
Риниофиты сообщение для 5 класса
Обновлено: 05.07.2024
Эта статья — о растениях Девонского периода (старое название Псилофиты). О современных растениях см. Псилотовые.
† Rhyniopsida Kryshtofovich, 1925
Кладограмма сосудистых растений, положение риниофитов показано синим. Диаграмма построена согласно Кенрику [3] . Современные молекулярные данные изменяют положение риниофитов на среднее между антоцеротовидными (на кладограмме — hornworts) и мхами (на кладограмме — mosses) [4]
Данная группа растений парафилетического происхождения включает в себя высшие растения, имеющие проводящие ткани. С начала XXI века к риниофитам относят роды растений Huvenia, Rhynia и Stockmansella [3] , которые возникли в девоне. Ископаемых представителей риниофитов находят в окаменевших породах, начиная с раннего девона [3] , что делает эти отложения очень важными в палеоботанике. Считается, что представители этого отдела являются первенцами сухопутных растений.
Риниофиты необычны тем, что прекрасно сохранились в очень древних окаменелостях и являются самыми ранними известными сосудистыми растениями. Данные окаменелостей содержат убедительные детали, подтверждающие наличие ребристого стебля, покрытого чешуевидными придатками, а также крупных двойных спорангиев. Окаменелости содержат следы спорофитов и гаметофитов других растений, остатки животных, например, членистоногих, которые обитали в одной экосистеме с риниофитами.
Известно, что риниофиты не имели полноценных корней, вместо них — корневищеподобные образования, покрытые ризоидами.
Этот отдел объединяет первые достоверные ранние палеозойские наземные растения, описанные по сохранившимся остаткам. Они обладали рядом признаков, отражавших начальную ступень в эволюции высших растений . У них еще не было типичных для высших растений олиственных побегов и корней.
Содержимое работы - 1 файл
CSR.docx
Этот отдел объединяет первые достоверные ранние палеозойские наземные растения, описанные по сохранившимся остаткам. Они обладали рядом признаков, отражавших начальную ступень в эволюции высших растений . У них еще не было типичных для высших растений олиственных побегов и корней.
Тело риниофитов состояло из цилиндрических осей, дихотомически ветвившихся во взаимно перпендикулярных плоскостях. Существенным отличием от водорослей было наличие в их осях проводящей системы, построенной по типу протостелы , и типичной эпидермы с устьичным аппаратом. Подобные первичные васкуляризованные осевые структуры получили название теломов . Одни теломы были вегетативные, другие заканчивались спорангиями . Подземные теломы, получившие название ризомоиды, т.е. корневищеподобные , были снабжены простыми одноклеточными ризоидами . Спорангии , обладавшие многослойной стенкой, имели овальную или округлую форму, но у наиболее примитивных видов они мало отличались от верхушек вегетативных осей.
Риниофиты были равноспоровыми растениями, их споры имели трехлучевые щели, по которым, по-видимому, происходило их вскрывание при прорастании. Наиболее ранние риниофиты, описанные только по спорам, датируются нижнесилурийским периодом , лишь из верхнего силура описан род куксония (Cooksonia) по макроостаткам.
Первое описание одного из представителей риниофитов было произведено в 1859 г. американским палеоботаником Даусоном, который назвал его псилофитом (Psilophyton) , что означает голое растение, так как оно было лишено листьев и корней. По роду псилофит вся группа первых наземных растений получила название псилофиты. Однако в результате дальнейшей ревизии род Psilophyton перестал существовать, и поэтому первоначальное название оказалось неправомочным. В настоящее время по наиболее полно описанному роду риния (Rhynia) вся эта группа растений получила название риниофиты, или риниофитовые (Rhyniophyta). Поскольку риниофиты являются родоначальниками всех последующих групп папоротникообразных , включая папоротниковидные , членистые , плауновидные и праголосеменные , то им дано другое название - прапапоротникообразные (Propteridophyta) . Разделение некоторыми авторами этого отдела на 2 - риниофитовые и зостерофиллофитовые не вполне оправданно, так как нет четких объективных критериев для того и другого отдела в связи с несовершенной сохранностью остатков. Кроме того, существуют так называемые синтетические виды, совмещающие в себе признаки обеих групп.
Изучение строения Риниофиты и их эволюционных взаимоотношений имеет большое значение для эволюционной морфологии и филогении высших растений. По-видимому, первоначальным органом спорофита высших растений был дихотомически ветвящийся стебель с верхушечными спорангиями; корни и листья произошли позже спорангия и стебля. Есть все основания считать Риниофиты исходной предковой группой, от которой произошли мохообразные, плауновидные, хвощевидные и папоротники. Согласно другой точке зрения, мохообразные и плауновидные имеют лишь общее происхождение с Риниофиты
Отдел риниофитовые условно разделен на 2 класса - риниевые и зостерофилловые и, в целом, классификация отдела выглядит следующим образом:
Отдел Риниофиты (риниофитовые), или прапапоротникообразные (Rhyniophyta, или propteridophyta)
Класс Риниевые (Rhyniopsida)
Порядок Риниевые (Rhyniales)
Порядок Тримерофитовые (Trimerophytales)
Класс Зостерофилловые (Zosterophyllopsida)
Открытие и изучение риниофитов послужили основой для создания теломной теории, автором которой был немецкий ученый Циммерманн. Эта теория раскрывала особенности строения первых наземных растений и показывала возможные пути, по которым происходило становление основных вегетативных и репродуктивных органов высших растений различных систематических групп.
Согласно представлению Циммерманна, тело первых наземных растений состояло из радиально симметричных осей. Их конечные веточки получили название теломов (от греч. telos - конец). Теломы ветвились дихотомически во взаимно перпендикулярных плоскостях, образуя объемную систему теломов. По мере ветвления теломы из конечных становились промежуточными, поэтому получили название мезомов . Поскольку все мезомы когда-то были теломами, их иначе называют теломами в широком смысле слова.
Отличительной особенностью теломов было наличие проводящих пучков, построенных по типу протостелы . Этим они принципиально отличались от водорослей . Подземные, или стелющиеся, теломы получили название ризомоидов , т.е. корневищеподобных, так как никаких побегов, а следовательно, и корневищ у первых наземных растений еще не было. На ризомоидах располагались ризоиды . Вертикальные теломы либо оставались стерильными - вегетативными, либо заканчивались спорангиями .
Совокупности теломов и мезомов нескольких порядков ветвления называются синтеломами ; они могли быть вегетативными, спороносными и смешанными, т.е. состоящими из вегетативных и спороносных теломов.
В ходе эволюции теломы претерпели ряд существенных изменений и в итоге превратились в основные органы высших растений - стебли , листья , корни , спорофиллы . Поэтому нельзя проводить гомологию между теломами и органами современных высших растений.
Существует понятие временной границы теломов - нижней и верхней; у водорослей теломов еще не было, а когда теломы превратились в производные органы, они уже перестали существовать как таковые.
Превращение теломов в более сложные органы могло осуществляться в результате ряда процессов, протекавших независимо друг от друга. Важнейшие из них - перевершинивание, планация, срастание, редукция.
Превращение происходило в результате неравной дихотомии, что хорошо прослеживается у риниофитовых. В итоге формировались главная ось и система боковых ветвей; это явилось предпосылкой для создания крупных форм растений.
Под планацией понимается расположение теломов в одной плоскости. Этот процесс явился важнейшим этапом в ходе формирования листьев папоротников , членистых , праголосеменных растений . Срастание теломов как конечный результат филогенетического процесса мог распространяться на теломы, расположенные как в одной, так и в нескольких плоскостях. Если срастание сочеталось с планацией, то формировались крупные рассеченные листья с системой разветвленных жилок.
Срастание теломов, расположенных в разных плоскостях, приводило к образованию стеблей со стелой сложной конфигурации (например, актиностела у астероксилон ).
Под редукцией понимается уменьшение числа порядков ветвления, что приводило к упрощенному строению органа. Так, простая листовая пластинка хвощей в ходе эволюции возникла из системы многократно ветвившихся теломов , которые подвергались редукции.
Принципиально важным положением теломной теории является положение о том, что возникновение вегетативных листьев и спорофиллов происходило одновременно и независимо друг от друга, и дальнейшее их развитие шло параллельными путями. Следовательно, формирование вегетативных и спороносных побегов происходило также независимо друг от друга.
Анализ палеоботанического материала и ход развития современных примитивных растений показывают, что если не у всех, то у большинства высших растений ход становления вегетативных и спороносных органов согласуется с основными положениями теломной теории.
Псилотовидные - это группа ныне живущих высших растений, весьма близко стоящая к вымершим риниофитовым . Они представлены 1 классом Psilotopsida , порядком Psilotales , семейством Psilotaceae , которое включает всего лишь 2 рода - псилот (Psilotum) с 2 видами и тмезиптерис (Tmesipteris) с 10 видами. Ареал этих видов ограничен тропическими и частично субтропическими влажными областями обоих полушарий.
Псилотовые представляют собой травянистые многолетние растения, ведущие наземный или эпифитный образ жизни. Они имеют наземные дихотомически ветвящиеся побеги , в основном прямостоячие, у эпифитов - свешивающиеся. У псилота плоский или трехгранный стебель достигает высоты 20-100 см и обильно ветвится. У тмезиптериса размеры побегов меньше - 5-30 см; они либо совсем не ветвятся, либо ветвятся однажды дихотомически. Подземные органы псилотовых, которые подобно риниофитовым 90 называют ризомоидами , достигают длины 1 м и также дихотомически ветвятся. Они лишены каких-либо чешуй и покрыты многочисленными ризоидами с кутинизированными оболочками. Ризомоиды иногда выходят наружу и превращаются в наземные побеги .
Главное своеобразие псилотовых заключается в отсутствии корней во все периоды их жизни, что рассматривается многими учеными как первичный признак; это сближает их с риниофитовыми .
К числу примитивных признаков относится также отсутствие типичных листьев. На стеблях псилота, в его верхних частях, располагаются чешуевидные придатки, лишенные жилок, хотя листовые следы в стебле имеются. Эти чешуйки очень напоминают листья астероксилона , у которого жилки лишь подходили к основанию листа. У тмезиптериса в основании побега расположены мелкие чешуевидные придатки, тоже без жилок. Выше по стеблю они становятся крупнее и представляют собой ланцетные, низбегающие по стеблю зеленые пластинки, каждая из которых снабжена неветвящейся жилкой. В отличие от псилота листья тмезиптериса имеют в эпидерме устьица.
Таким образом, листовидные органы псилотовых подобно плауновидным имеют энационное происхождение . Но у псилота они в своем развитии остановились на уровне девонских форм, а у тмезиптериса достигли уровня современных плаунов. Функцию фотосинтеза выполняют в основном стебли.
Для надземных стеблей псилота характерно наличие эпидермы, паренхимной хлорофиллоносной коры с листовыми следами и актиностелы с 5-10 лучами ксилемы . Протоксилема сложена спиральными и кольчатыми трахеидами, а метаксилема - в основном лестничными трахеидами, но иногда встречаются даже точечные. В центре ксилемы располагается механическая ткань , выполняющая опорную функцию. У тмезиптериса, поскольку он меньших размеров и почти не ветвится, в центре располагается паренхима, т.е. имеет место сифоностела . Подземные органы построены по типу протостелы , хотя самые тонкие ризомоиды (до 1 мм в диаметре) вообще лишены проводящей системы. Паренхима ризомоидов пронизана грибными гифами , которые проникают через кутинизированные стенки ризоидов наружу и выполняют функцию поглощения воды и минеральных веществ.
псилофиты (Psilophyta), самая древняя и примитивная вымершая группа (отдел) высших растений. Характеризовались верхушечным расположением спорангиев (См. Спорангий) и равноспоровостью, отсутствием корней и листьев, дихотомическим или дихоподиальным (псевдомоноподиальным) ветвлением, примитивным анатомическим строением. Проводящая система — типичная Протостела. Протоксилема располагалась в центре ксилемы; метаксилема состояла из трахеид с кольчатыми или (реже) лестничными утолщениями. Опорные ткани отсутствовали. Р. ещё не обладали способностью ко вторичному росту (меристемы (См. Меристема) у них были только верхушечные). Спорангии примитивные, от шаровидных (диаметром около 1 мм) до продолговато-цилиндрических (длиной до 12 мм), толстостенные. Гаметофиты Р. достоверно не известны (некоторые авторы считают гаметофитами горизонтальные корневищеподобные органы — так называемые ризомоиды). Р. произрастали на влажных и болотистых местах, а также в прибрежном мелководье. Отдел Р. включает один класс — риниопсиды (Rhyniopsida) с двумя порядками — Rhyniales (семества Cooksoniaceae, Rhyniaceae, Hedeiaceae) и Psilophytales (семейство Psilophytaceae). Для порядка Rhyniales характерны дихотомическое ветвление и тонкая, слабо развитая Стела. Ксилема из трахеид с кольчатыми утолщениями. Древнейший представитель Р. — род куксония, первоначально обнаруженный в Уэльсе в отложениях конца силурийского периода (около 400 млн. лет назад). Наиболее полно изучены нижнедевонские роды — риния и отчасти хорнеофит, у которого ризомоид (вверх от него отходили стебли, вниз — многочисленные Ризоиды) был расчленён на чётковидно расположенные клубневидные сегменты, лишён проводящих тканей и целиком состоял из паренхимных клеток.Полагают, что в процессе эволюции ризомоиды Р. дали начало корням. У обоих родов стенка спорангия была многослойной, покрытой кутикулой (См. Кутикула). Хорнеофит характеризуется своеобразной спороносной полостью, которая образует купол, сводообразно покрывающий центральную колонку стерильной ткани, представляющей собой продолжение флоэмы стебля. Этим хорнеофит напоминает современный Сфагнум. В семейства риниевых включают также род тениокрада, многие виды которого образовывали подводные заросли в среднем и верхнем девоне. В отдельное семейства хедеевых иногда выделяют нижнедевонские роды хедея и яравия. Нижнедевонский род сциадофит, обычно выделяемый в отдельное семейства сциадофитовых, — небольшое растение, состоявшее из розетки простых или слабо дихотомированных тонких стеблей со стелой. Для порядка Psilophytales характерны дихоподиальное ветвление и более сильно развитая стела. У наиболее известного рода — псилофит (из нижнедевонских отложений в Восточной Канаде) — неравно развитые ветви образовывали ложную главную ось дихоподия с более тонкими боковыми ветвями: стебель был окружен кутинизированной эпидермой с устьицами; поверхность стебля была голая или покрыта шипами длиной 2—2,5 мм, концы которых дисковидно расширялись, что, вероятно, указывало на их секреторную роль. Спорангии раскрывались продольной трещиной. К псилофиту близки нижнедевонские роды тримерофит и пертика.
Изучение строения Р. и их эволюционных взаимоотношений имеет большое значение для эволюционной морфологии и филогении высших растений. По-видимому, первоначальным органом Спорофита высших растений был дихотомически ветвящийся стебель с верхушечными спорангиями; корни и листья произошли позже спорангия и стебля. Есть все основания считать Р. исходной предковой группой, от которой произошли мохообразные, плауновидные, хвощевидные и папоротники. Согласно другой точке зрения, мохообразные и плауновидные имеют лишь общее происхождение с Р.
Лит.: Основы палеонтологии. Водоросли, мохообразные, псилофитовые, плауновидные, членистостебельные, папоротники, М., 1963; Traite de paleobotanique, t. 2, Bryophyta. Psilophyta. Lycophyta, P., 1967.
К отделу принадлежат самые древние из известных высших растений (рис. 2). Названия рода Риния (Rhynia) происходит от деревни Райни (недалеко от г. Абердина) в Шотландии, близ которой были обнаружены остатки этих примитивных растений в кремнистых породах врачом У. Макки в 1912 г. Эти отложения представляют собой пропитанные растворами кремниевой кислоты окаменевшие девонские торфа. У. Макки в свободное от медицинской практики время увлекался геологией. Свои материалы по ископаемым таксонам он передал шотландскому палеоботанику Р. Кидстону, который совместно с профессором Манчестерского университета У. Лангом опубликовали в 1917–1921 гг. результаты изучения растений ископаемой флоры. Они заложили начало систематическому изучению группы древнейших и наиболее примитивных высших растений.
ан



7б



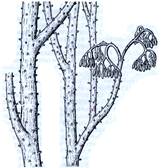
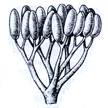
Рис. 2. Риниеобразные (Rhyniophyta): 1 – риния большая (Rhynia major), (м – мезомы, р – ризоиды, рз – ризомоид, ст – стерильные теломы, фт – фертильные теломы); 2 – хорнеофитон (Horneophyton), (с – спорангии); 3а,3б – лионофитон (Lyonophyton), (3а – реконструкция гаметофита; 3б – обоеполый гаметангиофор: ан – антеридий, арх – архегоний); 4 – сциадофитон (Sciadophyton) с однополыми гаметангиофорами; 5 – спорангии хедеи (Hedeia); – синангий ярравии (Jarravia); 7а,7б – пертика (Pertica), (7а – реконструкция спорофита, 7б – сильно разветвленные спороносные оси); 8 – фрагмент псилофита первичного (Psilophyton princeps); 9 – зостеролист (Zosterophyllum); 10 – госслингия (Gosslingia); 11 – савдония (Sawdonia).
Древнейшие наземные растения раньше объединяли в один таксон под названием Псилофиты. Теперь разными исследователями предлагается 3 таксона на уровнях отделов. Многие объединяют их в отдел Риниеобразные – Rhyniophyta, насчитывающий около 20 родов и большое число видов. Единство их подтверждается рядом важных общих признаков, отражающих начальную ступень в эволюции высших растений. Единство их подкрепляемся также наличием родов, которые сочетают признаки трех отмеченных групп.
Отдел включает самые древние достоверные и по мнению большинства исследователей – самые примитивные травянистые высшие растения. Они появились приблизительно 430 млн. лет назад в силурийском периоде палеозойской эры. Геологическая история отдела заканчивается в девоне (около 370 млн. лет назад). Они дали начало другим группам высших растений.
Вегетативное тело спорофита риниофотов представлено безлистными цилиндрическими дихотомически ветвящимися, первично васкуляризованными осевыми структурами. Наземные оси выполняли в первую очередь функцию воздушного питания. Их конечные ответвления – теломы (от греч. telos – конец), а соединяющие их части, расположенные между точками ветвления – мезомы (от греч. mesos – средний). Одни теломы были стерильными, другие – фертильными и заканчивались спорангиями (рис 2). Стерильные теломы осуществляли только функцию фотосинтеза. Горизонтально расположенные на поверхности или в почве влажных местообитаний теломы получили название ризомоиды (т. е. корневищеподобные). От них отходили многочисленные простые (несептированные) одноклеточные ризоиды, осуществляя функцию минерального питания. Ризомоид – прототип корня, ризоиды – прототипы корневых волосков. Наличием проводящих пучков теломы принципиально отличались от структур водорослей.
Таким образом, теломы, мезомы и ризомоиды были первыми конструктивными элементами спорофита наземных растений. Возникновение этих элементарных органов было крупным ароморфозом в эволюции растительного мира. Это сильно повысило уровень организации растений, увеличило их пластичность и открыло более широкие возможности дальнейшего эволюционного развития.
Ветвление надземных вертикальных осей спорофита риниофитов характеризовалось равной и неравной дихотомией. Вследствие резко выраженной анизофилии, ветвление приобретало характер псевдомоноподиального. Поверхность надземных осей была голой или покрыта многочисленными выростами в виде энаций (в т.ч. шипиками, зубчиками).
Риниофиты – первые растения суши. Как наземные растения, в отличие от водорослей, они были покрыты типичной эпидермой, характеризовались наличием проводящей системы. Наружные стенки клеток эпидермы утолщены и покрыты слоем кутикулы. Имелись немногочисленные просто устроенные устьица (доказательство их наземного существования) – с двумя замыкающими клетками и устричной щелью. Они были лишены побочных клеток. Наличие устьиц в первую очередь отличает эпидерму высшего растения от наружного слоя клеток многоклеточных талломов (слоевищ) водорослей. Проводящая система слабо развита и построена по типу гаплостели, которая была началом развития всех остальных типов стели. Ксилема состояло из трахеид с кольчатыми, спиральными или реже лестничными утолщениями. Риниофты не обладали способностью к вторичному росту (камбий отсутствовал, как и перицикл). Меристема только верхушечная. В отличие от большинства сосудистых растений отсутствовали также и механические (опорные) ткани. Их функции выполняла толстая кора.
Спорангии всегда непосредственно прикреплялись к осям, или являлись их продолжением. У большинства более высоко организованных высших растений они прикрепляются к специализированным придаткам.
Спорангии риниофитов располагались верхушечно (терминально) на теломах или в боковом положении, одиночно (риния, куксония, хорнеофитон – Horneophyton, госслингия – Gosslingia) или группами (пертика – Pertica, тримерофит – Trimerophyton, зостеролист – Zosterophyllum). Они также отличались достаточно примитивным строением. Имели толстую многослойную стенку (спорангии эуспорангиатного типа), споры многочисленные, одинакового строения (растения морфологически равноспоровые). Спорангии различались по величине и форме. У наиболее примитивных видов они мало отличались от верхушек вегетативных осей. У куксонии они шаровидные, диаметром 1 мм. У ринии – продолговато-цилиндрические, длиной до 12 мм и толщиной до 4 мм. Спорангии не имели специальных приспособлений для вскрывания или они были крайне примитивно плана. У одних видов споры освобождались после разрушения стенки спорангия. У других видов спорангии вскрывались трещиной, располагавшейся продольно, поперечно (близ основания или верхушки спорангия), апикальной порой (у хорнеофитона). Нередко трещины проходили по определенным бороздам. В спорангий хорнеофитона глубоко вдавалась колонка, на которой располагался куполообразный спорангий. Интересно, но уже среди риниофитов наблюдалась тенденция к сближению спорангиев (хедея – Hedeia) и в дальнейшем к их срастанию в синангии (ярравия – Jarravia). Ярравия – нижнедевонское растение. В дальнейшем синангии будут неоднократно формироваться в различных линиях эволюции высших растений. Таким образом, спороносные структуры риниофитов, несмотря на примитивность их организации, характеризуются разнообразием морфологического строения. Эта особенность характерна для представителей большинства отделов высших растений и широко используется при классификации.
В спорангиях развивались многочисленные споры, одинакового морфологического строения (риниофиты были равноспоровыми растениями). Строение толстой оболочки спор – типичное для высших растений. Они были снабжены трехлучевым тетрадным рубцом.
Гаметофиты ринофитов существовали независимо от спорфитов и были свободно живущими растениями, однодомными (антеридии и архегонии развивались на одном растении). Под родовым названием лионофитон (Lyonophyton) описаны гаметофиты с обоеполыми гаметангиофорами. Предполагается, что они принадлежали хорнеофитону. На вершине некоторых неразветвленных осей имелось апикальное чашевидное расширение с лопастным краем. В центре чаши формировался небольшой выступ (архегониефор), к которому прикреплялись архегонии с сильно вытянутыми шейками. Булавовидные антеридии располагались по периферии чаши на внутренних стенках. Выявлены также гаметофиты с раздельнополыми гамитангиофорами (сциадофитон – Sciadophyton).
Произрастали риниофиты во влажных и болотистых местообитаниях вокруг морских и материковых водных бассейнов, а также в прибрежных мелководьях. Иногда они образовывали значительные заросли, являлись ландшафтными растениями. Возможно среди них были сухопутные растения. Некоторые были водными растениями с выступающими над водой спороносными частями.
Наиболее древней из достоверных находок высших растений является куксония. Более ранние риниофиты описаны только по спорам. Уже древнейшие силурийские риниофиты прошли длительный путь эволюционного развития. Это были довольно сложными, специализированными растениями разнообразного строения, приспособленными к жизни в наземных условиях.
Произошли они от зеленых водорослей. Наиболее древние формы еще сохраняли сходство с водорослеообразными предками. В частности, очень сходный характер их ветвления.
Многообразие признаков риниофитов явилось основой для дальнейших эволюционных направлений. Они дали начало многим независимым группам высших растений. В первую очередь плунообразным, хвощеобразным, псилотообразным и папоротникообразным, а также мохообразным (одна из гипотез). Изучение строения первенцев наземной суши имеет большое значение для эволюционной морфологии и филогении всего подцарства высших растений. Установлено, что первым органом спорофита высших растений был дихотомически разветвленный радиальный телом, строение которого, как отмечено, характерно для сухопутных растений (наличие проводящей системы, появление эпидермы, устьиц и т. д.). В ходе дальнейшего приспособления к наземной жизни от них произошли растения, обладавшие побегами и корнями.
В ходе эволюции растений из теломов формировались не только листья, но и другие органы высших растений. Например, срастание теломов, расположенных в разных плоскостях, привело к формированию стеблей с более сложным строением стели, чем протостела риниофитов. В процессе эволюции наземных растений наблюдались не только явления усложнения организации, но и явления редукции (уменьшение порядков ветвления, что приводило к упрощению строения органа). Таким путем сформировалась редуцированная листовая пластинка современных хвощей. Регрессивное развитие спорофита в целом наблюдалось у мохообразных. Одним из доказательств является наличие на спорофите некоторых печеночных и антоцеротовых мхов рудиментарных устьиц. При освоении растениями менее влажных местообитаний в процессе адаптации и последующей эволюции размеры листьев уменьшались из-за ослабления их роста. Появляются различные приспособления для ограничения транспирации. Это приводило к расширению адаптационных возможностей растений.
Некоторые листья стали выполнять функцию защиты зоны нарастания стебля и молодых зачатков листьев. Это обусловило появление почек. Сформировался побег (осевая часть – стебель, несущий листья и почки). Дифференциация и усложнение надземных органов, приведшая к формированию побегов, сопровождалась образованием корней в эволюции большинства высших растений для укрепления растения в почве и поглощения воды с растворенными минеральными веществами.
Спорофиллы и трофофиллы (вегетативные листья) согласно теломной теории возникают одновременно и независимо друг от друга. Дальнейшие их развитие шло параллельными путями. Следовательно, независимо друг от друга происходило также формирование вегетативных и спороносных побегов (стробилов). В процессе дальнейшей эволюции высших растений подземные органы совершенствовались, приобретая новые функции, видоизменялись, формируя метаморфозы (корневища, клубни, луковицы и др.). Возникают также новые ароморфозы (семена, цветки, плоды).
Как отмечено, со времени открытия, систематическое положение, классификация, таксономический ранг, объем единиц классификации риниофитов не однократно пересматривались. Это касается большинства групп ископаемых растений благодаря накоплению нового палеонтологического материала.
Некоторые исследователи разделяют риниофиты на 2 или 3 самостоятельных отдела, что не вполне оправдано. Все они связаны рядом важных общих признаков, отличающих их от более организованных высших споровых растений (плаунов, хвощей, папоротников). Они полностью лишены вторичных тканей, корней и побегов; оси всегда протостелические, стела нерасчлененная, спорангии прикрепляются непосредственно к наземным осям (терминально или латерально). Сходство прослеживается и по другим признакам (близость строения спор, ризоидов). Кроме того, существуют так называемые синтетические таксоны (тениокрада – Taeniocrada, куксония, стеганотека – Steganotheca, нотия – Nothia, реналия – Renalia и др.) в которых комбинируются признаки двух или даже трех указанных групп. Они, по-видимому, должны располагаться вместе, в составе одного отдела, в основе филогенетической системы высших растений, подчеркивая их монофилетическое происхождение.
Нередко отдел Риниеобразные условно разделяют на 2 класса – Риниеподобные и Зостеролистоподобные.
Читайте также: