
Виды и режимы работы мышц реферат
Обновлено: 02.07.2024
Виды работы мышц
Существует два вида работы мышц: статическая и динамическая. Если при работе мышц происходят движения в суставах, то ее называют динамической. Если суставы неподвижны, то работа мышц заключается в поддержании тела в определенном положении. В таком случае мышечную работу называют статической.
Легко понять, чем отличается динамическая и статическая работа мышц: в первом случае суставы совершают движение, во втором случае – остаются неподвижно. Теперь разберемся, чем характеризуется каждый вид работы.
Ключевой характеристикой динамичной работы мышц является энергозатратность. Несмотря на то, что мышцы тратят энергию в статике, во время активного движения ее затрачивается в разы больше.
Динамическая работа мышц
Под динамической работой мышц подразумевается двигательная активность, при которой происходит попеременное расслабление и сокращение мышц для перемещения тела в пространстве или выполнения определенного движения.
При выполнении динамической работы происходят физиологические реакции организма, которых не возникает во время статической мышечной работы. Примером таких реакций служит увеличение пульса и артериального давления во время активности. Интенсивность проявления реакций зависит от разных факторов: тренированности человека, силы и частоты мышечного сокращения, и даже от того, в каком положении находилось тело до начала активности.
Динамическую работу классифицируют по количеству работающих мышц:
- Глобальная – если в движении принимают участие более двух третей от всех мышц тела;
- Региональная – если в движении задействовано менее двух третей от общего количества мышц;
- Локальная – если в движение участвует менее трети от всех мышц.
Например, базовые упражнения, вроде приседаний, становой тяги, прыжков задействуют огромное количество мышц, в результате чего происходит глобальная или региональная динамическая работа. Изолированные упражнения, например, подъем штанги на бицепс, разгибания на трицепс подключают в работу не слишком много мышц, а потому происходит локальная динамическая работа.
Динамическая работа мышц может быть преодолевающей и уступающей, что значит преодоление сопротивления и непротиводействие. Рассмотрим на примере мышц рук: при отведении выполняется преодолевающая динамическая работа, при приведении – уступающая. А при удержании руки в определенном положении выполняется статическая или удерживающая работа мышц.
Статическая работа мышц
Если вам интересно, какая работа мышц называется динамической и статической, то с первой уже разобрались. Динамическая работа возникает во время любого движения или физической активности. Теперь узнаем, какое отличие у статической работы.
При статической работе мышцы постоянно сокращаются, чтобы удерживать тело в определенном положении или обеспечивать выполнение простых бытовых действий.
При статической работе не происходит чрезмерного потребления кислорода и активации кровотока, но проявляются различные физиологические реакции и происходят энергетические затраты. Например, при выполнении статических упражнений, планки или стульчика тело тратит энергию на удержание определенного положения. Поэтому нагрузку мышц можно получить в статике, хотя энергозатраты, конечно, не сравнятся с динамической работой. Физиологические реакции организма в виде учащения пульса и повышения давления зависит от продолжительности работы и силы сокращений.
Между статической и динамической работой мышц есть различия, например, динамическая работа обеспечивается сокращающимися и расслабляющимися мышцами, а статическая – непрерывно сокращающимися. Но эти виды работы последовательно сменяют друг друга в нашей повседневной деятельности и не могут существовать друг без друга.
Что еще важно знать
В реальной жизни мышцы не работают изолированно, поэтому таблиц о конкретно динамической или статической работе вы не найдете. Важно помнить, что в статике всегда есть элементы динамики и наоборот.
Планируя тренировки на увеличение силы мышц, следует включать плиометрические и статические упражнения чтобы развивать медленные и быстрые мышечные волокна, что сделает вас сильнее и выносливее.
Многих интересует быстрое утомление мышц при статической нагрузке. Дело в том, что перманентное сокращение определенных мышц затрудняет насыщение клеток кислородом и выведение продуктов распада, что приводит к усталости. Во время динамических движений такого не происходит, поэтому поднимать и опускать руку вы можете дольше, чем удерживать ее в неподвижном положении.

Мышечное волокно является многоядерной структурой, окруженной мембраной и содержащей специализированный сократительный аппарат — миофибриллы. Кроме этого, важнейшими компонентами мышечного волокна являются митохондрии, системы продольных трубочек — саркоплазматическая сеть (ретикулум) и система поперечных трубочек — Т-система. Функциональной единицей сократительного аппарата мышечной клетки является саркомер (рис. 2.20,А); из саркомеров состоит миофибрилла. Саркомеры отделяются друг от друга Z-пластинками. Саркомеры в миофибрилле расположены последовательно, поэтому сокращение саркомеров вызывает сокращение миофибриллы и общее укорочение мышечного волокна.
Изучение структуры мышечных волокон в световом микроскопе позволило выявить их поперечную исчерченность. Электронно-микроскопические исследования показали, что поперечная исчерченность обусловлена особой организацией сократительных белков миофибрилл — актина (молекулярная масса 42 000) и миозина (молекулярная масса около 500 000). Актиновые филаменты представлены двойной нитью, закрученной в двойную спираль с шагом около 36,5 нм. Эти филаменты длиной 1 мкм и диаметром 6—8 нм, количество которых достигает около 2000, одним концом прикреплены к Z-пластинке. В продольных бороздках актиновой спирали располагаются нитевидные молекулы белка тропомиозина. С шагом, равным 40 нм, к молекуле тропомиозина прикреплена молекула другого белка — тропонина. Тропонин и тропомиозин играют важную роль в механизмах взаимодействия актина и миозина. В середине саркомера между нитями актина располагаются толстые нити миозина длиной около 1,6 мкм. В поляризационном микроскопе эта область видна в виде полоски темного цвета (вследствие двойного лучепреломления) — анизотропный А-диск. В центре его видна более светлая полоска Н. В ней в состоянии покоя нет актиновых нитей. По обе стороны А-диска видны светлые изотропные полоски — I-диски, образованные нитями актина. В состоянии покоя нити актина и миозина незначительно перекрывают друг друга таким образом, что общая длина саркомера составляет около 2,5 мкм. При электронной микроскопии в центре Н-полоски обнаружена М-линия — структура, которая удерживает нити миозина. На поперечном срезе мышечного волокна можно увидеть гексагональную организацию миофиламента: каждая нить миозина окружена шестью нитями актина (рис. 2.20, Б).
При электронной микроскопии видно, что на боковых сторонах миозиновой нити обнаруживаются выступы, получившие название поперечных мостиков. Они ориентированы по отношению к оси миозиновой нити под углом 120°. Согласно современным представлениям, поперечный мостик состоит из головки и шейки. Головка приобретает выраженную АТФазную активность при связывании с актином. Шейка обладает эластическими свойствами и представляет собой шарнирное соединение, поэтому головка поперечного мостика может поворачиваться вокруг своей оси.
Использование микроэлектродной техники в сочетании с интерференционной микроскопией позволило установить, что нанесение электрического раздражения на область Z-пластинки приводит к сокращению саркомера, при этом размер зоны диска А не изменяется, а величина полосок Н и I уменьшается. Эти наблюдения свидетельствовали о том, что длина миозиновых нитей не изменяется. Аналогичные результаты были получены при растяжении мышцы — собственная длина актиновых и миозиновых нитей не изменялась. В результате этих экспериментов выяснилось, что изменялась область взаимного перекрытия актиновых и миозиновых нитей. Эти факты позволили Н. Huxley и A. Huxley предложить независимо друг от друга теорию скольжения нитей для объяснения механизма мышечного сокращения. Согласно этой теории, при сокращении происходит уменьшение размера саркомера вследствие активного перемещения тонких актиновых нитей относительно толстых миозиновых. В настоящее время выяснены многие детали этого механизма и теория получила экспериментальное подтверждение.
В настоящее время принято считать, что биохимический цикл мышечного сокращения состоит из 5 стадий (рис. 3):
3) это взаимодействие обеспечивает высвобождение АДФ и Н3РО4 из актин-миозинового комплекса. Актомиозиновая связь имеет наименьшую энергию при величине угла 45°, поэтому изменяется угол миозина с осью фибриллы с 90° на 45° (примерно) и происходит продвижение актина (на 10–15 нм) в направлении центра саркомера (см. рис. 3, в);
4) новая молекула АТФ связывается с комплексом миозин–F-актин (см. рис. 3, г);

В естественных условиях в организме одиночного мышечного сокращения не наблюдается, так как по двигательным нервам, иннервирующим мышцу, идут серии потенциалов действия. В зависимости от частоты приходящих к мышце нервных импульсов мышца может сокращаться в одном из трех режимов (рис. 2, Б).
• Одиночные мышечные сокращения возникают при низкой частоте электрических импульсов. Если очередной импульс приходит в мышцу после завершения фазы расслабления, возникает серия последовательных одиночных сокращений.
• При более высокой частоте импульсов очередной импульс может совпасть с фазой расслабления предыдущего цикла сокращения. Амплитуда сокращений будет суммироваться, возникнет зубчатый тетанус – длительное сокращение, прерываемое периодами неполного расслабления мышцы.
• При дальнейшем увеличении частоты импульсов каждый следующий импульс будет действовать на мышцу во время фазы укорочения, в результате чего возникнет гладкий тетанус – длительное сокращение, не прерываемое периодами расслабления.
Фазы мышечного сокращения
При раздражении скелетной мышцы одиночным импульсом электрического тока сверхпороговой силы возникает одиночное мышечное сокращение, в котором различают 3 фазы (рис. 2, А):
• латентный (скрытый) период сокращения (около 10 мс), во время которого развивается потенциал действия и протекают процессы электромеханического сопряжения; возбудимость мышцы во время одиночного сокращения изменяется в соответствии с фазами потенциала действия;
• фаза укорочения (около 50 мс);
• фаза расслабления (около 50 мс).
Оптимум и пессимум частоты
Амплитуда тетанического сокращения зависит от частоты импульсов, раздражающих мышцу. Оптимумом частоты называют такую частоту раздражающих импульсов, при которой каждый последующий импульс совпадает с фазой повышенной возбудимости (рис. 2 A) и соответственно вызывает тетанус наибольшей амплитуды. Пессимумом частоты называют более высокую частоту раздражения, при которой каждый последующий импульс тока попадает в фазу рефрактерности (рис. 2, A), в результате чего амплитуда тетануса значительно уменьшается.
Поскольку основной задачей скелетной мускулатуры является совершение мышечной работы, в экспериментальной и клинической физиологии оценивают величину работы, которую совершает мышца, и мощность, развиваемую ею при работе.
Согласно законам физики, работа есть энергия, затрачиваемая на перемещение тела с определенной силой на определенное расстояние: А = FS. Если сокращение мышцы совершается без нагрузки (в изотоническом режиме), то механическая работа равна нулю. Если при максимальной нагрузке не происходит укорочения мышцы (изометрический режим), то работа также равна нулю. В этом случае химическая энергия полностью переходит в тепловую.
Согласно закону средних нагрузок, мышца может совершать максимальную работу при нагрузках средней величины.
При сокращении скелетной мускулатуры в естественных условиях преимущественно в режиме изометрического сокращения, например при фиксированной позе, говорят о статической работе, при совершении движений — о динамической.
Сила сокращения и работа, совершаемая мышцей в единицу времени (мощность), не остаются постоянными при статической и динамической работе. В результате продолжительной деятельности работоспособность скелетной мускулатуры понижается. Это явление называется утомлением. При этом снижается сила сокращений, увеличиваются латентный период сокращения и период расслабления.
В реальных условиях необходимо учитывать состояние ЦНС — снижение силы сокращений сопровождается уменьшением частоты импульсации нейронов, обусловленное как их прямым угнетением, так и механизмами центрального торможения. Еще в 1903 г. И. М. Сеченов показал, что восстановление работоспособности утомленных мышц одной руки значительно ускоряется при совершении работы другой рукой в период отдыха первой. В отличие от простого отдыха такой отдых называют активным.
Работоспособность скелетной мускулатуры и скорость развития утомления зависят от уровня умственной деятельности: высокий уровень умственного напряжения уменьшает мышечную выносливость.
В динамическом режиме работоспособность мышцы определяется скоростью расщепления и ресинтеза АТФ. При этом скорость расщепления АТФ может увеличиваться в 100 раз и более. Ресинтез АТФ может обеспечиваться за счет окислительного расщепления глюкозы. Действительно, при умеренных нагрузках ресинтез АТФ обеспечивается повышенным потреблением мышцами глюкозы и кислорода. Это сопровождается увеличением кровотока через мышцы примерно в 20 раз, увеличением минутного объема сердца и дыхания в 2—3 раза. У тренированных лиц (например, спортсмена) большую роль в обеспечении повышенной потребности организма в энергии играет повышение активности митохондриальных ферментов.
При максимальной физической нагрузке происходит дополнительное расщепление глюкозы путем анаэробного гликолиза. Во время этих процессов ресинтез АТФ осуществляется в несколько раз быстрее и механическая работа, производимая мышцами также больше, чем при аэробном окислении. Предельное время для такого рода работы составляет около 30 с, после чего возникает накопление молочной кислоты, т. е. метаболический ацидоз, и развивается утомление.
Анаэробный гликолиз имеет место и в начале длительной физической работы, пока не увеличится скорость окислительного фосфорилирования таким образом, чтобы ресинтез АТФ вновь сравнялся с его распадом. После метаболической перестройки спортсмен обретает как бы второе дыхание. Подробные схемы метаболических процессов приведены в руководствах по биохимии.
Согласно первому закону термодинамики, общая энергия системы и ее окружения должна оставаться постоянной.
Скелетная мышца превращает химическую энергию в механическую работу с выделением тепла. А. Хиллом было установлено, что все теплообразование можно разделить на несколько компонентов:
1. Теплота активации — быстрое выделение тепла на ранних этапах мышечного сокращения, когда отсутствуют видимые признаки укорочения или развития напряжения. Теплообразование на этой стадии обусловлено выходом ионов Са2+ из триад и соединением их с тропонином.
2. Теплота укорочения — выделение тепла при совершении работы, если речь идет не об изометрическом режиме. При этом, чем больше совершается механической работы, тем больше выделяется тепла.
3. Теплота расслабления — выделение тепла упругими элементами мышцы при расслаблении. При этом выделение тепла не связано непосредственно с процессами метаболизма.
Как отмечалось ранее, нагрузка определяет скорость укорочения. Оказалось, что при большой скорости укорочения количество выделяющегося тепла мало, а при малой скорости велико, так как количество выделяющегося тепла пропорционально нагрузке (закон Хилла для изотонического режима сокращения).
При совершении работы развиваемое мышцей усилие передается на внешний объект с помощью сухожилий, прикрепленных к костям скелета. В любом случае нагрузка преодолевается за счет вращения одной части скелета относительно другой вокруг оси вращения.
Передача мышечного сокращения на кости скелета происходит при участии сухожилий, которые обладают высокой эластичностью и растяжимостью. В случае сокращения мышцы происходит растяжение сухожилий и кинетическая энергия, развиваемая мышцей, переходит в потенциальную энергию растянутого сухожилия. Эта энергия используется при таких формах движения как ходьба, бег, т. е. когда происходит отрыв пятки от поверхности земли.
Скорость и сила, с которой одна часть тела перемещается относительно другой, зависят от длины рычага, т. е. взаимного расположения точек прикрепления мышц и оси вращения, а также от длины, силы мышцы и величины нагрузки. В зависимости от функции, которую выполняет конкретная мышца, возможно превалирование скоростных или силовых качеств. Как уже указывалось в разделе 2.4.1.4, чем длиннее мышца, тем выше скорость ее укорочения. При этом большую роль играет параллельное расположение мышечных волокон относительно друг друга. В этом случае физиологическое поперечное сечение соответствует геометрическому (рис. 2.27, А). Примером такой мышцы может служить портняжная мышца. Напротив, силовые характеристики выше у мышц с так называемым перистым расположением мышечных волокон. При таком расположении мышечных волокон физиологическое поперечное сечение больше геометрического поперечного сечения (рис. 2.27, Б). Примером такой мышцы у человека может служить икроножная мышца.
У мышц веретенообразной формы, например у двуглавой мышцы плеча, геометрическое сечение совпадает с физиологическим только в средней части, в других областях физиологическое сечение больше геометрического, поэтому мышцы этого типа по своим характеристикам занимают промежуточное место
При определении абсолютной силы различных мышц максимальное усилие, которое развивает мышца, делят на физиологическое поперечное сечение. Абсолютная сила икроножной мышцы человека составляет 5,9 кг/см , двуглавой мышцы плеча — 11,4 кг/см2 .
Оценка функционального состояния мышечной системы у человека
При оценке функционального состояния мышечной системы у человека используют различные методы.
Эргометрические методы. Эти методы используют для определения физической работоспособности. Человек совершает работу в определенных условиях и одновременно регистрируются величины выполняемой работы и различные физиологические параметры: частота дыхания, пульс, артериальное давление, объем циркулирующей крови, величина регионарного кровотока, потребляемого О2, выдыхаемого СО2 и т.д. С помощью специальных устройств — велоэргометров или тредбанов (бегущая дорожка) — создается возможность дозировать нагрузку на организм человека.
Эти методы исследования скелетной мускулатуры человека нашли широкое применение в физиологической и клинической практике. В зависимости от задач исследования проводят регистрацию и анализ суммарной электромиограммы (ЭМГ) или потенциалов отдельных мышечных волокон. При регистрации суммарной ЭМГ чаще используют накожные электроды, при регистрации потенциалов отдельных мышечных волокон — многоканальные игольчатые электроды.
Преимуществом суммарной электромиографии произвольного усилия является неинвазивность исследования и, как правило, отсутствие электростимуляции мышц и нервов. На рис. 2.28 приведена ЭМГ мышцы в покое и при произвольном усилии. Количественный анализ ЭМГ заключается в определении частот волн ЭМГ, проведении спектрального анализа, оценки средней, амплитуды волн ЭМГ. Одним из распространенных методов анализа ЭМГ является ее интегрирование, поскольку известно, что величина интегрированной ЭМГ пропорциональна величине развиваемого мышечного усилия.
Используя игольчатые электроды, можно регистрировать как суммарную ЭМГ, так и электрическую активность отдельных мышечных волокон. Регистрируемая при этом электрическая активность в большей степени определяется расстоянием между отводящим электродом и мышечным волокном. Разработаны критерии оценки параметров отдельных потенциалов здорового и больного человека. На рис. 2.29 приведена запись потенциала двигательной единицы человека.
• изотонически – мышца укорачивается при постоянном напряжении (внешней нагрузке); изотоническое сокращение воспроизводится только в эксперименте;
• изометричеки – напряжение мышцы возрастает, а ее длина не изменяется; мышца сокращается изометрически при совершении статической работы; ![]()

• ауксотонически – напряжение мышцы изменяется по мере ее укорочения; ауксотоническое сокращение выполняется при динамической преодолевающей работе.
Саркоплазматический ретикулум
Саркоплазматический ретикулум [СР (SR)] -разветвленная подобная эндоплазматическому ретикулуму органелла, окружающая индивидуальные миофибриллы подобно сетке (в верхней части схемы в качестве примера приведен СР клетки сердечной мышцы). В покоящихся клетках концентрация Са2+ очень низка (менее 10-5 М). Однако в саркоплазматическом ретикулуме уровень ионов Са2+ существенно выше (около 10-3 М). Высокая концентрация Са2+ в СР поддерживается Са2+-АТФ-азами. Кроме того, в СР имеется специальный белок кальсеквестрин (55 кДа), который благодаря высокому содержанию кислых аминокислот способен прочно связывать ионы Са2+.
Переносу потенциала действия на СР индивидуальной миофибриллы способствуют поперечные трубочки Т-системы, представляющие трубчатые впячивания клеточной мембраны и находящиеся в тесном контакте с индивидуальными миофибриллами. Деполяризация плазматической мембраны передается через Т-трубочки на потенциал-управляемый мембранный белок (так называемый "SR-foot") прилегающей мембраны СР, который открывает Са2+-каналы. Результатом является выброс ионов Са2+ из СР в пространство между филаментами актина и миозина до уровня ≥10-5 M. В конечном итоге выброс ионов Са2+ является пусковым механизмом процесса сокращения миофибрилл.
В. Регуляция ионами кальция
В расслабленной скелетной мышце комплекс тропонина (субъединицы = Т, С, I) с тропомиозином препятствует взаимодействию миозиновых головок с актином.
Быстрое увеличение в цитоплазме концентрации ионов кальция в результате открывания каналов СР приводит к связыванию Са2+ с С-субъединицей тропонина. Последняя по свойствам близка кальмодулину (см. рис. 375). Связывание ионов Са2+ вызывает конформационную перестройку в тропонине, тропонинтропомиозиновый комплекс разрушается и освобождает на молекуле актина участок связывания с миозином (на схеме выделен красным цветом). Это инициирует цикл мышечного сокращения (см. с. 324)
В отсутствие последующего стимулирования АТФ-зависимые кальциевые насосы мембраны СР быстро снижают концентрацию ионов Ca2+ до исходного уровня. Как следствие, комплекс Са2+ с тропонином С диссоциирует, тропонин восстанавливает исходную конформацию, место связывания миозина на актине блокируется и мышца расслабляется.
Таким образом, при сокращении мышечного волокна скелетных мышц позвоночных происходит следующая последовательность событий. При поступлении сигнала от двигательного нейрона мембрана мышечной клетки деполяризуется, сигнал передается на Сa2+-каналы СР. Са2+-каналы открываются, внутриклеточный уровень ионов Са2+ возрастает. Ионы Сa2+ связывается с тропонином С, вызывая конформационную перестройку в тропонине, что влечет за собой разрушение комплекса тропонин-тропомиозин и дает возможность головкам миозина связываться с актином. Происходит инициация актин-миозинового цикла.
По завершении сокращения уровень ионов Са2+ снижается за счет активного обратного транспорта Са2+ в СР, тропонин С отдает Са2+, комплекс тропонин-тропомиозин занимает исходное положение на молекуле актина, блокируя актин-миозиновый цикл. Результатом является расслабление мышцы.
Описаны режимы работы мышц (режимы мышечного сокращения, режимы сокращения мышц): изометрический, преодолевающий (концентрический), уступающий (эксцентрический). Дана характеристика изменений (гипертрофии, силы и повреждений мышц), происходящих в скелетных мышцах при выполнении силовых упражнений в различных режимах их работы.
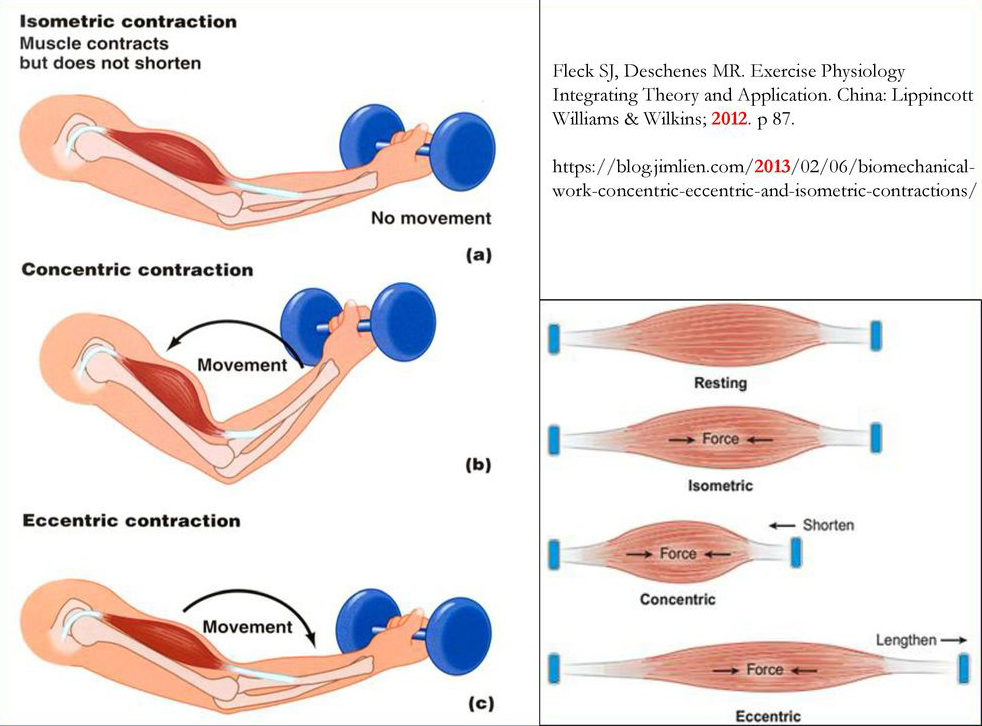
Режимы работы (сокращения) мышц
Классификация режимов работы мышц на основе изменения длины мышцы
В биомеханике основным классификационным признаком является длина мышцы. На основе того, что происходит с длиной мышцы режимы работы мышц делятся на изометрический и динамический.
Изометрический режим работы мышц
Скелетные мышцы могут работать не меняя своей длины. Такой режим работы мышц называется изометрический. Иногда говорят, что мышца работает в статическом режиме. Как пример такой работы — удержание гантели в руке, не меняя ее положения. В этом случае мышцы-сгибатели предплечья (двуглавая мышца плеча, плече-лучевая мышца и др.) не меняют своей длины. В чем особенность этого режима? Мышца возбуждена, должна укорачиваться, а ее длина не меняется. Это происходит из-за того, что внешняя сила уравновешивает силу, которую развивает мышца (правильнее, конечно говорить, что момент внешней силы равен моменту силы тяги мышцы, но этот нюанс можно опустить).
Динамический режим работы мышц
Если длина мышцы меняется, неважно она уменьшается или увеличивается, то принято говорить, что мышца работает в динамическом режиме. Как пример такой работы — сгибание и разгибание руки в локтевом суставе, удерживая в руке гантель. В этом случае мышцы-сгибатели предплечья вначале укорачиваются (это происходит при сгибании руки), затем — удлиняются (это происходит при разгибании руки в локтевом суставе).
Преодолевающий режим работы мышц (концентрический режим работы мышц)
Уступающий режим работы мышц (эксцентрический режим работы мышц)
Различные режимы работы мышц иллюстрируют рис.1 и рис.2.
Рис.1. Режимы работы мышц (классификация) Рис.2. Режимы работы мышц
Следует обратить внимание на тот факт, что мышцы-антагонисты при выполнении движения работают в различных режимах. Например, при сгибании руки мышцы-сгибатели укорачиваются (преодолевающий режим), а мышцы-разгибатели (их антагонисты) — удлиняются (уступающий режим).
Изменения, происходящие в мышцах непосредственно или сразу после тренировочного занятия (срочный эффект тренировки)
Многочисленными исследованиями доказано, что выполнение физических упражнений в эксцентрическом (уступающем режиме, когда мышца удлиняется) режиме вызывает большие структурные повреждения мышечных волокон, чем другие режимы сокращения мышцы. Эти повреждения затрагивают в первую очередь Z-диски саркомеров, а также белки цитоскелета.
С биохимической точки зрения эксцентрические упражнения (упражнения, выполняемые в эксцентрическом режиме) представляют для организма значительно больший стресс, чем упражнения, производимые в других режимах: уровень креатинкиназы в крови (фермента, содержащегося в мышечных волокнах и выделяющегося в кровь при их разрушении) при работе в эксцентрическом режиме значительно превышает соответствующий показатель при работе в концентрическом (преодолевающем) и изометрическом режимах.
Если измерить силу мышц после выполнения упражнений в эксцентрическом режиме, то окажется, что она уменьшается значительно больше, чем при выполнении упражнений в концентрическом режиме. О чем это говорит? Это говорит о том, что в эксцентрическом режиме повреждено больше мышечных волокон.
Изменения, происходящие в мышцах после длительного применения физических упражнений (кумулятивный тренировочный эффект)
Показано, что долговременная адаптация скелетных мышц к упражнениям, выполняемым в эксцентрическом режиме, проявляется в несколько большей гипертрофии скелетных мышц по сравнению с другими режимами. Силовые тренировки в эксцентрическом режиме приводят к увеличению силы и жесткости скелетных мышц.
При выполнении силовых упражнений в изометрическом режиме увеличивается степень перекрытия мышечных и сухожильных волокон, несколько утолщается сухожилие и увеличивается площадь прикрепления сухожилия к кости. Именно поэтому рекомендуется в конце тренировки выполнять упражнения в изометрическом режиме (около 15 минут). Считается, что это позволяет уменьшить количество травм опорно-двигательного аппарата человека.
Если мышца сокращается в динамическом режиме (концентрическом или эксцентрическом режимах), в ней через некоторое время увеличивается длина мышечных волокон и уменьшается длина сухожилия. Компьютерное моделирование (U. Proske, D.L. Morgan, 2001) подтвердило целесообразность удлинения мышечной части и укорочения сухожильной. Авторами показано, что долговременная адаптация к выполнению эксцентрических упражнений проявляется в увеличении количества саркомеров в миофибриллах мышечных волокон и уменьшении сухожильной части. Это приводит к изменению оптимальной длины мышцы при развитии активного напряжения.
При выполнении силовых упражнений в динамическом режиме (концентрическом или эксцентрическом) возрастает количество нервных волокон, иннервирующих скелетную мышцу (в 4-5 раз больше, чем в изометрическом режиме).
Классификация режимов работы мышц на основе изменения длины и (или) тонуса мышцы
В физиологии принята несколько иная классификация режимов работы скелетных мышц. В качестве классификационных признаков используется длина и тонус мышцы. Согласно этим признакам режимы работы мышц делятся на три вида: изотонический, изометрический, ауксотонический. Эту классификацию даю по учебнику А.С. Солодкова, Е.Б.Сологуб (2005)
Изотонический режим работы мышцы
Изотонический режим (режим постоянного тонуса мышцы) наблюдается при отсутствии нагрузки на мышцу, когда мышца закреплена с одного конца и свободно сокращается. Напряжение в ней при этом не изменяется. Это происходит при раздражении изолированной мышцы лягушки, закрепленной одним концом на штативе. В таком режиме в организме человека работает только одна мышца — мышца языка. В настоящее время в литературе в качестве изотонического рассматривается такой режим работы мышцы с нагрузкой, при котором по мере изменения длины мышцы ее тонус не меняется.
Изометрический режим работы мышц
Изометрический режим (режим постоянной длины мышцы) характеризуется напряжением мышцы в условиях, когда она закреплена с обеих концов или когда она не может поднять слишком большой груз. В этом случае в мышечном волокне (миофибрилле) происходят процессы сокращения, при этом одни саркомеры укорачиваются, а другие — удлиняются.
Ауксотонический режим работы мышц
Ауксотонический режим (смешанный режим) характеризуется изменением и длины и тонуса мышцы. При этом режиме сокращения происходит перемещение груза. Этот режим также называется динамическим. Имеются две разновидности этого режима: преодолевающий (концентрический) — длина мышцы уменьшается, уступающий (эксцентрический) — длина мышцы увеличивается.
Классификация режимов работы мышц на основе изменения скорости сокращения мышцы
Изокинетический режим работы мышц
Литература
- Самсонова, А.В. Моторные и сенсорные компоненты биомеханической структуры физических упражнений /А.В. Самсонова: автореф. дис…докт. пед. наук.- СПб.- 1998.- 48 с.
- Самсонова А.В, Барникова И.Э., Азанчевский В.В. Влияние силовых тренировок, выполняемых в различных режимах сокращения, на гипертрофию скелетных мышц человека // Труды каф. биомеханики. Сб. статей /Под ред. А.В.Самсоновой. В.Н.Томилова.- СПб, 2010.- С. 115-131.
- Jones, D.A. Human muscle strength training: the effects of tree different regimes and the nature of the resultant changes / D.A.Jones, O.M. Rutherford // J. Physiol. ,1987.- V.391.– P. 1-11.
- Frieden J, Lieber R.L. Eccentric exercise-induced injuries to contractile and cytoskeletal muscle fibre components //Acta Physiol. Scand. 2001, 171, P.321-326.
- Proske U. , Morgan D.L. Muscle damage from eccentric exercise: mechanism, mechanical signs, adaptation and clinical applications //Journal of Physiology, 2001.– V. 537.– N2, P.333–345.
Похожие записи:
Effect of KAATSU-training on the maximum voluntary isometric contraction of lower extremity muscles of qualified football players
Изучалось влияние KAATSU-тренинга на изометрическую силу мышц квалифицированных футболистов. Установлено, что интенсивный рост максимальной силы мышц…
Удаление фасции снижает силу мышц
В опытах на диких индейках показано, что удаление фасции снижает силу мышц на 30%. Эти результаты ставят под…
Искусственный интеллект в спортивной тренировке
Описана система комплексного контроля в спорте, построенная на основе искусственного интеллекта. Разработанная система внедрена в подготовку пауэрлифтеров высокой…
Тест времени реакции на сигнал
Представлена программа расчета времени реакции на сигнал, предназначенная для использования в учебных целях, например на занятиях по…
Саркоплазматическая гипертрофия мышц
Дано определение и описаны механизмы саркоплазматической гипертрофии скелетных мышц. Показано, что этот вид гипертрофии мышц широко…
Классификация типов конституции человека М.В. Черноруцкого
Рассмотрена классификация типов конституции человека, разработанная выдающимся терапевтом М.В. Черноруцким в 1925 году. Классификация типов конституции человека М.В.
Типы гипертрофии скелетных мышц человека
В статье дается классификация различных видов гипертрофии скелетных мышц человека на основе ряда классификационных признаков: времени проявления…
Миомейкер: Мембранный активатор слияния миобластов и образования мышц
Ученые установили, что для образования мышечных волокон необходимо слияние клеток-предшественников, которые называются миобластами. Эти клетки имеют только…
Отсроченное начало болезненности мышц. Стратегии лечения и факторы эффективности
Описаны симптомы, причины, теории отсроченного начала болезненности мышц (запаздывающих болезненных ощущений, DOMS), а также способы уменьшения этих болей:…
Срочные гормональные ответы у элитных тяжелоатлетов-юниоров
Изучалось изменение концентрации в крови: тестостерона, кортизола, гормона роста, бета-эндорфина и лактата у тяжелоатлетов-юниоров…
Гидролиз АТФ в мышечных волокнах
Описано протекание реакции гидролиза АТФ в мышечных волокнах. Показано, что именно реакция гидролиза АТФ приводит к ацидозу,…
Вы можете заказать написание любой учебной работы на любую тему.
* Данная работа не является научным трудом, не является выпускной квалификационной работой и представляет собой результат обработки, структурирования и форматирования собранной информации, предназначенной для использования в качестве источника материала при самостоятельной подготовки учебных работ.
Содержание Физиология мышц классификация мышечных волокон Функции скелетных и гладких мышц Режимы сокращения мышц Виды сокращений Строение мышцы Функциональные особенности гладких мышц Физиология , мышц классификация мышечных волокон Мышечные волокна делят на 3 вида : скелетные, сердечные и гладкие . Скелетные волокна подразделяются на фазные ( они генерируют ПД ) и тонические ( не способны генерировать полноценный потенциал действия распространяющегося типа ). Фазные волокна делятся на быстрые волокна ( белые, гликолитические ) и медленные волокна ( красные, окислительные волокна ). Гладкие мышц ы делятся на тонические и фазно тонические . Тонические волокна не способны развивать " быстрые " сокращения . В свою очередь фазно тонические мышцы можно условно разделить на обладающие автоматией - способные к спонтанной генерации фазных сокращений, и на мышцы, не обладающие свойством автоматии . Основным морфо - функциональным элементом нервно - мышечного аппарата является двигательная единица ( ДЕ ). ДЕ - это мотонейрон с иннервируемыми им мышечными волокнами . Аксон мотонейрона из спинного мозга проходит в составе периферических нервов до мышцы, внутри которой разветвляется на множество концевых веточек . Каждая концевая веточка заканчивается на одном мышечном волокне, образуя нервно-мышечный синапс . Импульсы, идущие по аксону мотонейрона, активируют все иннервируемые им мышечные волокна . Поэтому ДЕ функционирует как единое морфофункциональное образование . Функции скелетных и гладких мышц Скелетные мышцы составляют 40% от массы тела и выполняют ряд важных функций : 1 - передвижение тела в пространстве, 2 - перемещение частей тела относительно друг друга, 3 - поддержание позы, 4 - передвижение крови и лимфы, 5 - выработка тепла, 6 - участие в акте вдоха и выдоха, 7 - двигательная активность как важнейший антиэнтропийный и антистрессовый фактор ( тезисы " движение - это жизнь " или " кто много двигается, тот много живет " - имеют реальную материальную основу ), 8 - депонирование воды и солен, 9 - защита внутренних органов ( например, органов брюшной полости ). Гладкие мышцы обеспечивают функцию полых органов, стенки которых они образуют . В частности, благодаря гладким мышцам осуществляется изгнание содержимого из мочевого пузыря, кишки, желудка, желчного пузыря, матки . Гладкие мышцы обеспечивают сфинктерную функцию - создают условия для хранения содержимого полого органа в этом органе, например, мочу в мочевом пузыре, плод в матке . Важнейшую роль выполняют гладкие мышцы в системе кровообращения и лимфообращения - изменяя просвет сосудов, гладкие мышцы тем самым адаптируют регионарный кровоток к местным потребностям в кислороде, питательных веществах . Гладкие мышцы могут существенно влиять на функцию связочного аппарата, т.к содержатся во многих связках и при своем сокращении меняют состояние данной связочной структуры . Например, ГМК ( гладкомышечные клетки ) содержатся в широкой связке матки . Режимы сокращения мышц Для скелетной мышцы характерны два основных режима сокращения - изометрический и изотонический . Изометрический режим проявляется в том, что в мышце во время ее активности нарастает напряжение ( генерируется сила ), но из-за того, что оба конца мышцы фиксированы ( например, мышца пытается поднять большой груз ) - она не укорачивается . Изотонический режим проявляется в том, что мышца первоначально развивает напряжение ( силу ), способную поднять данный груз, а потом мышца укорачивается - меняет свою длину, сохраняя напряжение, равное весу поднимаемого груза . Так как изотоническое сокращение не является " чисто " изотоническим ( элементы изометрического сокращения имеют место в самом начале сокращения мышцы ), а изометрическое сокращение тоже не является " чисто " изотоническим ( элементы смещения все-таки есть, несомненно ), то предложено употреблять термин " ауксотоническое сокращение " - смешанное по характеру . Понятия " изотонический ", " изометрический " важны для анализа сократительной активности изолированных мышц и для понимания биомеханики сердца . Режимы сокращения гладких мышц . Целесообразно выделить изометрический и изотонический режимы ( и, как промежуточный - ауксотонический ). Например, когда мышечная стенка полого органа начинает сокращаться, а орган содержит жидкость, выход для которой перекрыт сфинктером, то возникает ситуация изометрического режима : давление внутри полого органа растет, а размеры ГМК не меняются ( жидкость не сжимается ). Если это давление станет высоким и приведет к открытию сфинктера, то ГМК переходит в изотонический режим функционирования - происходит изгнание жидкости, т.е. размеры ГМК уменьшаются, а напряжение или сила - сохраняется постоянной и достаточной для изгнания жидкости . Виды сокращений У скелетной мышцы выделяют одиночное сокращение и суммированное сокращение, или тетанус . Одиночное сокращение - это сокращение, которое возникает на одиночный стимул, достаточный для вызова возбуждения мышцы . После короткого скрытого периода ( латентный период ) начинается процесс сокращения . При регистрации сократительной активности в изометрических условиях ( два конца неподвижно закреплены ) в первую фазу происходит нарастание напряжения ( силы ), а во вторую - ее падение до исходной величины . Соответственно эти фазы называют фазой напряжения и фазой расслабления . При регистрации сократительной активности в изотоническом режиме ( например, в условиях обычной миографическо й записи ) эти фазы будут называться соответственно фазой укорочения и фазой удлинения . В среднем сократительный цикл длится около 200 мс ( мышцы лягушки ) или 30 - 80 мс ( у теплокровных ). Если на мышцу действует серия прямых раздражении ( минуя нерв ) или непрямых раздражении ( через нерв ), но с большим интервалом, при котором всякое следующее раздражение попадает в период после окончания 2-й фазы, то мышца будет на каждый из этих раздражителей отвечать одиночным сокращением . Суммированные сокращения возникают в том случае, если на мышцу наносятся 2 и более раздражения, причем всякое последующее раздражение ( после предыдущего ) наносится либо во время 2-й фазы ( расслабления или удлинения ), либо во время 1-й фазы ( укорочения или напряжения ). Одиночное сокращение А - потенциал действия ; Б - сокращение мышцы 1 - фаза напряжения ; 2 - фаза расслабления Суммированное сокращение а - одиночное сокращение ; б-г - зубчатый тетанус ; д - гладкий тетанус В случае, когда всякое второе раздражение попадает в период фазы расслабления ( удлинения ), возникает частичная суммация - сокращение еще полностью не закончилось, а уже возникло новое . Если подается много раздражителей с подобным интервалом, то возникает явление зубчатого тетануса . Если раздражители наносятся с меньшим интервалом и каждое последующее раздражение попадает в фазу укорочения, то возникает так называемый гладкий тетанус . Строение мышцы Скелетная мышца состоит из пучков вытянутых в длину клеток - мышечных волокон, обладающих тремя свойствами : возбудимостью, проводимостью и сократимостью . Отличительной чертой мышечных клеток от клеток, не обладающих свойством сократимости, является наличие саркоплазматического ретикулума . Он представляет собой замкнутую систему внутриклеточных трубочек и цистерн, окружающих каждую миофибриллу . В мембране саркоплазматического ретикулума находятся две транспортные системы, обеспечивающие освобождение от ретикулума ионов кальция при возбуждении и их возврат из миоплазмы обратно в ретикулум при расслаблении мышцы . В механизме освобождения ионов кальция из ретикулума при возбуждении мышечной клетки важную роль играет система поперечных трубочек ( Т-система ), представляющих собой в ы пячивания поверхностной мембраны мышечного волокна . Мышечные волокна имеют диаметр от 10 до 100 мкм и длину от 5 до 400 мм ( в зависимости от длины мышцы ). В каждом мышечном волокне содержится до 1000 и более сократительных элементов миофибрилл, толщиной 1-3 мкм . Каждая миофибрилла состоит из множества параллельно лежащих толстых и тонких нитей - миофиламентов . Толстые нити состоят из молекул белка миозина, а тонкие - из белка актина . Расположение миозиновых и тонких актиновых белковых нитей строга упорядочено (рис.4.1 ). Пучок лежащих в середине саркомера нитей миозина выглядит в световом микроскопе как темная полоска . Благодаря свойству двойного лучепреломления в поляризованном свете ( то есть анизотропии ) она называется А-диском . По обе стороны от А-диска находятся участки, которые содержат только тонкие нити актина и поэтому выглядят светлыми . Эти изотропные J -диски тянутся до Z -пластин . Благодаря такому периодическому чередованию светлых и темных полос миофибриллы скелетной мышцы выглядят исчерченными ( поперечно - полосатыми ). Если мышца расслаблена, то в средней части А-диска различается менее плотная Н-зона, состоящая только из толстых миофиламентов . Н-зона не просматривается во время сокращения мышцы . По середине J -диска проходит темная полоска - это Z линия . Участок миофибриллы между двумя Z линиями называется саркомером . Схема саркомера мышечного волокна и взаимного расположения толстых миозиновых и тонких актиновых миофиламентов . Z - линии, разделяющие два соседних саркомера; J - изотропный диск; А - анизотропный диск; Н - участок с уменьшенной анизотропностью Механизмы сокращения мышечного волокна . В покоящихся мышечных волокнах при отсутствии импульсации мотонейрона поперечные миозиновые мостики не прикреплены к актиновым миофиламентам . При сокращении мышцы длина А-дисков не меняется, J - диски укорачиваются, а Н-зона А-дисков может исчезать (рис.4.3 ). Эти данные явились основой для создания теории, объясняющей сокращение мышцы механизмом скольжения ( теорией скольжения ) тонких актиновых миофиламентов вдоль толстых миозиновых . В результате этого миозиновые миофиламенты втягиваются между окружающими их актиновыми . Это приводит к укорочению каждого саркомера, а значит, и всего мышечного волокна . Молекулярный механизм сокращения мышечного волокна состоит в том, что возникающий в области концевой пластинки потенциал действия распространяется по системе поперечных трубочек вглубь волокна, вызывает деполяризацию мембран цистерн саркоплазматического ретикулума и освобождение из них ионов кальция . Свободные ионы кальция в межфибриллярном пространстве запускают процесс сокращения . Совокупность процессов, обуславливающих распространение потенциала действия вглубь мышечного волокна, выход ионов кальция их саркоплазматического ретикулума, взаимодействие сократительных белков и укорочение мышечного волокна называют " электромеханическим сопряжением ". Временная последовательность между возникновением потенциала действия мышечного волокна, поступлением ионов кальция к миофибриллам и развитием сокращения волокна показана на рисунке . Схема сокращения мышцы А . Поперечные мостики между актином и миозином разомкнуты . Мышца находится в расслабленном состоянии . Б. Замыкание поперечных мостиков между актином и миозином . Совершение головками мостиков гребковых движений по направлению к центру саркомера . Скольжение актиновых нитей вдоль миозиновых, укорочение саркомера, развитие тяги . При возбуждении волокна Са +2 выходит из цистерн саркоплаз матического ретикулума и, следовательно, концентрация его вблизи миофибрилл возрастает . Под влиянием активирующих ионов Са молекула тропонина изменяет свою форму таким образом, что выталкивает тропомиозин в желобок между двумя нитями актина, освобождая тем самым участки для прикрепления миозиновых поперечных мостиков к актину . В результате поперечные мостики прикрепляются к актиновым нитям . Поскольку головки миозина совершают " гребковые " движения в сторону центра саркомера происходит " втягивание " актиновых миофиламентов в пром ежутки между толстыми миозиновы ми нитями и укорочение мышцы . Источником энергии для сокращения мышечных волокон служит АТФ . При однократном движении поперечных мостиков вдоль актиновых нитей ( гребковых движениях ) саркомер укорачивается примерно на 1% его длины . Следовательно, для полного изотонического сокращения мышцы необходимо совершить около 50 таких гребковых движений . Только ритмическое прикрепление и отсоединение головок миозина может втянуть нити актина вдоль миозиновых и совершить требуемое укорочение целой мышцы . Напряжение, развиваемое мышечным волокном, зависит от числа одновременно замкнутых поперечных мостиков . Скорость развития напряжения или укорочения волокна определяется частотой замыкания поперечных мостиков, образуемых в единицу времени, то есть скоростью их прикрепления к актиновым миофиламентам . С увеличением скорости укорочения мышцы число одновременно прикрепленных поперечных мостиков в каждый момент времени уменьшается . Этим и можно объяснить уменьшение силы сокращения мышцы с увеличением скорости ее укорочения . Поскольку возврат ионов кальция в цистерны саркоплазматического ретикулума идет против диффузионного градиента, то этот процесс требует затрат энергии . Ее источником служит АТФ . Одна молекула АТФ затрачивается на возврат 2-х ионов кальция из межфибриллярного пространства в цистерны . Таким образом, кальций в мышечных волокнах играет роль внутриклеточного посредника, связывающего процессы возбуждения и сокращения . Регуляция силы сокращения мышц . Для регуляции величины напряжения мышцы центральная нервная система использует три механизма . 1 . Регуляция числа активных ДЕ . Чем больше число ДЕ мышцы включается в работу, тем большее напряжение она развивает . При необходимости развития небольших усилий и соответственно малой импульсации со стороны центральных нервных структур, регулирующих произвольные движения, в работу включаются, прежде всего, медленные ДЕ, мотонейроны, которые имеют наименьший порог возбуждения . По мере усиления центральной импульсации к работе подключаются быстрые, устойчивые к утомлению ДЕ, мотонейроны которых имеют более высокий порог возбуждения . И наконец, при необходимости увеличения силы сокращения более 20-25 % от максимальной произвольной силы ( МП С ), активируются быстрые, легко утомляемые мышечные волокна, иннервируемые крупными мотонейронами с самым высоким порогом возбуждения . Таким образом, первый механизм увеличения силы сокращения состоит в том, что при необходимости повысить величину напряжения мышцы в работу вовлекается большее количество ДЕ . Последовательность включения разных по морфофункциональным признакам ДЕ определяется интенсивностью центральных возбуждающих влияний и порогом возбудимости спинальных двигательных нейронов . 2 . Регуляция частоты импульсации мотонейронов . При слабых сокращениях скелетных мышц импульсация мотонейронов составляет 5 - 10 имп/с . Для каждой отдельной ДЕ чем выше ( до определенного предела ) частота возбуждающих импульсов, тем больше сила сокращения ее мышечных волокон и тем больше ее вклад в развиваемое всей мышцей усилие . С увеличением частоты раздражения мотонейронов все большее количество ДЕ начинает работать в режиме гладкого тетануса, увеличивая тем самым свою силу по сравнению с одиночными сокращениями в 2-3 раза . В реальных условиях мышечной деятельности человека большая часть ДЕ активируется в диапазоне от 0 до 50% МПС . Лишь около 10% ДЕ вовлекаются с дальнейшим возрастанием силы сокращения . Следовательно, при увеличении силы сокращения более 50% от максимальной - основное значение, а в диапазоне сил от 75 до 100% МПС - даже исключительное, принадлежит росту частоты импульсации двигательных нейронов . 3 . Синхронизация активности различных ДЕ во времени . При сокращении мышцы всегда активируется множество составляющих ее ДЕ . Суммарный механический эффект при этом зависит от того, как связаны во времени импульсы, посылаемые разными мотонейронами к своим мышечным волокнам . При небольших напряжениях большинство ДЕ работают несинхронно . Совпадение во времени импульсов мотонейронов отдельных ДЕ называется синхронизацией . Чем большее количество ДЕ работает синхронно, тем большую силу развивает мышца . Синхронизация активности ДЕ играет важную роль в начале любого сокращения и при необходимости выполнения мощных, быстрых сокращений ( прыжки, метания и т.п.) . Чем больше совпадают периоды сокращения разных ДЕ, тем с большей скоростью нарастает напряжения всей мышцы и тем большей величины достигает амплитуда ее сокращения . Функциональные особенности гладких мышц Гладкие мышцы находятся в стенках внутренних органов и кровеносных сосудов . Регуляция их тонуса и сократительной активности осуществляется эфферентными волокнами симпатической и парасимпатической нервной системы, а также местными гуморальными и физическими воздействиями . Сократительный аппарат гладких мышц, как и скелетных, состоит из толстых миозиновых и тонких актиновых нитей . Вследствие их нерегулярного распределения клетки гладких мышц не имеют характерной для скелетной и с ердечной мышцы поперечной исчер ченности . Гладкомышечные клетки имеют веретенообразную форму, длину 50-400 мкм и толщину 2-10 мкм . Они отделены друг от друга узкими щелями ( 60-150 нм ). Возбуждение электротонически распространяется по мышце от клетки к клетке через особые плотные контакты ( нексусы ) между плазматическими мембранами соседних клеток . Волокна гладких мышц сокращаются в результате относительного скольжения миозиновых и актиновых нитей, но скорость их сокращения и скорость расщепления АТФ в 100-1000 раз меньше, чем в скелетных мышцах . Поэтому гладкие мышцы хорошо приспособлены к длительному тоническому сокращению без развития утомления . Гладкие мышцы, обладающие спонтанной активностью, способны сокращаться и при отсутствии прямых возбуждающих нервных и гуморальных воздействий ( например, ритмические сокращения гладких мышц кишечника ). Спонтанная активность гладкомышечных клеток связана и с их растяжением, вызывающим деполяризацию мембраны мышечного волокна, возникновение серии распространяющихся потенциалов действия, с последующим сокращением клетки . Гладкие мышцы, не обладающие спонтанной активностью сокращаются под влиянием импульсов вегетативной нервной системы . Так, в отличие от мышц кишечника, мышечные клетки артерий, семенных протоков и радужки обладают слабой спонтанной активностью, или вообще не проявляют ее . Отдельные нервные импульсы не способны вызвать пороговую деполяризацию таких клеток и их сокращение . Потенциал действия волокна с последующим сокращением возникает лишь при поступлении к нему серии импульсов с частотой 1 имп/с и выше . В гладких мышцах, не обладающих спонтанной активностью возбуждение также передается от одной клетки к последующим через плотные контакты их мембран ( нексусы ). Подобно скелетной и сердечной мышцам гладкие мышцы расслабляются, если концентрация ионов кальция снижается ниже 10 -8 моль/л . Однако в связи со слабо развитым саркоплазматическим ре тикулумом и медленным переносом ионов кальция через мембрану клетки, расслабление происходит гораздо медленнее, чем в случае поперечно-полосатых волокон скелетных мышц .
Читайте также: