конформация его меняется, и метаболит высвобождается с другой стороны мембраны.
Существует несколько типов белков, образующих каналы в мембранах. Порины, присутствующие в клетках некоторых прокариот, в митохондриях и в нексусах, где соединяется цитоплазма соседних клеток, обеспечивают прохождение метаболитов в соответствии с размером их молекул.
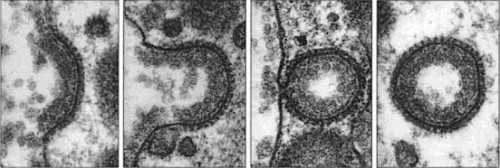
К более селективным каналам относятся комплексы ядерных пор и каналы, обеспечивающие перемещение белков через мембрану эндоплазматического ретикулума. В статьях на сайте будут рассмотрены ионные каналы и аквапорины, через которые осуществляется селективный трансмембранный перенос ионов и молекул воды соответственно. К настоящему времени описано более 100 различных типов канальных белков. Все они обладают следующими характеристиками:
• селективностью по отношению к переносимому метаболиту,
• высокой скоростью переноса метаболита и
• воротным механизмом, регулирующим транспорт метаболитов.
Участок канального белка, через который осуществляется трансмембранный транспорт, называется порой канала. Канал может находиться в нескольких различных конфигурациях. Некоторые каналы состоят из одного белка, трансмембранные сегменты которого образуют пору. Другие существуют в виде олигомеров, состоящих из идентичных или разных субъединиц, которые вместе образуют пору.
Наконец существуют каналы, состоящие из двух и более субъединиц, каждая из которых сама образует пору. Часто комплексы олигомеров каналов регулируются или позиционируются на определенных мембранах с участием дополнительных субъединиц.
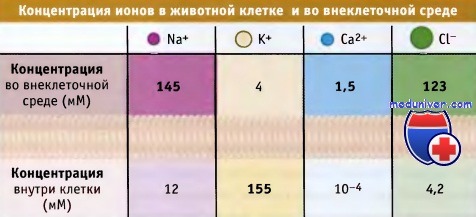
Большинство канальных белков обладают высокой селективностью в отношении определенных метаболитов, например ионов натрия (Na+), калия (К+), кальция (Са2+), хлорида (Сl-) или воды. Другие канальные белки являются неселективными катионными или анионными каналами. Как будет показано далее, при рассмотрении свойств специфических канальных белков, их поры характеризуются определенными структурными особенностями, т. е. являются селективным фильтром, который позволяет им проявлять избирательность по отношению к различным метаболитам.
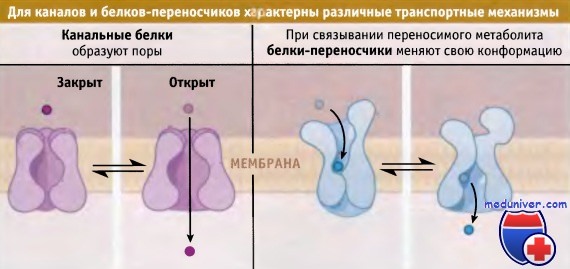
Каналы и переносчики представляют собой два основных типа белков мембранного транспорта.
Растворенные метаболиты с максимальной скоростью диффундируют через пору каналов,
а переносчики связывают метаболит с одной стороны мембраны, затем претерпевают конформационные изменения и высвобождают его с другой стороны.
Этот процесс протекает значительно медленнее.
Направление потока ионов растворенных веществ через канал определяется электрохимическим градиентом. Иными словами, метаболиты движутся через канал в энергетически-выгодном направлении, т. е. по электрохимическому градиенту. Например, для покоящейся клетки, схематически изображенной на рисунке ниже, существует направленный в клетку ионный поток через Na+-, Са2+-, Cl--каналы, и противоположный, проходящий через К+-каналы.
Так как других источников энергии, кроме электрохимического градиента, в данном случае не используется, то такой тип транспорта называется пассивным. Канальные белки обеспечивают высокую скорость транспорта. Так, для ионных каналов скорость составляет 108 в 1 с, что близко к максимальной скорости диффузии ионов в водной среде.
Транспортные мембранные белки контролируют различные клеточные функции при участии воротного механизма. В основе этого механизма лежит индукция конформационных изменений в белке в ответ на воздействие определенного стимула. Эти изменения приводят к открытию или закрытию канала. Например, открытие и закрытие ионных каналов могут регулироваться лигандами, величиной электрического потенциала, механическими или температурными воздействиями. Активация каналов может происходить очень быстро, что делает их идеальным инструментом обработки поступающих в клетку сигналов.
Например, передача нервного импульса зависит от электрического заряда, который создается ионным каналом в мембране. Каналы также играют важную роль в регуляции объема клетки и внутриклеточного pH, в транспорте солей и воды через эпителиальные клетки, в закислении внутриклеточных органелл и во внеклеточной передаче сигналов.
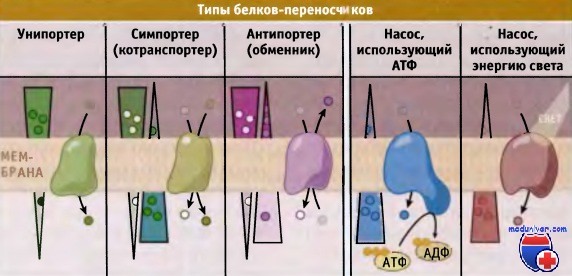
Транспортеры и насосы представляют собой два основных типа белков-переносчиков.
Транспортеры делятся на три типа, каждый из которых осуществляет один из типов переноса: унипорт, симпорт, или антипорт.
Показано направление электрохимического градиента через мембрану.
В зависимости от типа белка-переносчика транспорт может происходить или в направлении градиента (от высокой концентрации к низкой), или против него.
При транспорте субстратов против градиента концентрации, белки-переносчики используют свободную энергию электрохимического градиента, АТФ или других источников. Поскольку при этом происходит потребление энергии, процесс носит название активный транспорт. Белки-переносчики подразделяются на две группы, транспортеры и насосы.
Для транспортировки субстратов через мембрану транспортеры используют энергию электрохимического градиента. Они подразделяются на унипортеры, симпортеры (или котранс-портеры) и антипортеры (обменники). Для энергозависимого транспорта метаболитов мембранные насосы используют энергию, например высвобождающуюся при гидролизе АТФ. По сравнению с канальными белками, переносчики характеризуются более медленной скоростью транспорта, порядка 1000 молекул в 1 с.
Различают два типа активного транспорта, первичный и вторичный. Белки-переносчики, осуществляющие первичный активный транспорт, в качестве источника энергии используют АТФ и транспортируют метаболиты против электрохимического градиента. Таким образом, они помогают поддерживать градиент концентрации метаболитов по обеим сторонам мембран клетки. Са2+-АТФаза и Na+/К+-АТФаза являются характерными примерами транспортных насосов, которые управляют процессом первичного активного транспорта.
Белки-переносчики, которые участвуют во вторичном активном транспорте, непосредственно не используют энергию АТФ. Вместо этого они используют свободную энергию электрохимического градиента, который возникает за счет первичного активного транспорта. Вторичный активный транспорт происходит с участием симпортеров и антипортеров. Механизмы транспорта метаболитов представлены на рисунке ниже.
В клетке все типы мембранных транспортных белков-каналы, транспортеры и насосы-функционируют совместно. Таким образом, работа какого-либо одного типа транспортного белка зависит от работы других. В данной главе мы рассмотрим несколько примеров такого взаимодействия транспортных белков. Например, ионный градиент по обеим сторонам мембраны поддерживается за счет сложных взаимодействий между различными типами транспортных белков. Для правильного функционирования эпителиальных клеток, например почек, слизистой кишечника и легкого, необходимо протекание транспортных процессов с участием различных ионов и метаболитов. Мы также рассмотрим роль нарушений процессов транспорта в развитии различных заболеваний.

Транспорт растворенных метаболитов через мембрану может быть пассивным или активным.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Переносчики и каналы — устройства, обеспечивающие специфические транспортные процессы. В клетках высших организмов, и в том числе в кишечных, существуют транспортеры многих типов. К ним относятся переносчики глюкозы, аминокислот и др. Каждое такое устройство переносит один или ограниченное число типов органических молекул через мембрану по электрохимическому градиенту или благодаря сопряжению с механизмом транспорта другого вещества, движение которого до градиенту концентраций служит источником энергии для сопряженного с ним процесса.
Для вторичной энергизации используются многие ионные градиенты, но преимущественно градиент Na + без участия АТФ (табл. 8). Na + -зависимые переносчики используются в различных системах, включающих в себя систему всасывания (например, тонкая кишка), систему обеспечения собственных энергетических и пластических потребностей, транспорт ряда веществ (например, тех же глюкозы и аминокислот) для собственных синтезов и т.д. В печени механизмы активного транспорта глюкозы используются для депондрования веществ и их иммобилизации из депо. Мобильный переносчик, представляющий собой, как правило, белковую молекулу, движется от одной поверхности мембраны к другой, совершая вертикальные или вращательные движения, с тем чтобы связывать транспортируемые субстраты на одной поверхности мембраны и освобождать с другой. Типичным примером такого переносчика служат ионофоры. Канал как устройство характеризуется наличием постоянной или индуцированной поры, через которую проходит транспортируемое вещество.
Nа + -зависимый транспорт органических веществ в клетках животных
Ткани и клетки, для которых характерен Na + -завксимын транспорт аминокислот
Эпителиальные ткани Тонкая кишка (животные различных видов) Почки Желчный пузырь (кролик) Мочевой пузырь земноводных Стенка тела (морские беспозвоночные) Неэпителиальные ткани Жировые ткани Хрусталик глаза Кости и хрящи Печень Мозг Поперечнополосатые мышцы Одиночные клетки Асцитная карцинома Эрлиха Фибробласты мыши Морские бактерии Ретикулоциты (кролик) Эритроциты голубя Лейкоциты (человек)
Вместе с тем, как упомянуто выше, наряду с Na + -зависимым транспортом существует Ма + -пезависимый транспорт. Этот тип транспорта обнаружен при изучении переноса через мембрану кишечных клеток моносахаридов, аминокислот и других мономеров, образующихся в результате мембранного гидролиза соответствующих олигомеров.
Таблица 10
Некоторые примеры генетических нарушений транспортных систем у человека
Благодаря распространению Na + -зависимых транспортеров на апикальной мембране, а натриевых насосов — на базолатеральной создается транзитный перенос, необходимый для обеспечения за счет работы каждой кишечной клетки потребностей многих тысяч других соматических клеток. Транспортные функции клеток других типов реализуются практически тем же самым набором функциональных блоков. Действительно, транспортные характеристики клеток тонкой кишки и плаценты во многом совпадают.
Ряд клеток характеризуется тем, что универсальные транспортные блоки расположены на клеточной поверхности иначе. В частности, в ацинарных клетках поджелудочной железы, осуществляющих внешнюю секрецию ферментов, насосы и Na + -зависимые транспортеры расположены в базолатеральной мембране. Этого достаточно для обеспечения градиента Na + между вне- и внутриклеточной жидкостями и поступлением нутриентов из крови в цитоплазму, а также для энергизации многих других процессов. Кроме того, локализация насосов и каналов в базолатеральной мембране исключает интенсивное поступление потоков воды и солей в апикально-базальном направлении.
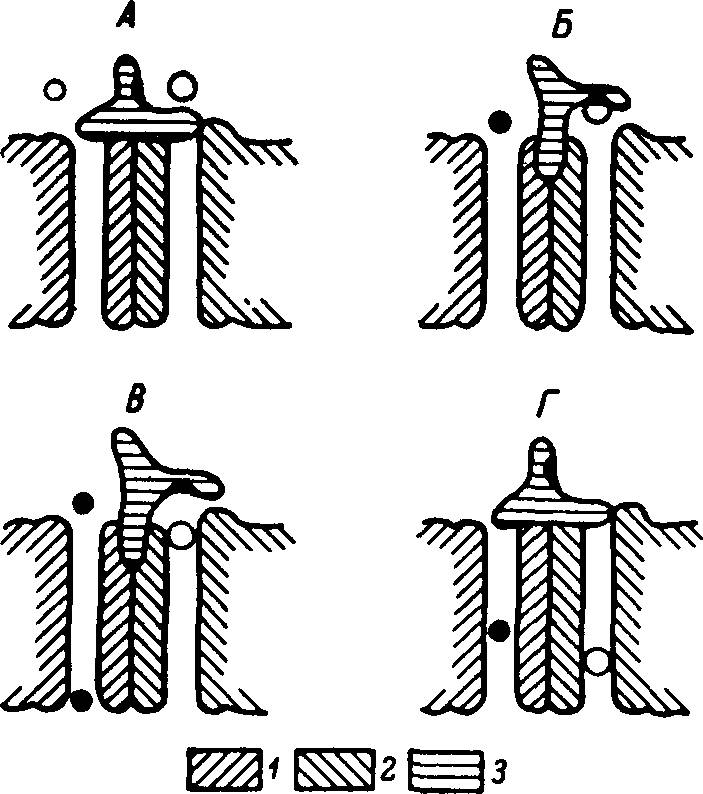
Рис. 28. Схема действия транспортера глюкозы и натрия в плазматической мембране.
А — исходное состояние транспортера: оба канала закрыты; Б — после связывания глюкозы с аллостерическим центром на воротном устройстве: натриевый канал открылся; В — при прохождении натрия по натриевому каналу произошла аллостерическая активация глюкозного канала и молекула глюкозы перешла с воротного устройства на вход своего канала; Г — освобождение центра на воротном устройстве влечет за собой закрытие натриевого и глюкозного каналов. 1 — натриевый канал; 2 — глюкозный канал; 3 — воротное устройство. Светлые пружки — молекулы глюкозы, черные — ионы натрия.
Согласно современным представлениям, при Na + -зависимом транспорте глюкозы через плазматическую мембрану клеток эукариотов происходит образование тройного комплекса: Na + —глюкоза—транспортер. Однако более адекватной представляется наша модель транспортера (рис. 28). Она характеризуется наличием двух параллельно взаимодействующих каналов (для Na + и для глюкозы) и поверхностного (воротного) белка, связывающего глюкозу на входе в транспортную систему. Транспортный цикл такой модели состоит в следующем: 1) исходно глюкозный и натриевый каналы неактивны; 2) при связывании глюкозы с аллостерическим центром на воротном белке натриевый канал активируется (эта стадия напоминает контроль натриевой проницаемости ацетилхолином) и Na + движется из экстра- в интрацеллюлярную жидкость; 3) на определенном этапе движения Na + по каналу происходит аллостерическая активация глюкозного канала; 4) через активированный канал транспортируется молекула глюкозы, первоначально фиксированная в воротном устройстве; 5) освобождение воротного устройства сопровождается дезактивацией натриевого канала. Реактивация последнего происходит при связывании с аллостерическим центром следующей молекулы глюкозы. При температуре 16° С глюкозный канал инактивируется, а натриевый продолжает функционировать. Из нашей модели вытекает, что скорость транспорта глюкозы и Na + в определенном диапазоне зависит от концентрации Na + во внеклеточной жидкости. Важно, что Ка + -зависимые транспортеры глюкозы и аминокислот, поскольку показана их идентичность в клетках различных типов и у организмов разных групп, должны соответствовать одной из моделей молекулярных машин.
Несмотря на то что сравнительная физиология ионных каналов изучена недостаточно, существуют данные о значительном сходстве их молекулярной структуры и функциональной организации. Вероятно, у млекопитающих и насекомых имеются все основные типы ионных каналов — натриевые, калиевые, хлорные и др.
Далее, у млекопитающих, рыб и насекомых продемонстрировано наличие специализированных натриевых каналов, чувствительных к амилориду, а также каналов, реализующих котранспорт Na + и Сl - , чувствительных к фуросемиду. Интересно, что те же самые ингибиторы специфически тормозят транспорт ионов в мальпигиевых канальцах и толстой кишке насекомых. Недавно выявлен котранспортер, осуществляющий котранспорт в клетку одного иона Na + , одного иона К + и двух ионов Сl - . Этот механизм обнаружен в клетках почки и тонкой кишки млекопитающих, в клетках различных органов рыб и насекомых.
До недавнего времени считалось, что функции секреции, экскреции и всасывания имеют глубокие различия, касающиеся механизмов функционирования и эволюционного происхождения. Однако между этими процессами принципиальной разницы не существует. Все основные функциональные блоки, упомянутые выше, лежат и в основе перечисленных функций, в ряде случаев полярных по своему физиологическому значению (например, всасывание и экскреция).
Характеристики ферментных и транспортных систем клеток, выполняющих различные функции (например, всасывание в тонкой кишке, реабсорбция воды в протоках пищеварительных желез и т.д.), очень близки. В апикальной мембране клеток тонкой кишки и почечных канальцев присутствуют трансмембранные ферменты, Ка + -зависимые транспортеры глюкозы, аминокислот и других нутриентов. В базолатеральной мембране этих клеток локализованы близкие или идентичные по молекулярной структуре АТФазы. Системы с множеством транспортеров, локализованных в апикальной мембране, и натриевых насосов, расположенных в базолатеральной мембране, распространены очень широко. В некоторых структурах они выполняют не только резорбтивную, но и секреторную функцию.
Существенно, что при электронной микроскопии многие субклеточные и молекулярные структуры клеток тонкой кишки человека и высших животных неотличимы от таковых клеток кишки рыб, насекомых, моллюсков, круглых червей и т.д., а клетки тонкой кишки — от клеток плаценты, канальцев почек и других клеток.
1. Детерминанты половых клеток
1. Детерминанты половых клеток Будущие половые клетки — гоноциты иногда можно отличить уже на очень ранних стадиях развития. Классическим примером является аскарида, у которой после первого деления яйца в одном из двух бластомеров теряются концевые участки хромосом и
2. Пол половых клеток
2. Пол половых клеток После того как гоноциты попали в половые железы, их судьба зависит от того, какие половые клетки они должны образовать — мужские, т. е. сперматозоиды, или женские, т. е. яйца. В первом случае их путь развития называют сперматогенезом, а во втором —
1. Форма клеток
1. Форма клеток Форма клеток зависит от их внутренней структуры и свойств клеточной оболочки и от их окружения — соседних клеток и поверхностей контакта. Так, при культивировании отдельных клеток на поверхности стекла все клетки стремятся распластаться по субстрату.
3. Перемещения отдельных клеток
3. Перемещения отдельных клеток Многие процессы формообразования происходят не путем перемещения клеточных пластов, а путем перемещения отдельных клеток. Иногда движение клеточного пласта происходит при частичной потере его эпителиальной структуры. Так, например,
Два главных органа иммунной системы вырабатывают два сорта клеток: Т–и В–лимфоциты.
Два главных органа иммунной системы вырабатывают два сорта клеток: Т–и В–лимфоциты. — Следовательно, иммунная система каждого организма имеет два главных органа, не так ли? — Так? — Один, тимус, заведует выработкой лимфоцитов, способных превращаться в
§ 8. Заряды мембраны нервных клеток
§ 8. Заряды мембраны нервных клеток Однако основные свойства нервной системы обусловлены способностью быстро реагировать на изменение ситуации внутри или вне организма. Скоростные процессы не могут осуществляться по медленным гуморальным законам, они происходят по
7.8. Сигналы нервных клеток
7.8. Сигналы нервных клеток Нервная система возникает в ходе эволюции для анализа организмами поступающей информации. С этой целью нервные клетки используют электрические и химические сигналы. Электрические сигналы бывают двух видов.Градуальный потенциал возникает в
5.3. Транспортные системы клеток. Насосы
5.3. Транспортные системы клеток. Насосы Под насосами подразумеваются системы, в которых сочетаются механизмы энергизации и трансмембранного переноса. Принимается, что источником энергии в большинстве случаев служит энергия макроэргических связей АТФ. Деятельность
2.7. Попытки моделирования примитивных клеток
2.7. Попытки моделирования примитивных клеток В ранних работах о происхождении жизни была выдвинута идея предклеточных структур, существующих в водной среде в форме “микросфер” (Fox, 1965; 1991) и “коацерватных капель” (Опарин, 1966). Микросферы формировались из т. н.
9.3. Возникновение клеток
9.3. Возникновение клеток Предполагается, что этапом возникновения жизни на Земле следует считать период, когда сформировались простейшие клеточные системы, ставшие элементарной ячейкой живого. Сведения, касающиеся этой проблемы, освещены в обзорах, посвященных
9.3. Возникновение клеток
9.3. Возникновение клеток Предполагается, что этапом возникновения жизни на Земле следует считать период, когда сформировались простейшие клеточные системы, ставшие элементарной ячейкой живого. Сведения, касающиеся этой проблемы, освещены в обзорах, посвященных
Изучение динамических процессов, происходящих в клетке во время жизни, является по-прежнему одной из наиболее трудных и увлекательных областей исследований современной науки. Она содержит множество тайн, и каждая раскрытая тайна спасает сотни тысяч жизней, поскольку дает ключ к созданию уникальных способов сохранения здоровья и улучшения самочувствия человека. Именно поэтому над раскрытием тайн природы, заложенных в клетке, сегодня трудятся не только биологи и генетики, но и биофизики, биохимики, системные аналитики, специалисты в области информатики, лингвистики и других областей знаний.
Клетка является сложной физической системой. В один и тот же момент времени в ней происходят десятки тысяч разнообразных динамических процессов. Для их изучения ученые используют модели. Модель клетки должна отражать свойства и функции живой клетки. Например, свойство клетки приспосабливаться при изменении параметров внешней среды (температуры, давления, влажности воздуха и других). Однако ни в какой модели нельзя учесть все свойства реальной клетки. Даже если бы нам и удалось встроить в модель значительную часть этих свойств, то задача получилась бы столь сложной, что решение ее было бы чрезвычайно затруднено или даже вовсе невозможно.
Тем не менее, ученые постоянно трудятся над созданием различных моделей, обладающих свойствами живой системы. По совокупности процессов, происходящих в клетке, ее можно сравнить с биороботом, наделенным, с точки зрения современной науки, фантастически совершенными свойствами: самовоспроизведения, самообучения и самонастройки.
В технике робот представляет собой информационно-вычислительный комплекс. Систему его функционирования можно разделить условно на пять основных подсистем: техническую, программную, лингвистическую, информационную и организационную. В свою очередь данные подсистемы можно разделить на два класса. Первый класс включает техническую подсистему, представляющую собой материальные средства комплекса (специалисты называют их “железом”), и второй класс – остальные четыре подсистемы, отвечающие за организацию информационного процесса.
Аналогом технической подсистемы в клетке являются ее биологическая субстанция, имеющая определенную форму и строение. В качестве строительного материала в ней используются органические вещества (биополимеры). Подробно об этом написано в разделе 1.3.2. Второй класс объединяет подсистемы, отвечающие за организацию динамических процессов – информационную жизнь клетки. Устройство клетки является настолько сложным, что воспроизвести искусственно подобную ей систему не по силам ни одной лаборатории мира.
В последние полвека ученые создали немало моделей разных искусственных систем: самолетов, ядерных реакторов, роботов. Более сложным оказалось моделирование природных явлений. Одним из таких примеров является моделирование процессов, позволяющее предсказывать погоду. Опыт, накопленный при проведении таких работ, позволил разработать общую теорию систем, обобщающую и раскрывающую фундаментальные свойства сложных объектов.
Для упрощения понимания протекания внутриклеточных процессов используем разные подходы к рассмотрению динамических процессов (биофизических, биохимических, энергетических, информационных). При этом мы будем вынуждены в большей или меньшей степени идеализировать свойства описываемой системы, учитывая только те решающие факторы, которые определяют черты поведения, обусловленные конкретным видом динамических процессов. Данный подход к рассмотрению вопроса позволит представить нам общие свойства клеток, тканей, органов или систем органов, организма в целом как системы.
Клетка является сложной открытой динамической системой, содержащей множество входов и выходов (смотри рисунок 1.4.1).
Рисунок 1.4.1. Системная модель клетки. Общие входы и выходы
В процессе жизнедеятельности клетка выполняет две основные задачи: обеспечивает поддержание стабильности жизнеобеспечения клеточной системы и реализует специфические функции, присущие определенному виду клеток (смотри рисунок 1.4.2).
Рисунок 1.4.2. Системная модель клетки. Разделение функций клетки
Основные функции клетки =
поддержание стабильности
подсистемы жизнеобеспечения +
выполнение специфических функций
Поддержание стабильности подсистемы жизнеобеспечения происходит за счет выработки энергии, трансмембранного переноса вещества, синтеза клеточных и тканевых структур, размножения клеток.
Выработка необходимой для жизни клетки и организма в целом энергии происходит в процессе протекания процессов распада клеточных и тканевых структур (катаболизма), а также сложных соединений, содержащих энергию.
Трансмембранный перенос веществ обеспечивает поступление на входы клетки необходимых веществ и выведение через ее выходы продуктов обмена и веществ, используемых другими клетками организма.
В процессе синтеза тканевых и клеточных структур, а также необходимых для жизнедеятельности соединений (анаболизма) энергия расходуется и накапливается. С пищей питательные вещества поступают, как правило, в виде продуктов, образующихся в результате гидролиза белков, жиров и углеводов. К ним относятся моносахара, аминокислоты, жирные кислоты и моноглицериды. Процесс синтеза обеспечивает восстановление структур клетки, подвергающихся распаду.
Размножение клеток в организме обеспечивает его рост и развитие, восстановление клеточных структур, способствует сохранению целостной структуры и нормальному функционированию организма.
Жизнедеятельность самой клетки обеспечивается взаимодействием всех ее органелл и клеточной мембраны. Как было сказано ранее в разделе 1.3.2, клеточные органеллы находятся в гиалоплазме, состоящей из воды и находящихся в ней различных ионов и органических веществ (глюкозы, аминокислот, белков, фосфолипидов и других). Гиалоплазма составляет внутреннюю среду клетки, обеспечивающую взаимодействие всех клеточных структур посредством транспорта веществ, потребляемых и синтезируемых клеткой. Гиалоплазма также хранит гликоген, липиды, пигменты. Большинство внутренних органелл имеют свои мембраны (ядро, эндоплазматический ретикулум, аппарат Гольджи, митохондрии, лизосомы). Они построены по тому же принципу, что и клеточные мембраны. Некоторые внутриклеточные органеллы не имеют собственной мембраны (рибосомы, микротрубочки, микрофиламенты и промежуточные филаменты). Каждая органелла выполняет свои специфические функции (таблица 1.4.1).
Таблица 1.4.1. Структура и функции основных клеточных элементов
Специфические функции характеризуются выполнением каждой клеткой определенной задачи, которая, в свою очередь, определяется генетически запрограммированным алгоритмом. Например, работа нервных клеток заключается в восприятии сигнала, его передаче, переработке и хранении информации. Возбуждение мембраны нейрона заканчивается выбросом медиатора в синаптическую щель. Таким образом, путем трансформации электрического импульса в химический сигнал происходит передача информации по всем звеньям нервной системы. Каждая секреторная клетка осуществляет синтез и выделение специфических веществ, важных для функционирования организма. В результате секреции выделяются слюна, желудочный и кишечный сок, желчь, молоко, гормоны и другие биологически активные соединения. Секреторные клетки участвуют в работе и регулировании функций многих органов: желудка, поджелудочной железы, щитовидной железы и других. Мышечные клетки в организме выполняют сократительную функцию: сокращения клеток поперечнополосатой мускулатуры обеспечивают работу опорно-двигательного аппарата, гладкой мускулатуры – работу внутренних органов.
В реальности организм человека существует, постоянно подвергаясь воздействию самых разнообразных и изменчивых внешних факторов. К ним могут быть отнесены температура окружающей среды, давление и влажность воздуха, концентрация в атмосфере вредных для организма веществ и так далее. Они могут меняться во времени как закономерным, так и случайным образом. На клеточном уровне схема внешних воздействий приведена на рисунке 1.4.3.
Рисунок 1.4.3. Системная модель клетки. Воздействие внешних факторов
Работа подсистемы жизнеобеспечения характеризуется поддержанием на генетически определенном уровне набора параметров: температура, концентрация белков, содержания воды, уровень кислотно-щелочного равновесия внутри клетки, ее мембранный потенциал и множество других. В процессе эволюции клетка научилась сохранять благоприятную внутреннюю среду, несмотря на изменение внешних условий. Главный механизм клетки как самоорганизующейся системы, способствующий поддержанию определенных величин в физиологически допустимых границах и заложенный в основу подсистемы жизнеобеспечения, называется гомеостатом. Само свойство клетки поддерживать постоянство внутренней среды на генетически заданном уровне называется гомеостазом. Клетка хранит информацию о значениях всех параметров, обеспечивающих ее жизнедеятельность и выполнение свойственных ей функций. Гомеостаз реализуется за счет использования механизма обратной связи (смотри рисунок 1.4.4). Более подробно об этом будет рассказано в разделе “Принципы и алгоритмы регуляции функций организма (информационный подход)”.
Рисунок 1.4.4. Системная модель клетки. Механизм обратной связи
Каждое мгновение жизни клетки характеризуется набором значений текущих параметров (показателей): температурой внутри клетки, концентрацией питательных веществ и других. Совокупность значений этих параметров в некоторый момент времени определяет состояние клетки как системы. Одни из данных параметров поддерживаются на неизменном уровне, другие могут меняться без потери устойчивости системы в целом.
Сам по себе известен и хорошо понятен принцип работы механизма обратной связи. Схема регулирования параметров клетки изображена на рисунке 1.4.4. Но, как внутри клетки одновременно и слаженно (синхронно) работают тысячи таких механизмов, и при этом происходит сравнение их текущих параметров с генетически заданными? Это остается загадкой природы.
Благодаря приспособительным (адаптационным) механизмам физические и химические параметры, определяющие жизнедеятельность клетки, меняются в сравнительно узких пределах, несмотря на значительные изменения внешних условий.
Зоны устойчивости характеризуются пределами изменений значений параметров входных сигналов подсистемы жизнеобеспечения, при которых процессы в клетке протекают нормально. В качестве входных сигналов можно рассматривать количество питательных веществ, содержание кислорода, углекислого газа, гормонов в крови и другие. Внутриклеточные параметры, например показатель кислотно-щелочного равновесия (рН), поддерживаются на заданном относительно постоянном уровне.
В цитоплазме клеток рН составляет 6,7-7,3 (разница, определяющая зону устойчивости, составляет 0,6). Более строгими являются требования к изменению этого показателя со стороны крови: рН крови может изменяться только в пределах 7,35-7,45 (зона устойчивости составляет 0,1, что в 6 раз меньше, чем для рН цитоплазмы клеток).
При отклонении значений этих параметров за пределы зон устойчивости изменяется скорость протекания биохимических реакций, вплоть до торможения. Активность большинства клеточных ферментов зависит от показателя рН, так как при его повышении внутри клеток нарушается структура белка и, в частности, ферментов. Считается, что увеличение рН внутри клеток поджелудочной железы служит одним из сигналов начала реакций запрограммированной их гибели (апоптоза).
Постоянство температуры внутри клетки также способствует оптимальному течению в ней химических реакций. Организм человека удерживает температуру тела на определенном уровне. Жизненные процессы в организме протекают в узких температурных границах: при температуре от 22 °C до 43 °C. Повышение температуры живых тканей выше 45-47 °С сопровождается необратимыми изменениями и прекращением жизни из-за свертывания белков и инактивации ферментов. При температуре ниже 22 °C наступает торможение работы клетки, обусловленное значительным замедлением обмена веществ и энергии.
Функционирование подсистемы, обеспечивающей выполнение специальных функций, также невозможно без механизма обратной связи, поддерживающего гомеостаз в клетке. Например, в системе гормональной регуляции постоянный уровень, в частности, кортикостероидов поддерживается благодаря такому механизму. Гипофиз отслеживает концентрацию данных гормонов в крови и при ее уменьшении выделяет в кровь адренкортикотропный гормон (АКТГ). АКТГ стимулирует образование кортикостероидов в корковом веществе надпочечников, концентрация гормонов увеличивается. При повышенном уровне гормонов, наоборот, идет сигнал на прекращение выработки АКТГ.
Существуют диапазоны колебаний внешних воздействий (температуры окружающей среды, уровня электромагнитных излучений и других), в пределах которых клетка остается устойчивой и работоспособной независимо от времени их воздействия. Приведем несколько примеров зон устойчивости при внешних воздействиях. Зимой и летом, при температуре окружающего воздуха в диапазоне от –70 до +50 °С температура тела человека остается практически постоянной, изменяясь всего на несколько долей градуса. В жаркий день даже небольшое повышение температуры тела дает сигнал к усилению активности потовых желез, кожа становится влажной, испарение воды с ее поверхности способствует охлаждению тела. И напротив, в холодную погоду поверхностные сосуды сужаются, потеря тепла уменьшается, а выработка – увеличивается, возникает защитная реакция – дрожь, “мурашки”.
Внутренние параметры клетки остаются в норме после прекращения действия возмущающего фактора, если он не превысил допустимые пределы. Таким образом, можно выделить допустимые интервалы внешних параметров (температуры, влажности, атмосферного давления, ионизирующего излучения и других), при которых система клеточного гомеостаза поддерживает относительное постоянство внутренней среды то есть возвращает параметры в нормальное состояние, при условии, что внешние воздействия не выводят их значения за пределы зон устойчивости.
Устойчивость в малом, но неустойчивость в большом. Будем говорить, что система устойчива в малом, но неустойчива в большом, если ограниченное изменение входного сигнала (набора входных сигналов) ведет к изменению в ограниченном диапазоне значений выходного сигнала (набора выходных сигналов).
Существование клетки в определенном диапазоне значений параметров хорошо прослеживается при воздействии радиации, или радиоактивного облучения. Учитывая, что каждый человек подвергается воздействию природной радиации, можно проследить, как ионизирующее излучение оказывает воздействие на клетку. Основу этого воздействия составляет передача энергии радиации клеткам организма.
На Земле всегда есть природный радиоактивный фон, который создают космическое излучение и радионуклиды, рассеянные в окружающей среде и всегда находящиеся в живых организмах. Радиация непрерывно воздействует на все живые организмы, в том числе на каждую клетку. Но ее уровень чрезвычайно мал, в среднем 0,2 сГрэй в год для человека, что в миллион раз меньше вредной для организма дозы облучения. Данный природный радиоактивный фон необходим для нормального существования клеточной системы.
Однако случайное облучение радиацией большой мощности способно привести к разрушению, повреждению и изменению определенных клеточных структур (белков, ДНК, РНК и их комплексов), гибели клеток. Большие дозы могут вызвать полное прекращение деления клеток.
К примеру, доказано, что кожа не выдерживает радиоактивного облучения или длительного и интенсивного облучения солнечным светом, так как разрушаются соединительнотканные структуры (коллаген и эластин), обеспечивающие плотность и упругость кожи, появляются признаки преждевременного старения.
Таким образом, если действует внешнее воздействие, при котором клетка теряет устойчивость, и нарушаются допустимые параметры ее существования, то возникает ряд патологических процессов, приводящих к гибели системы клетки в целом.
Управляемость клеточной системы. Каждая система, в том числе и система клетки, в любой момент времени находится в определенном состоянии, характеризуемом набором конкретных значений ряда параметров. Например, на мембранах нервных клеток существует определенная разность потенциалов, изменение которой приводит к возникновению и распространению нервного импульса по аксону. Переход из одного состояния системы в другое осуществляется за счет изменения значений параметров входных сигналов с учетом внешних воздействий. Так поступление внешнего стимула (света, шума) на рецепторную клетку приводит к изменению ее состояния (возбуждению), активации различных биохимических процессов. Будем говорить о том, что система управляема, если за счет изменения значений входных сигналов мы можем перевести ее состояние из начального в заранее определенное. Например, при избыточном поступлении кислорода в организм, происходит увеличение содержания кислорода в крови и соответствующая активация некоторых процессов в клетке (например, дыхания). Клетка начинает усиленно работать. Таким образом, управляемость системы – это способность перевода из текущего состояния в другие запланированные.
В данном разделе были отмечены и рассмотрены общие характеристики клетки как системы. Жизнедеятельность клетки связана с биофизическими, биохимическими, информационными и энергетическими процессами.
Читайте также:
- Смотренье что это кратко
- Почему экваториальные леса расположены только в западной части материка африка кратко
- К каким спектральным классам относятся самые горячие и самые холодные звезды кратко
- Организация деятельности школьного методического объединения учителей начальных классов
- Для чего нужны загадки детям в детском саду