
Что такое генетическое единство кратко
Обновлено: 04.07.2024
Генетическое единство - это общность ПТК, обусловленная единством их происхождения и процесса образования, возрастной однородностью, которые приводят к формированию относительно однородной внутренней морфологической структуры. Их генетическая однородность является следствием развития, на едином геологическом фундаменте. Генетическая однородность географических стран - следствие обособления их в пределах крупных тектонических структур: щитов, плит, платформ, геосинклинальных поясов. Географические области, ландшафты и некоторые местности формируются в пределах тектонических структур и блоков более низкого ранга. Некоторые местности, урочища, фации занимают генетически однородные формы и элементы форм рельефа. [1]
Генетическое единство ПТК и их видов, одинаковость их происхождения и процесса образования. [2]
Как проявляется генетическое единство живых существ . [3]
Некоторые дополнительные данные о генетическом единстве старогрозненских нефтей будут приведены далее в разделе, посвященном термолизу асфальтенов. Сопоставление изменения углеводородного состава этих нефтей по разрезу ( снизу вверх) с изменением углеводородного состава нефтей в процессе лабораторного моделирования процесса биодеградации дает, все основания отметить единую направленность всех превращений. [4]
В последние годы на основании геологических и геохимических исследований установлено генетическое единство подсо-левых и надсолевых нефтей восточной половины Прикаспийской впадины . [5]
Для установления характера миграционных процессов важнее выявлять не сходство в составе и степени зрелости ( это может быть следствием генетического единства или сходства состава ОВ нефтематеринских толщ), а направленности изменений того или иного параметра. Наиболее информативными показателями являются углеводородный, компонентный, фракционный и изотопный состав углеводородных систем. Но и для этих показателей нет строго установленных единых закономерностей, поскольку на направленность этих изменений влияет ряд факторов: форма переноса - струйная, диффузионная, растворенная; направленность миграционных процессов - латеральная или вертикальная и тесно связанные с этими факторами различные адсорбцион-но-хроматографические эффекты, которые определяются вещественным ( минералогическим, литологическим, гранулометрическим) составом среды, скоростью фильтрации и др. Состав пород определяет и многие физико-химические свойства нефти, которые также меняются в процессе миграции. [6]
Данные о распространенности химических элементов и их изотопном составе в изученных космических объектах Солнце, метеориты, лунные породы) и материале Зеили свидетельствуют о генетическом единстве всего вещества Солнечной сшетемы. [7]
Хант и др.) установили, что природные нефти, рассеянное органическое вещество в осадочных породах и липоиды планктона обогащены легким изотопом углерода ( 12С), что указывает на их генетическое единство . [8]
Важно отметить, что, за исключением углистого сланца, установлен однотипный характер распределения высших биомаркеров ( стеранов и гопанов) по разрезу осадочных толщ данного месторождения ( рис. 1, 2), свидетельствующий о генетическом единстве углеводородов и близкой степени их катагенетической преобразованности в отложениях различного возраста, что дает основание однозначно полагать о поступлении углеводородов в пермские отложения в результате миграционных процессов из нижележащих пород каменноугольного и девонского возраста и их геохимической преобразованности в результате постмиграционных процессов, протекавших в нефтевмещающих пластах. [9]
Количества термобитумоидов по данным Л. И. Хотынцевой и др. вполне соизмеримы с количеством УВ, сорбированных в ОВкл, устанавливаемых нами масс-спектрометрическими исследованиями, и температурные интервалы их максимального выхода также совпадают с нашими данными, что говорит об их генетическом единстве и решающей роли процессов десорбции при их выделении из ОВКЛ - Так, в частности, согласно нашим данным интенсивный выход УВ происходит до температуры 300 С ( максимум при 170 - 250 С), у Л. И. Хотынцевой и др. при 200 - 250 С. [10]
Согласно теории органического происхождения УВ ( ее осадоч-но-миграционному варианту) эволюция исходной материи ( остатков животных и растительных организмов) до конечного продукта в виде нефти и газа идет параллельно с процессом превращения осадка в горную породу. Этим подчеркивается теснейшая связь процессов преобразования неорганического и органического материала и генетическое единство природных УВ с определенными типами ОВ и вмещающих его пород. Однако, в то время как для осадочных пород эволюция достаточно ясно намечена в виде соответствующих стадий литогенеза, для находящегося в породе органического материала этапы его преобразования еще нуждаются в уточнении. [11]
Все они определенным образом субординированы в составе общественно-экономической формации. Эти формации структурированы, в том числе в плане изменений, обусловливая генетическое единство общественного развития в целом. [12]
Мы привели результаты наших экспериментально-лабораторных работ и основанные на них логические построения, подтверждающие генетическое единство УВ залежей и ОВ пород, которое мы рассматриваем как основной источник нефти и газа. Результаты этих исследований, по нашему мнению, позволяют уточнить существующие представления о стадийности нефте-газообразования и эмиграции УВ в связи с литогенезом, качеством ОВ и эволюцией геогидродинамических систем и предложить новый вариант объемно-генетического метода определения потенциальных ресурсов и прогнозных запасов нефти и газа. [13]
Таким образом, микробы и бактерии играют важную роль в преобразовании органического вещества, перерабатывая, насыщая его своими органическими соединениями. Широкое присутствие остатков микроорганизмов в форме гопана в ископаемых углях и неф-тях является хорошим доказательством генетического единства процесса образования и накопления в земной коре различных горючих полезных ископаемых. Происходит своеобразная утечка углерода при его круговороте между атмосферой и поверхностью Земли, которая за сотни миллионов лет привела к аккумуляции органического углерода в осадочных отложениях в количестве 10 т, что в 10 тыс. раз больше массы всех живых организмов. [14]
Проанализировав прежние схемы и способы интегрально го структурного анализа ( ИСА) тяжелых компонентов нефти [ 33, 44, 45, 52 - 56 и др. ], мы пришли к заключению о том, что ванкнейшим их недостатком является использование ряда допущений, не согласующихся с современными сведениями о составе и строении молекул нефтяных углеводородов и гете-росоединений, и что достоверность результатов ИСА можно повысить, по возможности исключив некорректные постулаты. Обобщение накопленных данных в работах [ 5, 63, 64 и др. ] показало, что особенности строения молекул нефтяных компонентов различных классов подчиняются принципам структурного и генетического единства [65], действие которых должно распространяться и на нефтяные ВМС. Иначе говоря, структурные характеристики полициклических блоков ( единиц) в макромолекулах нативных нефтяных ВМС должны быть очень близкими к характеристикам остальных компонентов нефти. [15]
Генетическое единство - это общность ПТК, обусловленная единством их происхождения и процесса образования, возрастной однородностью, которые приводят к формированию относительно однородной внутренней морфологической структуры. Их генетическая однородность является следствием развития, на едином геологическом фундаменте. Генетическая однородность географических стран - следствие обособления их в пределах крупных тектонических структур: щитов, плит, платформ, геосинклиналь-ных поясов. Географические области, ландшафты и некоторые местности формируются в пределах тектонических структур и блоков более низкого ранга. Некоторые местности, урочища, фации занимают генетически однородные формы и элементы форм рельефа.[ . ]
В пределах видов ПТК сходно протекает рост и развитие насаждений; для видов ландшафтных фаций свойственны свои специфические ряды сукцессий (Киреев, 1973; Фуряев, Киреев, 1979).[ . ]
ЛТК - это природные образования, относительно однородные по строению, структуре и экологическим режимам земель.[ . ]
Когда рассматриваются сложные ПТК: урочища, местности, ландшафты, то можно говорить только об их структурной однородности. Каждый сложный ПТК однороден структурно, так как состоит из фаций, урочищ, местностей только одного вида. Если внутри ландшафта появляются новые урочища, урочища другого вида, то это признак того, что мы перешли природный рубеж и вступили в другой ландшафт.[ . ]
Очевидно, и не требует доказательств, положение о том, что природная однородность увеличивается от крупных и сложных ПТК к более мелким и простым. Это происходит потому, что сужается ассортимент ПТК и их видов: их становится всё меньше, пока мы не дойдём в нашем анализе до простейшего ПТК - ландшафтной фации.[ . ]
Фации и их виды на одинаковых стадиях лесовосстановления однородны и идентичны по таксационным показателям древостоя. В их пределах значительно сужаются амплитуды варьирования целого ряда таксационных показателей древостоев, а взаимосвязь параметров полога леса и крон деревьев с таксационными показателями древостоев становится особенно прочной.[ . ]
Поэтому уже само выявление по аэроснимкам ПТК является и таксационным дешифрированием, так как делает лесные выделы более однородными по характеру древостоя. В пределах видов фаций снижается варьирование многих показателей, которые не всегда уверенно дешифрируют непосредственно по признакам изображения - класс бонитета древостоя, средней высоты, состава, запаса и др. Экологически однородные виды фаций предопределяют и такие важные показатели насаждений, как густоту, характер размещения деревьев по площади, состав и высоту ярусов, проективное покрытие полога, структуру насаждений и др. Таксационное сходство видов фаций связано с тем, что в них одинаково протекает процесс лесовосстановления.[ . ]
В литературе можно встретить указание на абсолютную однородность ландшафтной фации по всем составляющим её компонентам. С этим трудно согласиться. В пределах фации и её вида могут быть коренные, условно коренные и производные фитоценозы. Комплексные фации также однородны лишь структурно.[ . ]
Географические различия между видами основаны на относительной самостоятельности ареала каждого вида. Несомненно, что размер ареала, его форма, расположение в биосфере — важные видовые признаки. Для каждого вида свойственны свои границы пригодных для жизни условий, своя история возникновения (определяющая в значительной степени очертания и размер ареала), свои специфические взаимодействия с видами-конкурентами и т. д. Поэтому любая характеристика вида будет неполной без указания на специфические особенности его ареала.
Однако существуют причины, по которым географический критерий не может считаться достаточно универсальным для характеристики видов в природе. Во-первых, существует множество видов с практически совпадающими ареалами. Например, ареал кедрового стланика (Pinus pumila) совпадает с ареалами березы Миддендорфа (Betula middendorfii), даурской лиственницы (Larix daurica) и душистого тополя (Populus suaveolens). Эти виды тесно связаны биогеоценотически, входят в состав одних и тех же растительных сообществ, и их историческое развитие шло, видимо, параллельно, отражая общность исторических условий формирования определенных природных зон. Во-вторых, для видов-космополитов, ареал которых охватывает огромные пространства в биосфере Земли (в океане, например, дельфин косатка — Orcinus orca; на суше — некоторые виды рода дрозофил), характеристика ареала как видового признака также теряет смысл. В-третьих, определенность ареала отсутствует у некоторых быстрорасселяющихся видов, поскольку очертания ареала у них изменяются очень быстро и непредсказуемо (в последние столетия часто в связи с антропогенными изменениями ландшафтов или вселениями, как, например, для домовой мухи, городского воробья, многих рудеральных растений и т. д.).
Генетическое единство — главный критерий вида. Целостность вида. Возникает вопрос: а есть ли такой критерий? Есть, и он характеризует вид и как всеобщую форму существования жизни на нашей планете, и как качественный этап эволюционного процесса. Основной критерий вида заключается в его генетическом единстве. Единство вида как динамической системы в эволюции основано на возможности нивелировок различий, возникающих в популяциях путем скрещивания. Как бы ни были изолированы отдельные популяции и подвиды, поток генетической информации между ними всегда существует. Даже единичные миграции отдельных особей из одной популяции в другую, из зоны обитания одного подвида в зону обитания другого, повторяясь на протяжении сотен и тысяч поколений, создают поток генов. Это обеспечивает интеграцию относительно изолированных генофондов отдельных популяций (рис. 12.1).
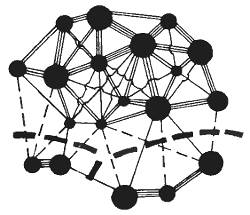
Рис. 12.1. Схема интеграции генофондов отдельных популяций в единый генофонд вида. Кружки — популяции, линии — потоки генов между ними, пунктир — возникающая изоляция частей видового генофонда (ориг. рис.)
Конечно, при отсутствии обмена гамет (у агамных, облигатно-партеногенетических и самооплодотворяющихся форм) не возникает такого генетического единства особей, что создает объективные трудности при разграничении видов.
Единство вида не нарушается даже и в том случае, если иногда по каким-то причинам границы между соседними видами оказываются нечеткими. Сейчас в разных группах таких случаев вскрывается все больше. Это связано с тем, что особи близких видов в определенных условиях иногда могут успешно скрещиваться друг с другом и давать плодовитое потомство в некоторых комбинациях. В таких случаях оказывается, что, за исключением некой гибридной зоны, между такими видами нарушения строения обоих видов как самостоятельных генетических систем не происходит: виды оказываются не генетически закрытыми системами, как представлялось еще в середине XX в., а генетически устойчивыми системами.
Гено́м — совокупность наследственного материала, заключенного в клетке организма [1] . Геном содержит биологическую информацию, необходимую для построения и поддержания организма. Большинство геномов, в том числе геном человека и геномы всех остальных клеточных форм жизни, построены из ДНК, однако некоторые вирусы имеют геномы из РНК [2] .
У человека (Homo sapiens) геном состоит из 23 пар хромосом, находящихся в ядре, а также митохондриальной ДНК. Двадцать две аутосомные хромосомы, две половые хромосомы Х и Y, а также митохондриальная ДНК человека содержат вместе примерно 3,1 млрд пар оснований [1] .
Содержание
Происхождение названия
Структура и размер генома
Известно, что ДНК, которая является носителем генетической информации у большинства организмов и, следовательно, составляет основу генома, включает в себя не только гены в современном смысле этого слова.
Генетическая информация в клетках содержится не только в хромосомах ядра, но и во внехромосомных молекулах ДНК. У бактерий к таким ДНК относятся плазмиды и некоторые умеренные вирусы, в клетках эукариот — это ДНК митохондрий, хлоропластов и других органоидов клеток (См. плазмон).
Объёмы генетической информации, заключённой в клетках зародышевой линии (предшественники половых клеток и сами гаметы) и соматических клетках, в ряде случаев существенно различаются. В онтогенезе соматические клетки могут утрачивать часть генетической информации клеток зародышевой линии, амплифицировать группы последовательностей и (или) значительно перестраивать исходные гены.
Следовательно, под геномом организма понимают суммарную ДНК гаплоидного набора хромосом и каждого из внехромосомных генетических элементов, содержащуюся в отдельной клетке зародышевой линии многоклеточного организма.
В определении генома отдельного биологического вида необходимо учитывать, во-первых, генетические различия, связанные с полом организма, поскольку мужские и женские половые хромосомы различаются.
Во-вторых, из-за громадного числа аллельных вариантов генов и сопутствующих последовательностей, которые присутствуют в генофонде больших популяций, можно говорить лишь о некоем усреднённом геноме, который сам по себе может обладать существенными отличиями от геномов отдельных особей.
Размер генома
Размер генома — общее число базовых пар ДНК в одной копии гаплоидного генома.
Размеры геномов организмов разных видов значительно отличаются друг от друга, и при этом часто не наблюдается корреляции между уровнем эволюционной сложности биологического вида и размером его генома.
Организация геномов
Эукариоты
У эукариот геномы находятся в ядре (Кариомы)и содержат от нескольких до многих нитевидных хромосом.
Прокариоты
У прокариот ДНК присутствует в виде кольцевых молекул.
Прокариотические геномы, как правило, гораздо меньше, чем у эукариот. Они содержат относительно небольшие некодирующие части (5-20 %).
Органеллы
Геномы митохондрий и пластид организованы как прокариотические геномы.
Вирусы
Вирусные геномы очень малы.
См. также
Примечания
Литература
- Сингер М., Берг П. Гены и геномы. — Москва, 1998.
- Стент Г., Кэлиндар Р. Молекулярная генетика. — Москва, 1981.
Ссылки
Информация должна быть проверяема, иначе она может быть поставлена под сомнение и удалена.
Вы можете отредактировать эту статью, добавив ссылки на авторитетные источники.
Эта отметка установлена 13 мая 2011.
Читайте также: