
Сообщение о древних папоротниках и голосеменных
Обновлено: 28.06.2024
Голосеменные— самые древние представители растений, которые произошли от папоротников.
Органы голосеменных: стебель, корень, листья.
Размножаются с помощью семян. Семязачатки располагаются на внутренней стороне листа или в шишке. Шишки, в свою очередь, делятся на мужские и женские. В мужской образуются пыльцевые зерна, которые с помощью ветра переносятся на женскую. Когда пыльцевое зерно попадает на женскую шишку происходит процесс оплодотворения.
У голосеменных преимущественно стержневая корневая система. Листья имеют различную форму:
- Чешуевидную;
- Игольчатую;
- Двулопастную;
- Разветвленную;
- Перистую.
Жизненные формы представлены кустарниками и деревьями, большинство из них вечнозеленые.
Голосеменные делятся на 4 класса:
- Класс хвойные (кедр, кипарис, можжевельник, пихта, лиственница, сосна, ель,тсуга, секвойя)
- Класс гинкговые (дерево гинкго билоба)
- Класс гнетовые (вельвичия)
- Класс саговниковые (саговые пальмы)
Самый многочисленный класс— хвойные. Первые хвойные появились еще во времена динозавров (370 млн. лет назад). Многие из них продолжают процветать и сегодня. Хвойные можно встретить на всех континентах Земли. Представлены деревьями (ель,сосна,секвойя,кипарис) и кустарниками (туя,можжевельник). Листья у хвойных игольчатые, которые называются хвоинками. А пучок хвоинок— это укороченный побег. В древесине вместо сосудов имеются трахеиды. Все хвойные имеют смолу, которая помогает заживлению древесных повреждений. Подразделяются на 7 семейств, самый распространенный— сосновые. Старейшим представителем, на сегодняшний день, является секвойя (3500 лет), его возраст можно определить по годовым кольцам. Секвойи могут достигать 100 метров высотой и 10-12 метров в диаметре.
Значение голосеменных огромно. Они образуют большие площади лесов, которые являются местом обитания для многих животных. Обогащают воздух кислородом, регулируют скорость таяния снегов, а также защищают почву от эрозии. Древесину широко используют в строительстве и фармакологии. Например из пихты изготавливают бумагу и используют в лекарственных целях. Из хвойных получают активированный уголь, эфирные масла и камфору. Также, они выделяют в воздух фитонциды, которые предотвращают развитие болезнетворных организмов. Поэтому большинство санаториев и больниц окружено этими деревьями.

Популярные сегодня темы
Наполеон Бонапарт это полководец и государственный деятель, заложивший крепкие основы нынешнего французского государства.
Антарктида является континентом, полностью покрытым льдом из-за весьма сурового климата. Но именно благодаря такой особенной экосистеме, в Антарктиде обитают разнообразные удивительные сущест
Древний Египет – одна из наиболее крупных и прогрессивных цивилизаций древнего мира, архитектурные сооружения которой дошли до наших дней. По всему миру известны египетские пирамиды – усыпаль
Впервые Смоленск упоминается в летописи (Повесть временных лет) в 862 году как главный город племенного союза кривичей. С той поры до настоящего времени он был центром разнообразных сражений
Человечество издавна пытало интерес к ночному небосводу и его космическим телам (звездам и галактикам). Пытливые умы не имели возможности подняться ввысь и рассмотреть эти странные рисунки на
Алоэ – травянистое многолетнее растение семейства Асфоделовые. Относится к роду суккулентных. Имеет короткий стебель с мясистыми сочными листьями. Края листьев могут быть зубчатыми и гладкими
Палеозой - эра древней жизни, продолжительность которой от 570 до 230 млн. лет. В эту эру в растительном и животном мире происходят значительные ароморфозы, связанные как с жизнью в воде, так и с освоением суши. Подразделяется на шесть периодов: кембрий, ордовик, силурий, девон, карбон, пермь.
Растения кембрия и ордовика населяют моря, представлены всеми отделами водорослей. В силурийском периоде (440 млн. лет назад) в зоне приливов и отливов от зеленых растений появляются первые наземные высшие растения - псилофиты (голые растения). Появление покровных, механических, проводящих тканей были теми ароморфозами, которые помогли выйти растениям в воздушную среду. У псилофитов еще отсутствуют корни, воду и минеральные соли они поглощали с помощью ризоидов. Чешуйки на стебле псилофитов увеличивали поверхность фотосинтеза.
В девоне появляются папоротникообразные - травянистые и древовидные хвощи, плауны, папоротники. Появление корней и листьев обеспечивало достаточное воздушное и минеральное питание разнообразным папоротникообразным. Размножаются папоротникообразные одноклеточными спорами, во влажных местах из них развиваются заростки, формирующие половые клетки. Для оплодотворения нужна вода, из зиготы развивается взрослое растение.
В карбоне устанавливается теплый и влажный тропический климат. Папоротникообразные достигают гигантских размеров - до 40 м в высоту. Каменноугольные леса впоследствии привели к образованию огромных залежей каменного угля. Вместе с тем в карбоне происходят два важнейших ароморфоза, в результате которых появились высшие семенные растения: во-первых, появляется опыление с помощью ветра, когда пыльца с мужскими половыми клетками по воздуху попадает на органы растений, содержащие женские половые клетки, вода для оплодотворения больше не нужна; во-вторых, после оплодотворения образуются семена. Такими растениями были семенные папоротники.
Семенные папоротники дали начало развитию голосеменных растений. В пермском периоде климат стал засушливым и более холодным. Тропические леса остаются у экватора, на остальной территории распространяются голосеменные.
В каменноугольной флоре широко были распространены споровые влаголюбивые растения (плуан, хвощи и папоротники). Затем началось появление голосеменных растений, в том числе и хвойных.Эти растения обладали значительными преимуществами по сравнению со своими предшественниками и поэтому не только шире их расселились по поверхности суши , но и в значительной мере вытеснили их из зон , которые те вначале безраздельно занимали. В связи с этим в мезозойскую эру господствующее положение на суши приобрели голосеменные растения.
Затем на смену им пришло господство покрытосеменных растений, еще более высокоорганизованных.
Отдел Папоротниковидные (Polypodiophyta)
В отделе папоротниковидные (Polypodiophyta) только один подкласс сальвинииды (Salviniidae) включает в себя разноспоровые растения. Десять видов современных сальвиний (Salvinia) являются остатками некогда крупного рода, возникшего по-видимому, в мезозое. Сальвинии – мелкие водные папоротники, замечательной чертой которой является разноспоровость. Мужские и женские гаметофиты развиваются из микро- и мегаспор, которые образуются в микро- и мегаспорангиях, заключенных в микро- или мегасорусах. Сорусы
образуются на спороносных сегментах погруженного листа. Мегасорусы, обычно на ножках, закладываются первыми у основания сегмента. Микросорусы, как правило, сидячие и размещаются на остальной части сегмента. Все сорусы по форме и величине сходны между собой. Мегасорус состоит из нескольких (до 25) мегаспорангиев, расположенных или одиночно на короткой толстой ножке, или группами на ветвистых ножках. В мегаспорангиях развивается только одна крупная мегаспора. Она остается все время окруженной стенкой спорангия и опадает вместе с ней. Окружающая мегаспору пенистая масса играет роль
плавательного приспособления как для мегаспоры, так и для развивающегося женского гаметофита.
В микросорусах сальвинии ушковидной образуется до 500 микроспорангиев. В каждом микроспорангии образуется 32 микроспоры. Микроспоры имеют ясно выраженный трехлучевой рубец. Созревшие микроспоры по 4 погружены в отвердевшую плазматическую массу, называемую массулой. Замечателен тот факт, что ископаемая сальвиния Рейза (Salvinia reusii) обладала амфиспорангиатными сорусами с многочисленными микроспорангиями и меньшим числом мегаспорангиев. У современных сальвиний были отмечены случаи развития наряду с нормальными не свойственных им амфиспорангиатных сорусов.
Сорусы созревают и погружаются на дно водоемов. У сальвинии плавающей (однолетнего растения) это происходит осенью. После сгнивания спорангия, поднимаются на поверхность. Микроспоры дают начало сильно редуцированным мужским гаметофитом, а мегаспоры прорастают в более крупные женские.
Прорастание спор происходит внутри спорангия. Прорастающая микроспора прорывает стенку спрангия и образует вначале трехклеточный мужской гаметофит. При прорастании мегаспоры ее оболочка разрывается на три лопасти, и зеленый женский гаметофит выступает наружу в виде веера.
Отдел Голосеменные (Pinophyta)
Растения, относящиеся к отделу голосеменных (Pinophyta) отличаются от всех остальных представителей растительного мира тем, что производят семена. А все семенные растения, как голосеменные, так и цветковые, являются разноспоровыми. Это значит, что споры у них двух разных типов – микроспоры (мужские споры) и мегаспоры (женские споры). Первые дают начало мужскому гаметофиту, а вторые – женскому. В этом отношении семенные
растения не отличаются от таких разноспоровых бесеменных растений, как селагинелла, полушник, марсилея или сальвиния. Тем не менее между ними имеются весьма существенные различия.
В то время как у огромного большинства бессеменных высших растений мегаспоры освобождаются из мегаспорангия, у семенных растений их единственная зрелая мегаспора остается постоянно заключенной внутри мегаспорангия и здесь же, внутри мегаспоры, происходит развитие женского гаметофита и процесс оплодотворения. Определенный шаг в этом направлении уже можно наблюдать у разноспоровых бессеменных растений. Так, у некоторых видов селагинеллы мегаспора остается внутри мегаспорангия и после оплодотворения, вплоть до значительного развития зародыша и даже молодого
растеньица. Еще больше приближение к семенным растениям наблюдается у некоторых вымерших плауновидных, например у каменноугольных родов миадесмия (Miadesmia) и лепидокарпон (Lepidocarpon), у которых мегаспорангий был окружен покровом из свернувшегося спорофиллоида (или его боковых выростов).
Это уже максимальное приближение к семени, но образование это является скорее аналогом, чем гомологом семени. Настоящее семя появляется только у голосеменных, при том значительно раньше, чем у таких растений, как миадесмия и лепидокарпон. В отличие от семенных плауновидных мегаспорангий у семенных растений окружен особым защитным покровом, называемым интегументом. Он умеет совершенно иное происхождение, чем покрывало мегаспорангиев миадесмии или лепитокарбона.
Мегаспорангий с окружающим его энтегументом называется семязачатком или семяпочкой. Это действительно зачаток семени, с которого после оплодотворения развивается семя.
Пыльцевые зерна у всех саговниковых мелкие, широкоэллиптические, лодочковидные и имеют на дистальной стороне одну борозду.
Например, у рода эфедра (Ephedra), относящийся к классу Гнетовые (Gnetopsida) при прорастании микроспоры отделяется сначала первая проталлиальная клетка. При втором делении образуется ядро второй проталлиальной клетки, но самой клетки не образуется, так ядро это не отделяется перегородкой от ядра антеридиальной клетки. Далее ядро антеридиальной клетки делится, образуя ядро генеративной клетки. Оно образует два ядра, одно из которых дает начало двум спермиям. А цель пыльцевых зерен попасть на женский гаметофит. Когда это произойдет, образуется короткая пыльцевая трубка, которая через шейку архегония достигает яйцеклетки. Спермий, достигнув яйцеклетки, сливается с ней,
У представителей подкласса Хвойные (Pinidae) каждая функционирующая материнская клетка микроспор дает начало четырем гаплоидным микроспорам, которые в течение некоторого времени оказываются заключенными внутри оболочки микроспор. Число микроспор обычно очень велико, иногда исключительно большое. Они чрезвычайно легкие, что помогает их распространению ветром. Чрезвычайно своебразной является форма пыльцевых зерен. У большинства представителей семейства сосновых и подокарповых она
бобовидная, с двумя выступами – воздушными мешками. Для большинства видов тсуги (Tsuga) типична шарообразная пыльца с одним опоясывающим воздушным мешком. Шарообразные безмешковые пыльцевые зерна имеются у лиственницы.
Самым старым деревом на Земле считается тоже голосеменное растение - сосна остистая (Pinus longaeva или P.aristata), растущая в горах Восточной Невады. Радиоуглеродный метод анализа показал, что возраст этого дерева составляет около 4900 лет.
Найдено недостающее звено между цветковыми и голосемянными растениями
Цветущее растение Амборелла (Amborella) оказалось живым ископаемым, выжившим на Земле в течение 130 миллионов лет. Профессор Вильям Фридман (William Friedman) из университета Колорадо в Болдере (University of Colorado at Boulder) полагает, что Амборелла — недостающее звено между цветущими растениями и их предками.
Амборелла — это маленький кустарник с крошечными зеленовато-жёлтыми цветами и красными фруктами, который растёт только в дождевых лесах Новой Каледонии.
Используя лазер, флуоресценцию и электронную микроскопию, Фридман и его коллеги установили, что цветущее растение Амборелла отличается от прочих покрытосемянных. Если коротко: Амборелла имеет одну дополнительную бесплодную клетку, которая сопровождает яйцеклетку в женской части её репродуктивного аппарата, зародышевом мешке.
Учёные считают, что ангиоспермы (покрытосемянные, они же — цветущие растения) отклонились на эволюционном дереве от растений-гимноспермов (голосемянных) примерно 130-150 миллионов лет назад, в эпоху динозавров, став теперь доминирующими растениями на Земле.
Дополнительная клетка в Амборелле, полагают авторы работы, является следом бурных эволюционных изменений в ранних покрытосемянных растениях, своего рода реликтом более примитивного механизма размножения. И также этот необычный эмбриональный мешок Амбореллы, согласно Фридману, — первая новая разновидность полового аппарата у цветковых растений, открытая за последние 50 лет.
Вероятно, что детальное изучение этого растения поможет пролить свет на то, что ещё Дарвин назвал "отвратительной тайной" — происхождение цветковых растений. Этот, мало ещё исследованный шаг эволюции растений представляет собой один из самых крупных вызовов для современных биологов.
Необычная структура Амбореллы, в конечном счёте, может связать её с голосемянными растениями, став, полагает Фридман, таким образом, недостающим звеном эволюции. Да ещё и живым, словно кистепёрая рыба.
Изучая палеозойские и раннемезозойские растения, находимые в земных слоях в виде отпечатков и окаменелостей, палеоботаники нередко встречают ископаемые листья, по строению неотличимые от перистых листьев настоящих папоротников, но несущие семена на своих верхушках или по бокам жилок.
Такое удивительное сочетание признаков семенных растений и папоротников, внервые установленное в 1904 г. выдающимися английскими палеоботаниками Ф. Оливером и Д. Скоттом, и дало повод назвать эту группу растений семенными папоротниками (Pteridospermae).
Это была обширная группа растений, из которой к настоящему времени описано несколько сот видов, объединяемых в четыре основных порядка — лигиноптерисовые (Lyginopteridales), медуллозовые (Medullosales), кейтониевые (Caytoniales) и глоссоптерисовые (Glossopteridales) и около 10 семейств.
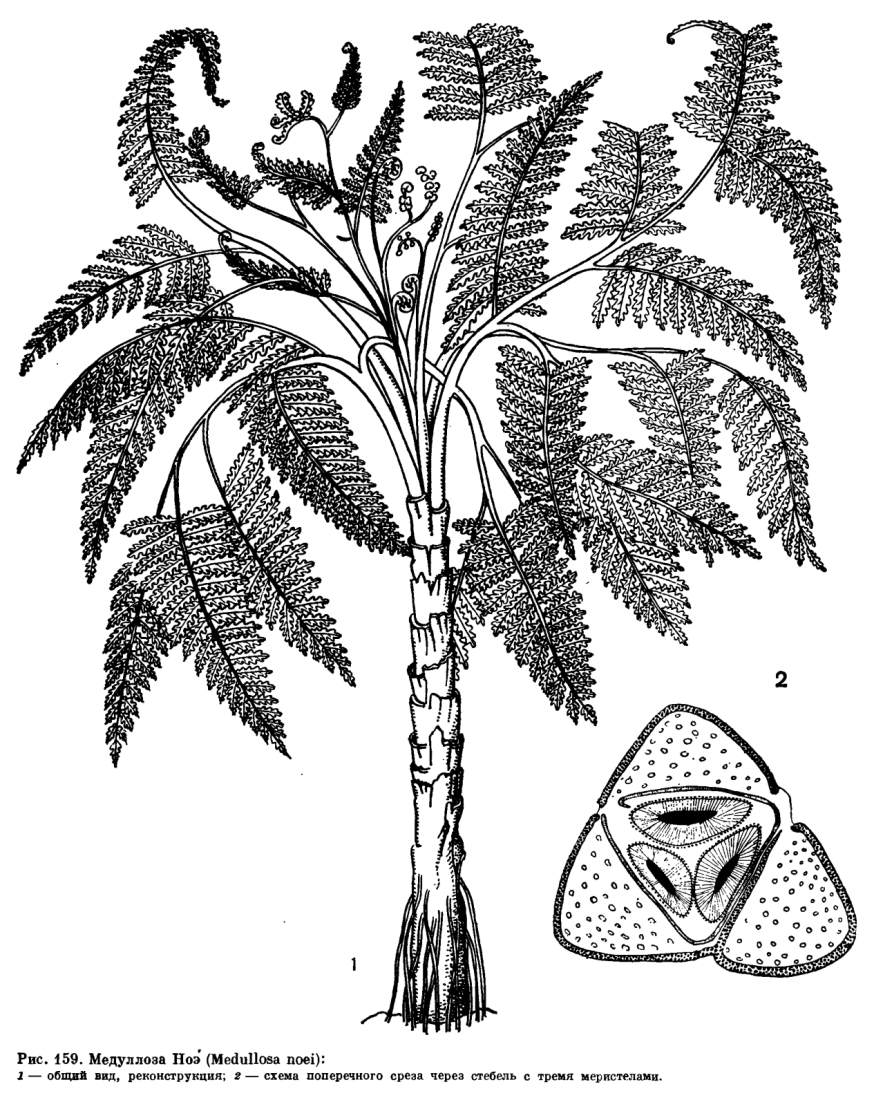
Внешний вид этих вымерших растений наиболее удачно передает реконструкция медуллозы Ноэ (Medullosa noei), выполненная американскими палеоботаниками В. Стюартом и Т. Делевориасом по отпечаткам листьев и срезам окаменелых стеблевых частей (рис. 159).
Семенные папоротники — полностью вымершие растения. Они существовали с позднего девона до раннего мела. Наиболее часто их остатки встречаются в отложениях карбона и перми. Одни из них были распространены по всему земному шару, другие, как, например, ангароптеридиум (Angaropteridium), были свойственны только ангарским флорам, а глоссоптерисовые и користоспермовые, напротив, приурочены главным образом к районам, соответствующим древнему материку Гондване, который существовал с середины карбона до конца триаса.
Наиболее распространено мнение, что семенные папоротники карбона были обитателями более возвышенных участков, обрамлявших приморские заболоченные леса. Однако все больше данных за то, что, например, заключительные этапы существования формации карбопового низинного заболоченного леса, как правило, сопровождались пышным развитием семенных папоротников или кордаитовых.
Семенные папоротники были похожи на настоящие папоротники не только по строению перистых листьев, но и по внешнему облику растений: древовидных, лиановидных и травянистых.
Стебли древовидных семенных папоротников были или прямыми, или дихо- и моноподиально ветвящимися. У одних листья сидели в очередном порядке на радиально-симметричном стебле, у других были сближены на верхушке и располагались в виде кроны, как у многих современных древовидных папоротников. Поверхность стебля многих растений была покрыта волосками, а иногда особыми железками, как, например, железками булавовидной формы у калиматотеки (Calymmatotheca), которые выполняли, по-видимому, секреторную функцию. У одних растений стволы были более или менее гладкие, с рубцами от опавших листьев; у других покрыты основаниями опавших листьев, как у медуллозы. У лигиноптерисовых (Lyginopteridaceae) стебли были диаметром 2—4 см, а у медуллозовых (Medullosaceae) их диаметр достигал 20 см. У некоторых семенных папоротников на стебле между листьями и в узлах располагались воздушные корни. Стебли пельтаспермовых (Peltaspermaceae) и кейтониевых (Caytoniaceae) до сих пор не найдены, и обе группы известны только по находкам стерильных и фертильных листьев и семян.
Листорасположение у семенных папоротников было спиральным. Как у лигиноптерисовых, так и у медуллозовых листья были довольно крупными и в основании вильчато делились на две симметричные половины, каждая из которых была перисто рассечена. В почкосложении листья были улиткообразно закручены, как у современных и многих ископаемых папоротников.
У растений из порядка глоссоптерисовых листья сидели спирально или мутовчато (глоссоптерис) в утолщенных узлах тонких стеблей. Они были довольно крупными, но очень различными по форме — цельные, цельнокрайние, линейные, ланцетные, лопатчатые, яйцевидные, сидячие и на коротких черешках. Если у более древнего рода гангамоптерис (Gangamopteris) не было ясно выраженной средней жилки, то у глоссоптериса (Glossopteris) она была развита хорошо.
Размножение семенных папоротников происходило с помощью семян, в которых до сих пор не найдены зародыши. Отсутствие зародыша, констатированное при изучении многих сотен ископаемых семян этих растений, до сих пор представляется загадкой. Существует точка зрения, согласно которой семенные папоротники не имели настоящего семени, хотя у них был семязачаток. В связи с этим их, а также современные саговниковые и гинкговые причисляли не к семенным, а к так называемым семязачатковым растениям. Однако едва ли это решает вопрос. Несомненно, что зародыш должен был развиться до прорастания этого семязачатка. По-видимому, развитие зародыша у семенных папоротников происходило в семязачатках, уже опавших с материнского растения на почву, как это происходит нередко у ныне живущего голосеменного гинкго двулопастного.
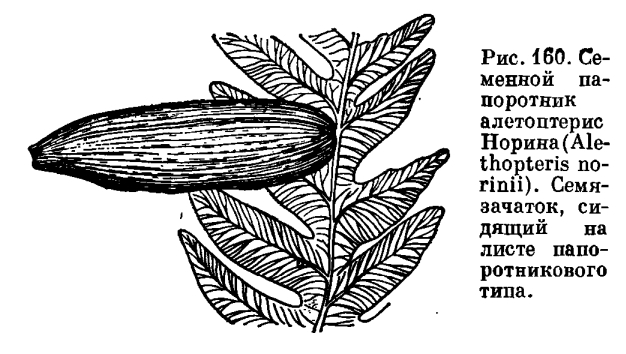
Семязачатки развивались на расчлененных папоротниковидных листьях, располагаясь или по краю, или на верхушке листа (рис. 160). У наиболее примитивных родов семязачатки сидели на концах боковых или верхушечных перышек (сегментов), у более подвинутых родов (например, у сфеноптериса тонкого — Sphenopteris tenuis) — на поверхности перышек. Иногда положение семязачатков было вторичноверхушечным за счет редукции и видоизменения перышек в результате их превращения в семяножку.
У большинства семенных папоротников семязачатки были окружены так называемой купулой, т. е. особым покровом, имеющим у различных растений очень разное строение и форму, иногда чашевидную, конусовидную или дисковидную. Считается, что купула возникла из сегментов (долек) листа, окружающих семязачаток. Листовое происхождение купулы подтверждается ее строением у наиболее древних представителей группы.
Семязачатки были одиночными, как у лигиноптерисовых, или многочисленными, как у глоссоптерисовых. У лигиноптерисовых они достигали в длину 5,5 мм, у пельтаспермовых— 7 мм, а у користоспермовых — 3,5—7 мм.
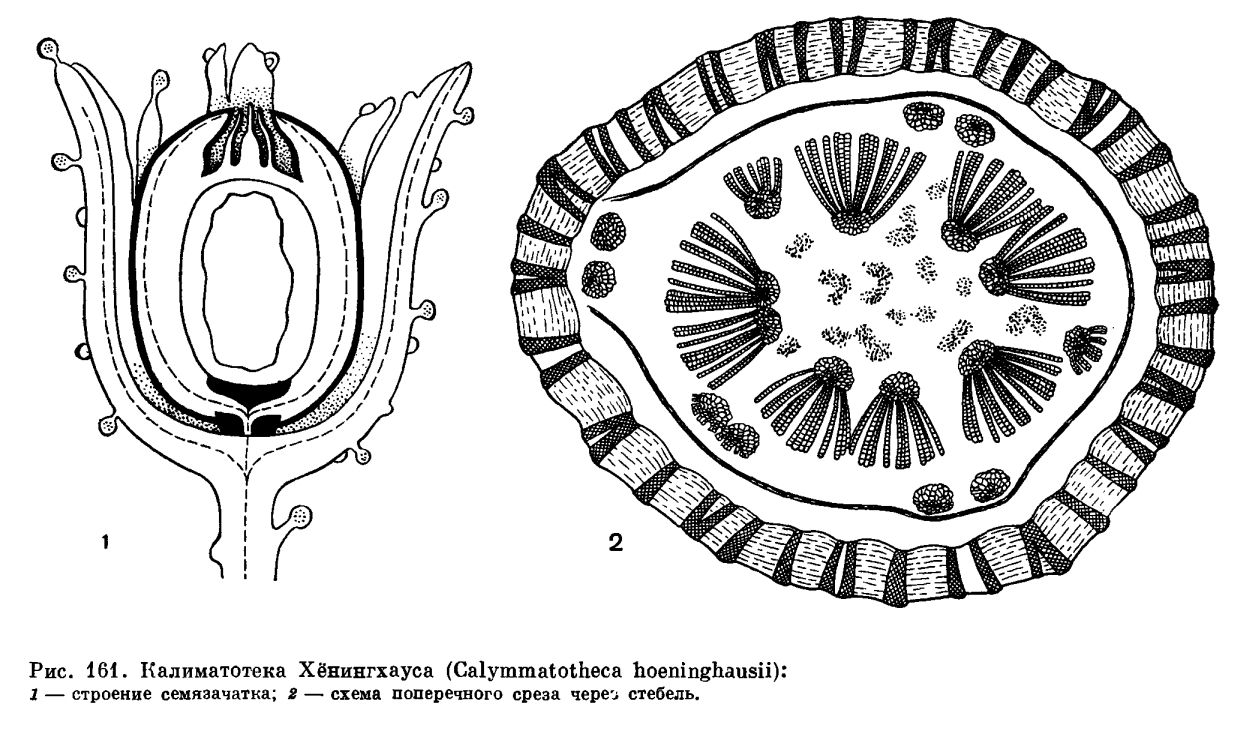
Рассмотрим строение семязачатка на примере калиматотеки Хёнингхауса (Calymmatotheca hoeninghausii, рис. 161). Снаружи семязачаток окружен купулой, несущей железки. Из ножки купулы проводящий пучок переходит в ножку семязачатка, имеющего овальную форму. По строению проводящий пучок ножки сходен с пучком листового черешка. Наружный покров семязачатка (интегумент) возникает, по мнению ученых, в результате длительной эволюции из кольцеобразно расположенных спорангиев, которые, потеряв фертильность и срастаясь между собой, обеспечили защиту центральному, сохранившему фертильность мегаспорангию. Таким образом, семязачаток — это синангии. Подтверждение этому — большое количество примитивно устроенных семязачатков, у которых сохранились признаки синангиальной структуры. Нуцеллус, соответствующий мегаспорангию, обычно тесно спаян с интегументом, но иногда, оставаясь свободным в верхушечной области семязачатка, он образует так называемую колонку пыльцевой камеры, в которой нередко находят пыльцу, причем иногда сохранившуюся в проросшем состоянии. На этом основании сделано предположение, что пыльцевая камера была заполнена сахаристой жидкостью, которая являлась благоприятной средой для прорастания пыльцы. Центральная часть семязачатка занята большой функционирующей мегаспорой, от которой у семенных папоротпиков чаще сохраняется только оболочка, но нередко также ткань гаметофита с архегониями, в которых даже сохраняются яйцеклетки.
Мужские спороношения у растений этой группы — обычного папоротникового типа. Они представлены или краевыми микроспорангиями, как у лигиноптерисовых, или микросинангиями, иногда сложно устроенными, как у медуллозовых и кейтониевых. Пожалуй, наиболее интересны сложно устроенные и очень разнообразные по форме мужские спороношения медуллозовых.
Микроспоры семенных папоротников были разными по величине и строению. Например, у пельтаспермовых они были очень мелкими, диаметром всего 30 мкм, однобороздными, а у лигиноптерисовых и медуллозовых — диаметром до 400—500 мкм, и имели одну дистальную борозду наряду с трехлучевым тетрадным рубцом. Проросшие микроспоры этого типа были найдены в пыльцевых камерах некоторых семян, в частности, у пахитесты (Pachytesta). Предполагают, что гигантские микроспоры переносились на семязачатки, видимо, еще до полного развития мужского гаметофита, который завершал свое развитие в питательной жидкости пыльцевой камеры семязачатка.
Семенные папоротники — группа наиболее примитивная среди голосеменных растений. Одни ученые приходят к выводу, что они занимают промежуточное положение между настоящими папоротниками и голосеменными, в то время как другие считают эти группы возникшими и эволюционировавшими параллельно. Как уже сказано, листья семенных папоротников часто очень похожи на листья настоящих папоротников или даже неотличимы от них. Это представляет яркий пример параллельного развития гомологичных органов. Но наличие семян, анатомическое строение стебля и корня и способность семенных папоротников к вторичному росту достаточно резко отличают их от настоящих папоротников.
Анатомическое строение стеблей и корней семенных папоротников очень специфично. Так, стебель молодого растения калиматотеки был протостелическим, но с возрастом он становился сифоностелическим с хорошо развитой сердцевиной и гнездами склеренхимных клеток (рис. 161). Вокруг сердцевины располагались многочисленные мезархные пучки, их метаксилема состояла из крупных (в сечении) лестничных, сетчатых и точечных трахеид. Последние несли многорядные окаймленные поры на всех стенках. Как у большинства семенных папоротников, у калиматотеки была довольно мощная вторичная ксилема, состоящая также из точечных трахеид, но с порами только на радиальных стенках. Довольно широкие древесинные лучи, состоящие из паренхимных клеток, разбивали вторичную ксилему на секторы. Интересной особенностью внутреннего строения стебля этого семенного папоротника является структура наружной коры, которая состояла из сложного сетчатого переплетения тяжей склеренхимных клеток. На тангентальном срезе они имели вид ячеистой стенки, а на поперечном — вид радиально вытянутых полос, имеющих форму трабекул. Петли сетки были заполнены паренхимной тканью. Листовые следы имели двухпучковое строение. В основании листа эти два пучка срастались в F-образный (на поперечном срезе) пучок.
У медуллозовых строение как стебля, так и листовых черешков было принципиально иным. Их стела рассечена на ряд элементарных стел (меристел), каждая из которых развивала свои вторичные проводящие ткани — ксилему и флоэму (рис. 159). Число меристел у видов медуллозы было различным. Хотя меристелы имеют вид самостоятельных проводящих систем, они анастомозируют друг с другом, соединяются в узлах, затем снова расходятся. Наружная кора у медуллозовых была обычно более просто устроена, чем у лигиноптерисовых, и состояла из массивной паренхимной ткани, в которой были рассеяны одиночные секреторные каналы, как у мараттиевых и саговниковых. Меристелы были погружены в основную ткань стебля, представленную паренхимой, в наружной части которой проходили многочисленные листовые следы. Листовые следы в своей нижней части близ стелы стебля имели концентрическое строение, а в наружной зоне коры и в листовом черешке были представлены многочисленными коллатеральными пучками. Этим они очень напоминают стебли однодольных растений.
Остатки семенных папоротников играют большую роль в сложении каменных углей СССР, Западной Европы и Северной Америки. Почти на каждом срезе через угольные почки, составляющие окаменевшие участки древнего торфяника, превращенного в уголь, можно найти стебли, корни, семена и пыльцу семенных папоротников.
Группа семенных папоротников взята на вооружение биостратиграфами, занимающимися вопросами расчленения палеозойских и мезозойских отложений. Например, по появлению отпечатков листьев растения каллиптерис (Callipteris) проводится нижняя граница пермского периода.
Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров . 1974 .
Голосеменные растения являются группой растительных организмов, не обладающих специфическими способами защиты семян.
Классификация голосеменных выглядит следующим образом:
- царство — Растения;
- подцарство — Высшие растения;
- отдел — Голосеменные растения.
К голосеменным растениям причисляют:
- гинкговые;
- гнетовые;
- хвойные;
- саговниковые.
Голосеменные — это около тысячи видов, восемьдесят родов и четырнадцать семейств.
У таких растений наблюдаются определенные адаптации, благодаря которым они вышли на новый эволюционный уровень. Это:
- редуцированный гаметофит, находящийся внутри спорофита;
- неподвижность мужских гамет, переносимых при помощи ветра, насекомых и пыльцевых зерен. Их оплодотворение происходит без воды;
- развивающийся внутри семени зародыш получает защиту и систематическую пищу в виде питательных веществ;
- наличие устьиц на эпидерме наземных органов и эпидерме листьев. Это обеспечивает хороший газообмен между растением и атмосферой.
Появление семенного размножения и уменьшение роли гаплоидной стадии развития — важнейшие ароморфозы в ходе эволюции растений.
Развитие семян голосеменных происходит на поверхности спорофилла. Оно защищается в результате смыкания спорофилла.
Когда возникли голосеменные растения
Поздний карбон — а это примерно 319 млн. лет назад — время появления голосеменных. После они распространились по всем широтам, в том числе они стали обосновываться в достаточно сухих местах.
Для голосеменных свойственно размножение при помощи семени, а в их жизненном цикле преобладает диплоидный спорофит. Эта группа растений имеет женские и мужские половые органы (антеридии и архегонии).
Также у них есть еще одна особенность: длительное функционирование камбия, который образует ксилему или древесину, флоэму и луб.
Камбий представляет собой образовательную ткань голосеменного растения, способную дать начало любому типу растительной ткани.
Также у них имеется главный корень, стержневая корневая система, благодаря которой развиваются древесные формы. Для голосеменных также характерная такая жизненная форма как кустарник.
Нарастание побегов у голосеменных осуществляется моноподиально.
Древние папоротниковидные (вымерли в начале каменноугольного периода) — предки первых голосеменных растений. Расцвет голосеменных приходится на мезозойскую эру, которая характеризуется горообразованием, поднятием материков и иссушением климата. Но начиная с середины мелового периода на смену голосеменным постепенно приходят покрытосеменные.
Современные голосеменные растения представляют собой отдел, включающий свыше 700 видов — это растения, завоевавшие всю планету. Распространение голосеменных пришлось и на умеренные широты Северного полушария. Здесь они сформировали лесное пространство — тайгу.
Современные голосеменные включают 4 класса (гинкговые, саговниковые, гнетовые и хвойные). При этом гинкговые — это всего один вид: Ginkgo biloba. Особо стоит выделить саговниковые, являющиеся остатками вымерших групп, которые процветали в мезозое.
А вот у гнетовых довольно неясное происхождение: палеоботанических данных о них сохранилось немного. Зато класс Хвойные представлен широко и полно. Его видовое разнообразие превосходит все остальные группы голосеменных. Хвойные — это свыше 500 растительных видов. Большая часть таких растений — вечнозеленые. Их возраст говорит о древнем происхождении.
Североамериканская сосна — почетный долгожитель.
Эта группа растений находится где-то между папоротниками и покрытосеменными. В отличие от папоротников, у этой группы растений есть семязачаток, а в отличие от покрытосеменных — у них нет плодолистиков.
Листья голосеменных разнообразны по морфологическим и анатомическим признакам. У большинства видов листья игловидные, у некоторых растений листья довольно большие.
Время появления голосеменных точно установить трудно, их предков обычно ищут среди господствовавших в палеозое растений. Возможными их предками многие считают древнейших представителей отдела папоротниковидные. Среди них были разноспоровые древовидные формы с вторичной древесиной — они вполне могли дать начало голосеменным.
Также голосеменные могли появиться из группы высокоорганизованных папоротников девона — из рода археоптерис. Такие растения имеют сердцевину, хорошо развитую вторичную ксилему, а трахеиды характеризуются точечной формой с окаймленными порами. Уровень организации вегетативных органов ничем не отличает археоптериса от некоторых пермокарбоновых хвойных и определенных современных форм.
Некоторые ученые обозначают самостоятельность происхождения хвойных и родственных им видов. Однако поспорить с этой точкой зрения легко, если приводить в качестве аргумента наличие общего плана строения семязачатка у всех голосеменных.
На единство происхождения голосеменных указывают особенности строения вегетативных органов.
Также считают, что голосеменные могли иметь предка, имеющего протостелическую организацию стелы и слабо развитую вторичную ксилему. Для трахеид таких растений, вероятно, была характерна лестничная форма.
Поэтому можно говорить о том, что предками голосеменных были не настоящие папоротники, а одна из боковых разноспоровых ветвей папоротниковидных живых организмов. На логичность и стройность этого факта указывает на то, что в ходе эволюции новые растительные группы происходят не от высшего таксона предков, а от менее специализированных представителей растительной группы.
Читайте также: