
Взаимодействие неаллельных генов реферат
Обновлено: 05.07.2024
Взаимодействия аллельных генов. Явление, когда за один признак отвечает несколько генов (аллелей), называется взаимодействием генов. Если это аллели одного и того же гена, то такие взаимодействия называются аллельными, а в случае аллелей разных генов — неаллельными.
Выделяют следующие основные типы аллельных взаимодействий: доминирование, неполное доминирование, сверхдоминирование и кодоминирование, плейотролия.
Доминирование — тип взаимодействия двух аллелей одного гена, когда один из них полностью исключает проявление действия другого. Такое явление возможно при следующих условиях: 1) доминантный аллель в гетерозиготном состоянии обеспечивает синтез продуктов, достаточный для проявления признака такого же качества, как и в состоянии доминантной гомозиготы у родительской формы; 2) рецессивный аллель совсем неактивен, либо продукты его активности не взаимодействуют с продуктами активности доминантного аллеля.
Примерами такого взаимодействия аллельных генов может служить доминирование пурпурной окраски цветков гороха над белой, гладкой формы семян над морщинистой, темных волос над светлыми, карих глаз над голубыми у человека и т. д.
Неполное доминирование, или промежуточный характер наследования, наблюдается в том случае, когда фенотип гибрида (гетерозиготы) отличается от фенотипа обеих родительских гомозигот, т. е. выражение признака оказывается промежуточным, с большим или меньшим уклонением в сторону одного или другого родителя. Механизм этого явления состоит в том, что рецессивный аллель неактивен, а степень активности доминантного аллеля недостаточна для того, чтобы обеспечить нужный уровень проявления доминантного признака.
Неполное доминирование оказалось широко распространенным явлением. Оно наблюдается в наследовании курчавости волос у человека, масти крупного рогатого скота, окраски оперения у кур, многих других морфологических и физиологических признаков у растений, животных и человека.
Сверхдоминирование — более сильное проявление признака у гетерозиготной особи (Аа), чем у любой из гомозигот (АА и аа). Предполагается, что это явление лежит в основе гетерозиса.
Кодоминирвание— участие обоих аллелей в определении признака у гетерозиготной особи. Ярким и хорошо изученным примером кодоминирования может служить наследование IV группы крови у человека (группа АВ).
Взаимодействия неаллельных генов.
Среди них можно выделить следующие основные типы: комплементарность, эпистаз и полимерию.
Комплементарность — взаимодействие неаллельных генов, которые обусловливают развитие нового признака, отсутствующего у родителей. Примером комплементарного действия у человека могут служить случаи, когда у глухих родителей рождаются дети с нормальным слухом. Развитие нормального слуха находится под генетическим контролем десятков различных неаллельных генов, гомозиготное рецессивное состояние одного из которых может приводить к одной из форм наследственной глухоты. Таких форм у человека известно более 30. Если один из родителей является гомозиготой по рецессивному гену аа, а другой — гомозиготой по другому рецессивному гену bb, то все их дети будут двойными гетерозиготами и, следовательно, слышащими, поскольку доминантные аллели будут взаимно дополнять друг друга. Таким образом формируется новый по отношению к родителям признак — нормальный слух.
Эпистаз (от греч. ерi — над + stasis — препятствие) — взаимодействие неаллельных генов, при котором наблюдается подавление проявления одного гена действием другого, неаллельного гена.
Подавляющий ген называется геном- супрессором, а подавляемый — гипостатическим геном. По-видимому, действие гена- супрессора на подавляемый ген сходно с принципом доминантность — рецессивность. Но существенное различие заключается в том, что эти гены не являются аллельными, т.е. расположены в негомологичных хромосомах или занимают различные локусы в гомологичных.
Различают доминантный и рецессивный эпистаз. При доминантном эпистазе доминантный аллель гена- супрессора подавляет проявление доминантного аллеля другого гипостатического гена. При рецессивном эпистазе, или криптомерии, рецессивный аллель гена-супрессора, будучи в гомозиготном состоянии, не дает проявиться доминантной или рецессивным аллелям других генов.
Примером рецессивного эпистаза у человека может служить так называемый бомбейский фенотип, когда индивид, имеющий доминантный аллель группы крови системы АВ0 (например, аллель В, определяющий принадлежность человека к III или IV группе), идентифицируется в реакции агглютинации как человек с 0(I). Это состояние возникает в результате того, что данный индивид является рецессивной гомозиготой (hh) по другой, нежели система АВ0, генетической системе Hh. Для реализации аллелей IА и IЕ необходимо присутствие хотя бы одного доминантного I аллеля Н.
Полимерия (от греч. роlуs — много + meros — часть) — вид взаимодействия, когда эффекты нескольких неаллельных генов, определяющих один и тот же признак, примерно одинаковы. Подобные признаки получили название количественных, или полимерных признаков. Как правило, степень проявления полимерных признаков зависит от числа доминантных генов. Наследование полимерных признаков было впервые описано шведским генетиком Г. Нильсон-Эле в 1908 г.
Если вам нужна помощь в написании работы, то рекомендуем обратиться к профессионалам. Более 70 000 авторов готовы помочь вам прямо сейчас. Бесплатные корректировки и доработки. Узнайте стоимость своей работы.
Анализ особенностей расщепления показал, что в данном случае окраску зерен определяют два доминантных аллеля двух различных генов, а сочетания их рецессивных аллелей определяют отсутствие окраски. Поскольку полимерные гены имеют одно-направленное действие, их, как правило, обозначают одинаковыми буквами. Таким образом, исходные родительские формы имели генотипы А]А1А2А2 и а1а1а2а2. Наличие всех четырех доминантных аллелей определяло самую интенсивную окраску, трех доминантных аллелей (типа А1А1А2А2) — менее интенсивную окраску и т.д.
Примером полимерного наследования у человека является наследование окраски кожных покровов. В браке индивида негроидной расы (коренного жителя Африки) с черной окраской кожи и представителем европеоидной расы с белой кожей дети рождаются с промежуточным цветом кожи (мулаты). В браке двух мулатов потомки могут обладать любой окраской кожи: от черной до бе-лой, поскольку пигментация кожи обусловлена действием трех или четырех неаллельных генов. Влияние каждого из этих генов на окраску кожи примерно одинаково.
Полимерное наследование характерно для так называемых количественных признаков, таких, как рост, вес, окраска кожных покровов, скорость протекания биохимических реакций, артериальное давление, содержание сахара в крови, особенности нервной системы, уровень интеллекта, и многих других, которые нельзя разложить на четкие фенотипические классы. Чем большее число неаллельных генов контролируют развитие количественного признака, тем менее заметны переходы между фенотипическими классами.
- Для учеников 1-11 классов и дошкольников
- Бесплатные сертификаты учителям и участникам
МУНИЦИПАЛЬНОЕ БЮДЖЕТНОЕ УЧРЕЖДЕНИЕ
ДОПОЛНИТЕЛЬНОГО ОБРАЗОВАНИЯ
ИЛЛЮСТРАТИВНАЯ ЛЕКЦИЯ ПО ТЕМЕ
педагог дополнительного образования
Люминарская Наталья Александровна
Программа является модифицированной с углублённым уровнем изучения биологии.
Новизна и отличительная особенность данной программы состоит в том, что её содержание соответствует прогаммам курсов подготовки по биологии, организуемыми высшими и средними профессиональными учебными заведениями для абитуриентов. Кроме того, в процессе реализации данной программы для закрепления знаний учащихся используется множество тестов, что помогает им приобрести практические навыки, необходимые для успешной сдачи экзаменов в форме ЕГЭ.
Цель занятия : Изучить основные типы взаимодействия генов, приобрести знания и практические навыки, необходимых для успешного прохождения тестирования по биологии в школе.
- углубить знания о закономерностях наследственности;
- расширить представление о взаимодействии генов;
- продолжить формирование навыков составления схемы скрещивания;
- сформировать представление о системном характере генотипа.
Оборудование и материалы : интерактивная доска, рисунки, задачи по генетике – раздаточный материал, тетради, ручки, карандаши.
Методы обучения : лекционный, наглядно-демонстрационный, беседа.
План занятия :
Проверка домашнего задания.
Актуализация знаний по новой теме.
Изучение нового материала. Объяснение с использованием презентации.
Закрепление изученного материала (решение задач).
Подведение итогов. Домашнее задание.
Ход занятия :
1.Организационный момент.
2. Проверка домашнего задания.
Актуализация знаний.
Давайте вспомним определение генетических терминов, необходимых для изучения сегодняшней темы занятия.
Что такое ген? (Участок хромосомы, несущий информацию о первичной структуре одного белка)
Что какое локус? (место положения гена в хромосоме)
Что такое аллели? ( различные формы одного и того же гена, располагающиеся в одинаковых участках гомологичных хромосом и определяющие развитие одного признака).
Что такое аллельные гены? ( гены, расположенные в одних и тех же участках (локусах) гомологичных хромосом и определяющие развитие одного признака ).
Что такое неаллельные гены? (гены, расположенные в разных участках гомологичных хромосом или в негомологичных хромосомах и определяющие развитие разных признаков).
Что такое множественный аллелизм? ( явление существования более двух аллельных форм гена, имеющих различные проявления в фенотипе ( трёхаллельный ген, определяющий у человека группу крови А, В, О) .
Освоение нового материала.
Тема даётся в форме иллюстративной лекции. Материал объясняется частями с использованием презентации, проектора, подборки распечатанных рисунков, которые раздаются каждому ученику перед началом занятия. Изученный материал записывается в тетради под диктовку.
В он тогенезе действуют не отдельные гены, а весь генотип .
Генотип — это исторически сложившаяся целостная система из взаимодействующих между собой генов. Точнее, взаимодействуют не сами гены (участки молекул ДНК), а образуемые на их основе продукты (РНК и белки).
Свойства генов и особенности их проявления в при знаках :
Ген дискретен в своем действии, т. е. обособлен в своей актив ности от других генов.
Ген специфичен в своем проявлении, т. е. отвечает за строю определённый признак или свойство организма.
Ген может усиливать степень про явления признака при увеличении числа доминантных алле лей (дозы гена).
Один ген может влиять на развитие разных признаков — это множественное, или плейотропное, действие гена.
Разные гены могут оказывать одинаковое действие на разви тие одного и того же признака (часто количественных призна ков) — это множественные гены, или полигены.
Ген может взаимодействовать с другими генами , что приво дит к появлению новых признаков. Такое взаимодействие осу ществляется опосредованно — через синтезированные под их контролем продукты своих реакций.
Действие гена может быть модифицировано изменением его местоположения в хромосоме (эффект положения) или воз действием различных факторов внешней среды.
ВЗАИМОДЕЙСТВИЯ АЛЛЕЛЬНЫХ ГЕНОВ.
Явление, когда за один признак отвечает несколько генов (аллелей), называется взаи модействием генов .
Если это аллели одного и того же гена, то такие взаимодействия называются аллельными , а в случае аллелей разных генов — неаллельными .
Выделяют следующие основные типы взаимодей ствия аллельных генов :
● Полное доминирование — явление, когда доминантный ген полностью подавляет работу рецессивного гена , в результате чего развивается доминантный признак.
Примерами такого взаимодействия аллельных генов может служить доминирование пурпурной окраски цветков гороха над белой, гладкой формы семян над морщинистой, темных волос над светлыми, карих глаз над голубыми у человека и т. д.
● Неполное доминирование — явление, когда доминантный ген не полностью подавляет работу рецессивного гена , в результате чего развивается промежуточный признак . Примером неполного доминирования является наследование окраски цветков у растений ночной красавицы Гомозиготные растения имеют либо красные (АА), либо белые (аа) цветки, а гетерозиготные (Аа) — розовые. ● Сверхдоминирование — более сильное проявление призна ка у гетерозиготной особи (Аа) , чем у любой из гомозигот (АА и аа) . Это явление лежит в основе гетеро зиса.
При межпородных или межвидовых скрещиваниях у гибридов первого поколения происходит особенно мощное развитие и повышение жизнеспособности (мул – гибрид кобылы и осла, бройлерные цыплята).
● Кодоминирование (независимое проявление) — явление, когда в формировании признака у гетерозиготного организма участвуют одновременно два аллельных гена .
Наблюдается при наследовании групп крови у человека. Гены, обусловливающие группы крови А и В, являются кодоминантными по отношению друг к другу, и оба доминантны по отношению к гену, определяющему группу крови О.
J А (в гомозиготном состоянии контролирует II группу крови - А), а J В (в гомозиготном со стоянии контролирует III группу крови – В). Аллели J А и J В ра ботают в гетерозиготе как бы независимо друг от друга, что названо кодоминированием
ГЕНЕТИКА КРОВИ.
У человека по генотипу 6 групп крови, а по физиологии — 4.
► I группа (0) : J 0 J 0 II группа (А): J А J А , J А J 0 III группа (В): J В J В , J В J 0
IV группа (АВ): J А J В
Кроме того, кровь разных людей может отличать резус-фактором. Кровь может иметь положительный резус-фактор ( Rh + ) или отрицательный резус-фактор ( Rh - ) . Резус-фактор крови определяет ген R . R + дает информацию о выработке белка (резус положительный белок), а ген R - — не дает. Первый ген доминирует над вторым. Если Rh + кровь перелить человеку с R - кровью, то у него образуются специфические агглютинины (антитела), и повторное введение такой крови вызовет агглютинацию. Когда у резус отрицательной женщины развивается плод, унаследовавший у отца положительный резус, может возникнуть резус-конфликт.
Пример наследования групп крови иллюстрирует и проявление множественного аллелизма.
Есть гены, имеющие десятки аллелей. Все аллели одного гена получили название серии мно жественных аллелей.
ВЗАИМОДЕЙСТВИЯ НЕАЛЛЕЛЬНЫХ ГЕНОВ.
Взаимодействие неаллельных генов приводит к появлению в потомстве дигетерозиготы необычного расщепления по фено типу: 9:3:4, 9: 7, 9:6:1, 13:3, 12:3:1, 15:1 , т.е. модификации общей менделевской формулы 9:3:3:1.
► Можно выделить следующие основные типы взаимодействия
неаллельных генов :
► Комплементарным , или дополнительным, называется такое взаимодействие неаллельных доминантных генов, в результате ко торого появляется признак, отсутствующий у обоих родителей .
● Комплементарность можно рассмотреть на примере наследования окраски цветков у душистого горошка : существуют расы душистого горшка с пурпурными цветками и расы — с белыми цветками. При скрещивании двух белоцветковых сортов различного происхождения ( ААвв и ааВВ) наблюдается необычный характер наследования – появляются пурпурноокрашенные цветки.
Гибрид первого поколения с двумя доминантными генами (А и В) получил биохимическую основу для выработки пурпурного пигмента антоциана, в то время как поодиночке ни ген А, ни ген В не обеспечивали синтез этого пигмента . В остальных случаях (ааВ- и А-вв) цветки у растения белые и фенотипически не отличаются от сорта аавв. Рецессивные гены в гомозиготном состоянии подавляют проявление доминантных аллелей, т. е. блокируют синтез пигмента.
► Кооперация – появление новообразований при совместном действии двух доминантных неаллельных генов, когда в гомозиготном или в гетерозиготном состоянии развивается новый признак, отсутствующий у родительских форм. При таком взаимодействии во втором поколении ( F 2 ) возможно следующее расщепление – 9 : 3 : 3 : 1
Кооперацией является ● наследование формы гребня у кур :
РРгг, Рргг — гороховидный рр RR , рр Rr — розовидный
РР RR , Рр RR , Рр Rr , РР Rr — ореховидный рр rr — простой .
► Эпистаз — это такой тип взаимодействия генов, при котором один ген (как доминантный, так и рецессивный) подавляет действие другого неаллельного.
Гены, подавляющие действие других генов, называются эпистатическими, ингибиторами или супрессорами.
Различают ● доминантный эпистаз , когда действие одного доминантного гена подавляется другим доминантным геном (А > В) , и ● рецессивный эпистаз , когда рецессивные гены в го мозиготном состоянии оказывают подавляющее действие на доминантный ген или рецессивные гены из другой пары ал лелей (аа > В, аа> вв).
Примером • доминантного эпистаза служит наследование окраски оперения у кур, где ген С контролирует наличие пиг мента, с — отсутствие его; ген I (ингибитор) из другой пары хромосом подавляет действие гена С, ген i не оказывает тако го действия:
• Двойной рецессивный эпистаз – взаимное подавление рецессивными генами, находящимися в гомозиготном состоянии, доминантных аллелей рассмотрен на примере комплементарного действия генов при скрещивании белоцветковых ( Аавв и ааВВ ) растений душистого горошка. В данном примере вв > А и аа > В
► Полимерия — формирование признака несколькими парами неаллельных генов, обладающих одинаковым действием .
Такие гены называются полимерными . Полимерные гены (полигены) обычно обозначают одной буквой с нумерацией генов (А 1 , А 2 , А 3 ).
Полимерные гены могут действовать по типу
● кумулятивной (накопительной) полимерии : степень выраженности признака зависит от числа доминантных генов, ответственных за его появление. Действие таких генов суммируется, а фенотипическое проявление признака тем сильнее, чем больше доминантных генов участвуют в его контролировании.
Накопительную полимерию можно рассмотреть на примере наследования окраски зерна у пшеницы .
У зёрен пшеницы красный цвет (наиболее интенсивный) определяется генами А 1 А 2 А 3 .
Чем больше доминантных аллелей присутствуют в генотипе растения, тем более тёмная окраска зерна:
А 1 А 1 А 2 А 2 А 3 Аз — тёмно-красная окраска зерна
А 1 а 1 А 2 а 2 А 3 а 3 — насыщенно-красная
А 1 А 1 а 2 а 2 А 3 а 3 — умеренно-красная
А 1 а 1 а 2 а 2 а 3 а 3 — бледно-красная (самая светлая)
а 1 а 1 а 2 а 2 а 3 а 3 ─ белая.
● Некумулятивная (ненакопительная) полимерия .
Гены с однозначным действием могут определять и качественные признаки. Примером может служить наследование форм стручочков с семенами у пастушьей сумки:
Р: а 1 а 1 а 2 а 2 х А 1 А 1 А 2 А 2
овальная треугольная
G : а 1 а 2 , А 1 А 2
F 1 (Р): А 1 а 1 А 2 а 2
треугольная
F 2: А- : а 1 а 1 а 2 а 2
15/16 треугольная 1/16 овальная
► Плейотропия (множественный аллелизм) — влияние одного гена на формирование нескольких признаков.
► Модифицирующее действие генов — предполагается наличие генов в генотипах организмов, которые сами не определяют развитие признаков, но изменяют проявление других генов , т. е. в зависимости от присутствия в генотипе тех или иных генов-модификаторов основной ген, определяющий какой-либо признак, проявится различным образом.
● Доминантные гены не всегда проявляются в виде признаков или свойств организма . При этом в случае проявления доминантного гена, признак может быть развит в различной степени.
Вероятность реализации гена в фенотипе Н. В. Тимофеев-Ресовский (в 1927 г.) предложил называть пенетрантностью , а степень его влияния на признак — экспрессивностью .
Пенетрантность измеряется процентом особей, у которых данный ген проявился в виде признака или свойства.
Экспрессивность измеряется степенью выраженности признака, на который влияет данный ген.
Гены обладают • полной пенетрантностью , если у 100% особей признак проявляется. Доминантный ген полидактилии проявляется не у каждого его обладател (• неполная пенетрантность ).
При этом проявляется он у разных людей в различной степени (экспрессивность): могут развиваться дополнительные пальцы у одних людей на руках, у других — на ногах, или только на одной руке или ноге.
Обычно гены с неполной пенетрантностью обладают и различной экспрессивностью.
ЛИТЕРАТУРА .
ПЕРЕЧЕНЬ ПРИЛОЖЕНИЙ
Приложение 1. Рисунки по теме.
Приложение 2. Задачи.
План:
Введение.
1. Проявление действия гена.
2. Виды взаимодействия аллельных генов: полное доминирование, неполное доминирование, кодоминирование.
3. Виды взаимодействия неаллельных генов:Комплементарность, эпистаз, полимерия.
4. Плейотропное действие генов.
5. Влияние факторов внешней среды на действие генов.
Выводы.
Список использованной литературы.
1. Проявление действия гена.
Ген как единица наследственности, детерминирующая признаки, или свойства, организма, имеет определенные характеристики:
1) в своем действии ген дискретен; он определяет присутствие или отсутствие отдельной биохимическойреакции, степень развития или подавления определенного признака организма;
2) ген действует градуально: накопление дозы его в соматических клетках может приводить к усилению или ослаблению проявления признака;
3) каждый ген специфичен в своем действии, т.е. отвечает за синтез первичной структуры белковой молекулы;
4) ген может опосредованно воздействовать на течение разных реакций и развитие многих признаковорганизма, т.е. действовать множественно; это явление носит название множественного, или плейотропного эффекта гена;
5) разные гены, находящиеся в различных парах хромосом, могут действовать однозначным образом на развитие одного и того же органа или свойства организма, усиливая или ослабляя его; это так называемые множественные, или полимерные, гены (полигены):
6) ген вступает во взаимодействиес другими генами, и в силу этого его эффект может изменяться;
7) проявление действия гена зависит от факторов внешней среды;
8) гены взаимодействуют теми продуктами, которые они детерминируют и контролируют в процессе их синтеза.
Как мы увидим дальше, гены действуют на двух уровнях: на уровне самой генетической системы, определяя состояние генов и их работу, скорость репликации ДНК,стабильность и изменчивость генов, и на уровне работы клеток в системе целого организма.
Прежде чем перейти к рассмотрению указанных особенностей действия генов, необходимо условиться о системе их обозначения. До сих пор для обозначения гена мы пользовались любыми, большей частью начальными, буквами латинского алфавита. Поскольку в дальнейшем нам придется касаться.
♦ Свойства генов и особенности их проявления в признаках:
■ ген дискретен в своем действии, т.е. обособлен в своей активности от других генов;
■ один ген отвечает за проявление одного строго определенного признака или нескольких признаков (плейотропия);
■ один признак может быть результатом действия нескольких генов (аллельных или неаллельных);
■ ген может усиливать степень проявления признака при увеличении числа его доминантных аллелей;
■ ген может взаимодействовать с другими генами; это приводит к появлению новых признаков;
■ изменение положения гена в хромосоме или влияние факторов внешней среды могут модифицировать его проявление в признаках;
■ ген обладает способностью к мутациям.
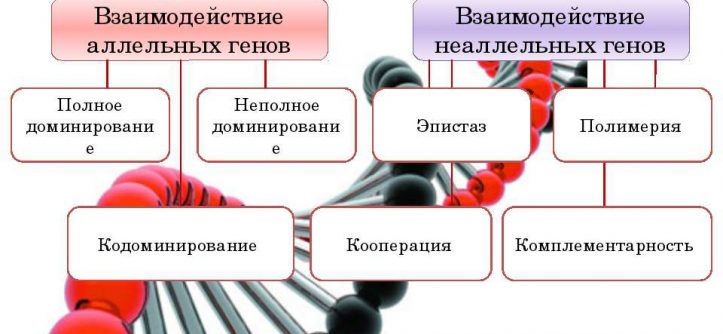
Взаимодействие аллельных генов
Взаимодействие генов — явление, когда за один признак отвечает несколько генов (или аллелей).
Аллельное взаимодействие — это взаимодействие аллелей одного и того же гена (за признак отвечает несколько аллелей одного и того же гена).
❖ Типы аллельных взаимодействий:
■ доминирование,
■ неполное доминирование,
■ сверхдоминирование,
■ кодоминирование.
Доминирование — тип взаимодействия двух аллелей одного гена, когда один (доминантный) из них полностью исключает действие другого (рецессивного). Примеры: доминирование у человека темных волос над светлыми, карих глаз над голубыми.
Неполное доминирование — степень активности доминантного аллеля недостаточна для того, чтобы полностью подавить действие рецессивного аллеля и обеспечить полное проявление доминантного признака.
■ В этом случае у гетерозигот формируется промежуточный (по отношению к родительским признакам) признак — имеет место промежуточный характер наследования. Этот признак будет наблюдаться у гибридов первого поколения и гетерозигот второго поколения. Во втором поколении расщепление по фенотипу и генотипу оказывается одинаковым 1:2:1 (одну часть составляет доминантная гомозигота АА с выраженным доминантным признаком, две части составляет гетерозигота Аа с промежуточным признаком и одну часть составляет гомозигота аа с рецессивным признаком).

■ Примеры неполного доминирования: наследование формы (курчавости) волос у человека, масти крупного рогатого скота, окраски цветков у растения ночная красавица (см. таблицу).
Сверхдоминирование — более сильное проявление признака у гетерозиготной особи (Аа), чем у любой из гомозигот (АА и аа).
Кодоминирование — оба аллеля равноценны, не подавляют друг друга и участвуют в определении признака у гетерозиготной особи. Пример: наследование IV группы крови у человека, которая детерминируется одновременным присутствием в генотипе двух кодоминантных генов I А и I в . Первый из этих генов детерминирует синтез в эритроцитах белка-антигена А, второй — синтез белка-антигена В; наличие обоих этих генов в генотипе приводит к тому, что у людей с IV группой крови эритроциты содержат как белок-антиген А, так и белок-антиген В.
Взаимодействие неаллельных генов
Неаллельное (или межаллельное) взаимодействие — это взаимодействие аллелей разных генов, т.е. генов, располагающихся в негомологичных хромосомах или разных локусах гомологичных хромосом.
■ Неаллельное взаимодействие генов приводит к модификации менделевского расщепления по фенотипу 9 : 3 : 1, т.е. к появлению в потомстве гетерозиготы иных расщеплений, например 9 : 3 : 4; 9 : 6 : 1; 12 : 3 : 1 и др.
❖ Основные типы межаллельных взаимодействий:
■ комплементарность;
■ эпистаз;
■ полимерия.
Замечание: комплементарное и эпистатическое взаимодействия возникают в тех случаях, когда признак контролируется одной парой неаллельных генов.
Комплементарное, или дополнительное, взаимодействие — такой тип межаллельного взаимодействия генов, при котором одновременное присутствие в генотипе гибрида доминантных генов разных аллельных пар приводит к появлению нового признака, отсутствующего у обоих родителей.
Пример: наследование окраски цветков душистого горошка (родительские растения с генотипами А-вв, ааВ- имеют белые цветки, гибриды с генотипом А-В- пурпурные; см. таблицу).

Объяснение: пурпурный пигмент образуется с помощью специального фермента, который синтезируется только при наличии обоих доминантных генов: как А, так и В. Цветки родительских особей имеют белый цвет, так как в генотипе каждого из них присутствует только один из этих генов.
Во втором поколении при самоопылении, обеспечивающем равновероятное (случайное) образование гамет и зигот разного типа, наблюдается расщепление по фенотипу в соотношении пурпурной и белой окраски цветков как 9 : 7 (9 пурпурных: A-В- и 7 белых: ЗА-Bb, ЗааВ-. 1aabb).
Эпистаз — такой тип межаллельного взаимодействия генов, при котором аллели одного гена подавляют проявление аллельной пары другого гена, и подавляемый признак не проявляется.
Супрессор (или ген-ингибитор) — ген, подавляющий действие других неаллельных генов. Супрессором может быть как доминантный, так и рецессивный ген.
Доминантный эпистаз — эпистаз, в котором супрессором является доминантный ген. При доминантном эпистазе во втором поколении наблюдается расщепление фенотипа 12:3:1 или 13:3.
Рецессивный эпистаз (криптомерия) — эпистаз, в котором супрессором является рецессивный ген. При рецессивном эпистазе во втором поколении наблюдается расщепление по фенотипу 9:3:4.
Пример эпистаза: наследование окраски шерсти у домашних кроликов. Синтез черного пигмента детерминирует рецессивный ген с, доминантная аллель I другого гена является супрессором, подавляя действие гена с. Тогда кролики с генотипами C-I-, ccl-будут белыми, кролики с генотипами C-ii — серыми, а с генотипом ссii — черными.
Многие признаки контролируются двумя и более парами неаллельных генов (называемых в этом случае полимерными).
Полимерия — взаимодействие нескольких неаллельных полимерных генов. При полимерии степень выраженности фенотипического признака часто зависит от числа полимерных генов, ответственных за его проявление. При кумулятивной полимерии действие генов суммируется; примеры: масса тела, молочность крупного рогатого скота, яйценоскость кур, некоторые параметры умственных способностей человека и др. При некумулятивной полимерии степень проявления признака от числа доминантных генов в генотипе не зависит (пример: оперенность ног у кур).
Плейотропия — зависимость нескольких признаков от одного гена. Каждый плейотропный ген оказывает какое-то основное действие, но модифицирует проявление других генов.
Сцепление генов. Опыты Моргана
Сцепленные гены — любые гены, расположенные в одной хромосоме.
Группа сцепления — все гены, расположенные в одной хромосоме.
■ Количество групп сцепления равно числу пар хромосом (т.е. гаплоидному числу хромосом). У человека 46 хромосом, т.е. 23 группы сцепления.
■ Наследование признаков, за которые отвечают гены из одной группы сцепления, не подчиняется законам Менделя.
Опыты Т. Моргана (1911-1912 гг.): анализ наследования двух пар альтернативных признаков у мух-дрозофил — серого (В) и черного (b) цвета тела и нормальной (V) или укороченной (v) длины крыльев.
Первая серия опытов: скрещивание гомозиготной доминантной (BBVV) особи (с серым цветом тела и нормальной длиной крыльев) с гомозиготной рецессивной (bbvv) особью черного цвета с короткими крыльями. Все потомки F1, в соответствии с первым законом Менделя, являются доминантными гетерозиготными (BbVv) особями серого цвета с нормальными крыльями.
Вторая серия опытов: анализирующее скрещивание гибридов первого поколения — гомозиготной рецессивной (черной короткокрылой) самки (bbvv) с дигетерозиготным (серым с нормальными крыльями) самцом (BbVv). Если предположить, что два гена, относящиеся к разным аллельным парам, локализованы в разных хромосомах, то у дигетерозиготы следует ожидать образования (в равных количествах) четырех типов гамет: BV, bV, Bv и bv. Тогда, согласно третьему закону Менделя, в потомстве должны присутствовать четыре разных фенотипа в равном количестве (по 25%). В действительности присутствовали только два фенотипа (в соотношении 1 : 1).
■ Это означает, что доминантные гены В и V, относящиеся к разным аллельным парам, локализованы в одной хромосоме (из пары гомологичных хромосом) и попадают в одну гамету, а оба рецессивных гена в и v локализованы в другой хромосоме и вместе попадают в другую гамету. Поэтому у дигетерозиготного самца мухи-дрозофилы образуется не четыре типа гамет (когда гены расположены в разных хромосомах), а только два: BV (50%) и bv (50%), и, следовательно, потомки F2 будут иметь два сочетания признаков.
Третья серия опытов: проверка предположения о полном сцеплении генов путем анализирующего скрещивания дигетерозиготной (серой с нормальными крыльями) самки (BbVv) из поколения F1 с гомозиготным рецессивным (черным короткокрылым) самцом (bbvv) из родительского поколения. В результате были получены потомки четырех фенотипов в следующем соотношении: по 41,5% особей с серым телом и нормальными крыльями (генотип BbVv) и особей с черным телом и короткими крыльями (генотип bbvv), и по 8,5% серых короткокрылых особей (генотип Bbvv) и черных особей с нормальными крыльями (генотип bbVv).
Отсюда следует, что сцепленные гены, т.е. гены, локализованные в одной хромосоме, не всегда передаются вместе, т.е. сцепление может быть неполным. Это связано с явлением кроссинговера. вероятность которого в данном случае составляет 17%.
Кроссинговер
Кроссинговер — явление обмена участками гомологичных хрома-тид в процессе их конъюгации в профазе мейоза I.
■ У гетерозиготных организмов кроссинговер приводит к пере-комбинации генетического материала.
■ Кроссинговер происходит не всегда; его частота зависит от расстояния между генами (подробнее о расстоянии между генами см. ниже).
■ Кроссинговер имеет место у всех растений и животных, за исключением самца мухи-дрозофилы и самки тутового шелкопряда.
■ Значение кроссинговера: он позволяет создавать новые комбинации генов и тем самым повысить наследственную изменчивость, необходимую для расширения возможностей адаптации организмов к изменившимся условиям среды.
Рекомбинация — появление новых сочетаний генов в результате кроссинговера, свободного комбинирования хромосом при образовании гамет или при их слиянии в ходе оплодотворения.
Кроссоверные (или рекомбинантные) особи — особи, образующиеся из гамет с новым, полученным путем кроссинговера, сочетанием аллелей.
Закон Моргана (закон сцепленного наследования): гены, расположенные в одной хромосоме (т.е. входящие в одну группу сцепления), наследуются преимущественно, т.е. с наибольшей вероятностью, вместе (сцепленно).
Хромосомная теория наследственности
Хромосомная теории наследственности экспериментально обоснована Т. Морганом с сотрудниками в 1911-1926 гг.
♦ Основные положения хромосомной теории наследственности:
■ гены, ответственные за хранение и передачу наследственной информации, локализованы в определенных участках (локусах) хромосом; различные хромосомы имеют разное число генов;
■ аллельные гены занимают одинаковые локусы в гомологичных хромосомах;
■ в хромосомах гены располагаются в линейной последовательности (друг за другом) и не перекрываются;
■ гены гомологичных хромосом образуют группу сцепления и наследуются преимущественно вместе; число групп сцепления равно гаплоидному набору хромосом (т.е. числу пар гомологичных хромосом);
■ между гомологичными хромосомами возможен обмен аллельными генами (кроссинговер);
■ вероятность кроссинговера пропорциональна расстоянию между генами в хромосомах.
Генетические карты
Генетическая карта хромосом — схема, на которой отображается взаимное расположение генов, находящихся в одной группе сцепления, с учетом их порядка следования и относительных расстояний между ними.
■ Возможность картирования хромосом основана на постоянстве процента кроссинговера между определенными генами.
Расстояние между генами выражается в морганидах.
■ Одна морганида — такое расстояние между генами, при котором вероятность кроссинговера равна 1 %.
♦ Значение генетических карт:
■ в селекции они позволяют прогнозировать возможность получения организмов с определенными сочетаниями признаков;
■ являются основой для генной инженерии;
■ в медицине используются для диагностики ряда тяжелых наследственных заболеваний человека;
■ сравнение генетических карт разных видов живых организмов помогает установить особенности эволюционного процесса.
Цитоплазматическая наследственность
Цитоплазматическая наследственность — наследственность, связанная с действием генов, находящихся в органоидах цитоплазмы, содержащих ДНК (митохондриях и пластидах).
■ Такие гены способны к автономной репликации и равномерному распределению между дочерними клетками.
■ В передаче признаков цитоплазматическая наследственность имеет второстепенное значение.
■ Цитоплазматическое наследование осуществляется только через материнский организм (в мужских половых клетках цитоплазмы мало и в ней митохондрии и пластиды отсутствуют). Примеры: с мутациями генов, локализованных в митохондриях, связано наследование нарушений в действии дыхательных ферментов у дрожжей; с мутациями генов, локализованных в пластидах, связано, в частности, наследование пестролистности у ряда растений (ночной красавицы, львиного зева и др.).
Читайте также: