
Строение и движение ресничек и жгутиков реферат
Обновлено: 18.05.2024
Обратимся к другим биологическим механохимическим системам, ответственным за движения клеток и внутриклеточных органоидов. Краткий перечень таких систем приведен на стр. 218.
Множество одноклеточных организмов — бактерий и простейших— перемещается в жидкой среде, совершая, тем самым, работу в результате движения специальных сократительных систем— жгутиков и ресничек. Реснички функционируют и в ряде органов многоклеточных. Так, гребневики регулируют ориентацию своих тел согласованными движениями ресничек; реснички
создают ток жидкости в жабрах двустворчатых моллюсков, в трахеях человека и т. д.
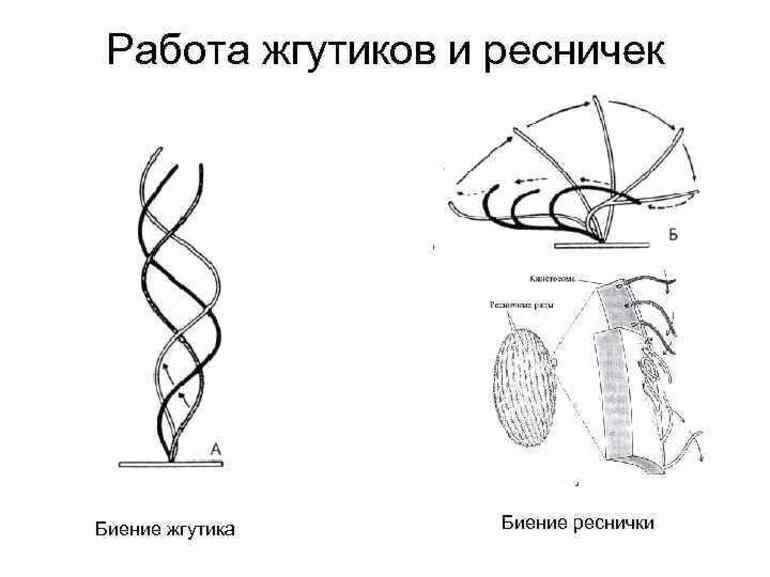
Жгутики выполняют волнообразное движение, распространяющееся вдоль жгутика от его основания к концу. Типы таких движений показаны схематически на рис. 5.28 ([159], см. также [160]).
Клетка может двигаться вперед или назад, вбок или по кругу. Движение ресничек сводится к колебаниям в одной плоскости, вызывающим перемещение клетки или ток жидкости, если рес - нички принадлежат некоторой эпителиальной ткани. Схема движения ресничек показана на рис. 5.29.
Устройство жгутиков и ресничек весьма сходно. Они представляют
Рис. 5.28. Различные типы движения жгу - Рис. 5.29. Схема движения тика Мопаз. Стрелки указывают направле - реснички,
Ние движения клетки.
Физические проблемы, относящиеся к движению жгутика, можно четко сформулировать. Необходимо установить тонкую структуру фибриллярной системы, ее молекулярное строение и
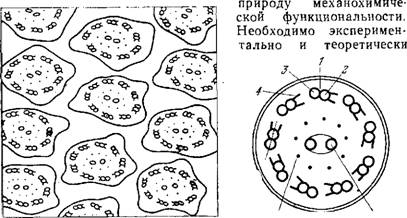
Рис. 5.30. Схема поперечных срезов Рис. 5.31. Схема строения по.
Жгутиков на основании электронной микроскопии.
Перечного среза жгутика. 1 — периферическая фибрилла,
Исследовать механику и гидродинамику специфического волнообразного движения жгутика (или реснички).
Белки жгутиков и ресничек пока недостаточно охарактеризованы. Установлена их АТФ-азная активность, показано, что и форма и частота волн зависят от концентрации АТФ. Белки являются сократительными, но их строение отличается от строения миозина или актина. Из жгутиков бактерий выделен белок флагеллин, имеющий специфический аминокислотный состав.
Качественная молекулярная модель движения жгутиков предложена в работе Сильвестра и Холвилла [164], Ранее способ распространения локальных сокращений вдоль фибриллы был рассмотрен Мэйчином [165]. Сократительная единица периферической фибриллы активируется пассивно распространяющейся волной, приходящей из соседней области жгутика. Надо понять, как химические события, вызывающие локальное сокращение, синхронизуются с механическим изгибанием. Мэйчин предполагал, что такая синхронизация возникает вследствие нелинейной связи между напряжением и деформацией сократительного элемента. Брокау также считает, что химические события инициируются механической волной [166].
В работе [164] предполагается, что сократительная единица находится в метастабильном состоянии до того, как ее достиг пик сократительной волны. Приходящая волна меняет стериче - ское и электростатическое окружение единицы, которая переходит в другое конформационное состояние и сокращается. В каждой единице происходит расщепление АТФ. Количественная оценка, основанная на биохимических данных, показывает, что в жгутике активируются 105—106 сократительных единиц в 1 с. Если они упакованы во внешних фибриллах, то расстояние между ними порядка 100 А. Скорость распространения волны

Рис. 5.32. Схема циклического Рнс. 5.33. Сдвиг и изгиб двухнитевой
Процесса в жгутике. И—нормальное состояние, S—сокращенное состояние; 1 — механическая стимуляция, 2 — сокращение, 3— релаксация.
Вдоль жгутика около 250 мкм/с, соответственно скорость стимуляции последовательных единиц есть 2,5-104 единиц в 1 с. После сокращения единица поддерживает напряжение вплоть до релаксации. На этой стадии происходит рефосфорилирование АДФ. Когда единица вновь имеет связанную АТФ, она готова опять принять участие в цикле. Следовательно, частота колебаний управляется скоростью замещения АТФ. Общая схема процесса показана на рис. 5.32 [164].
Другая точка зрения аргументируется в работе Рикменспоэла [167]. Расчет изгибающих моментов в жгутиках показывает, что активное сократительное событие сводится не к бегущей, а к стоячей волне. Изгибающий момент развивается по всей длине жгутика. Поэтому нет нужды в механизме, синхронизующем волну смещения и сократительные события.
Изгибание жгутика можно объяснить механизмом скольжения, создающим сдвиг. На рис. 5.33 изображена простая двух - нитевая модель жгутика [160, 171]. Если длины нитей, отстоящих друг от друга на р, не меняются, соотношение между сдвигом и изгибом есть
Где и — сдвиг, s—координата вдоль жгутика, 0 — угол изгиба, к — кривизна жгутика. Вводя безразмерный параметр сдвига, находим
Расчет дает линейное напряжение, равное 7-10-2 дин/см.
Если пары молекул динеина отстоят друг от друга на 170 А [172], то сила на одну его молекулу, т. е. на один мостик, равна 0,6-Ю-7 дин [167]. Это на порядок меньше величины f0 для мышцы (см. стр. 250). За один период изгибания скольжение происходит на пути 800 А [173]. Работа, совершаемая одной молекулой динеина, равна 5-Ю-13 эрг (ср. с 4,6-10~15 эрг для мышцы, стр. 250). Эта величина согласуется со значением свободной энергии расщепления АТФ [174].
Таким образом, скользящая модель жгутика представляется более привлекательной, чем модель локального сокращения [160]. Ясно, что модель должна содержать некие контрольные механизмы, генерирующие ритмический цикл изгибания. Как уже сказано, такой механизм может определяться нелинейностью системы, наличием обратной связи (см. [165]). Иными словами, скорость изгибания контролируется кривизной.
Общее выражение для распространения волны изгиба вдоль s есть периодическая функция s — Vt, где V скорость распространения волны. Следовательно,
Механизм скольжения автоматически создает разность фаз междусдвигом и кривизной, независимо от частоты колебаний [160J.
Однако этот механизм контроля не полон, так как он не учитывает вязкого сопротивления изгибу. В соответствии с идеями Мэйчина можно предположить, что кривизной контролируется не скорость нзгибання, но момент силы [160]
Где т — момент, генерируемый на единицу длины жгутнка, т0 — константа. Если к' — синусоидальная функция длины, то из (5.137) следует, что AfaKT(s)— также синусоидальная функция, но смещенная на четверть периода относительно к'.
В работе [175] (см. также [160]) с помощью компьютера было исследовано движение моделей, содержащих от 25 до 35 прямых сегментов, шарннрно соединенных друг с другом. Математическая модель основывалась на модели скольжения. Оказалось, что достаточно включить в модель два основных предположения, а именно, существование изгибающего момента, генерируемого активным процессом скольжения вдоль жгутика, н пропорциональность масштаба этого активного процесса локальной кривизне жгутнка. Тогда действительно объясняются и инициация и распространение изгиба. Варьируя параметры модели, можно получить различные волновые картины, в том числе все картины, наблюдаемые на опыте.
Имеется ряд данных, показывающих, что н внешние факторы могут влиять на движение жгутнка (см. [160]). Здесь требуются дальнейшие теоретические н экспериментальные исследования.
В работе [176] рассчитана скорость распространения волны н расход мощности на преодоление вязкого сопротивления неоднородно колеблющихся жгутиков. В работе [177] проведен анализ гипотетических форм волнового движения жгутиков — синусоидальных, меандровых н круговых дуг, соединенных прямыми линиями. Наконец, в работе [178] теоретически рассмотрено движение микроорганизмов, создаваемое трехмерным волновым движением жгутиков.
Ясно, что движение жгутиков и ресничек ставит перед физикой ряд еще не решенных проблем. Очень мало известно о детальных молекулярных механизмах, ответственных за активное скольжение н его контроль.
Как и в случае мышцы, ничего неизвестно о локализации и характере конформационных превращений в белках жгутика, более того, сами этн белки не охарактеризованы. Установлена лишь АТФ-азная нх активность. В связи с этим необходимо указать на работу Энгельгардта н Бурнашевой, выделивших из сперматозоидов сократительный АТФ-азный белок, названный имн спермазином [179]. Дальнейшие подробности, относящиеся к сократительным белкам жгутиков и ресничек, приведены в монографии Поглазова [180].
Очень важно, конечно, что установлены определенные черты сходства между движением жгутиков и мышечным сокращением. Различные типы внутренней и внешней регуляции движения еще не изучены, и изложенные в этом параграфе теоретические модели и представления имеют феноменологический характер.
• Реснички и жгутики содержат высокоупорядоченную структуру, называемую аксонемой
• Аксонема состоит из девяти наружных дуплетов микротрубочек, окружающих пару центральных микротрубочек
• Радиальные спицы представляют собой комплекс из нескольких полипептидов, которые связывают каждый наружный дуплет с центром аксонемы
• С каждым внешним дуплетом связываются динеины, домены моторов которых распространяются на примыкающие внешние дуплеты
• Динеин сдвигает внешние дуплеты по отношению друг к другу; структурные связи между другими дуплетами переводят скольжение в наклон аксонемы
• Кинезины участвуют в сборке жгутиков, транспортируя белки аксонемы в отдаленный конец жгутика
• Неподвижные первичные реснички участвуют в сенсорных процессах
Наряду с транспортировкой карго в клетках, микротрубочки также участвуют в перемещении клеток в окружающей среде. Это перемещение осуществляется с помощью ресничек и жгутиков, длинных и тонких выростов, которые отходят как волоски на поверхности многих клеток. Каждая такая органелла состоит из длинного пучка микротрубочек, окруженного выростом плазматической мембраны. Взаимодействие между микротрубочками вызывает изгиб структуры, который возможен в разных направлениях. В результате этих движений жидкость может продвигаться вдоль поверхности клеток, как показано на рисунке ниже. Для группы неподвижных клеток, например эпителиальных, это позволяет жидкой среде и различным объектам перемещаться вдоль поверхности ткани. В случае изолированных, неприкрепленных клеток они сами движутся в жидкости (т. е. плавают). Реснички и жгутики присутствуют у многих одноклеточных организмов, таких как Paramecium и Chlamydomonas (зеленая водоросль), а также находятся на сперматозоидах у большинства эукариот. У млекопитающих реснички покрывают апикальные участки некоторых эпителиальных клеток и совершают синхронные движения. При этом возникают волны движения ресничек, которые проходят по поверхности ткани. В трахее это движение используется для удаления слизи и дебриса из дыхательных путей; в яйцеводе оно обеспечивает транспортировку яйцеклетки из яичника в матку, а в мозгу создает циркуляцию цереброспинальной жидкости.
Реснички и жгутики обладают одинаковой структурой и механизмом подвижности, однако в некоторых отношениях они различаются. Наиболее существенные различия касаются длины, их количества в клетке и характера движения. Реснички короче (10-15 мкм) и количество их на клетку достигает 100 или больше. Каждая ресничка генерирует усилие за счет наклона у основания. Внешняя часть реснички остается жесткой и перегиб у основания сдвигает ее в движение, напоминающее гребок весла в воде. После этого наступает восстановительный размах, при котором перегиб реснички распространяется от основания на конец, подготавливая ресничку к следующему циклу движения. На рисунке ниже представлено движение реснички Для того чтобы проследить за отдельными стадиями движения, для регистрации использовали замедленную видеосъемку. Движения реснички столь интенсивны (много наклонов в секунду), что при съемке в обычном режиме она выглядит нечетко.
Жгутики обычно длиннее (10-200 мкм), чем реснички, и обычно клетка содержит только один или два жгутика. Они также генерируют силу при изгибе; при этом, как показано на рисунке ниже, от основания к вершине жгутика распространяется S-образная волна. Для ресничек и жгутиков характерен общий механизм движения, в основе которого лежит изгибание структуры. Различные пути распространения изгиба вдоль органеллы обусловливают различные формы волн, генерируемых ресничками и жгутиками. Поскольку два типа органелл представляют собой вариации на одну и ту же тему, мы рассмотрим их общие свойства и будем пользоваться термином жгутик для описания структуры и подвижности обеих органелл, если специально не оговаривается, что речь идет о ресничках.
При отделении от клетки жгутики продолжают биение. Это говорит о том, что оно обеспечивается самой органеллой. После удаления плазматической мембраны изолированные жгутики также продолжают биться при условии, что у них сохранился запас АТФ. Эти данные свидетельствуют о том, что движения обеспечиваются белковым содержимым жгутика и энергией гидролиза АТФ.
Внутренний белковый компонент жгутика представляет собой упорядоченную структуру, состоящую по крайней мере из 250 различных полипептидов. Эта структура называется аксонемой. Структура аксонемы жгутиков одинакова для столь различных организмов, как одноклеточное простейшее Chlamydomonas и человек.
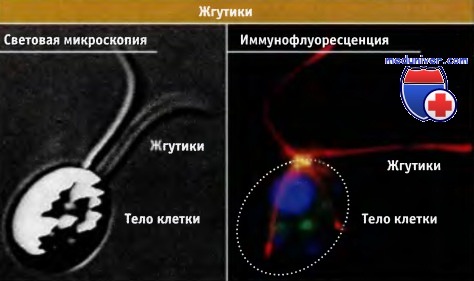
Одноклеточная водоросль Chlamydomonas reinhardtii в световом и флуоресцентном микроскопах.
В верхней части клетки видны два больших жгутика.
Во флуоресцентном микроскопе микротрубочки флуоресцируют красным; показывая, что жгутики состоят из микротрубочек.
Клетки Chlamydomonas способны передвигаться в воде за счет биений жгутиков.
Структура и расположение динеинов в остальной части аксонемы представляются сложными. В аксонеме содержится более одной формы динеина, каждая из них достаточно велика и состоит из большего числа различных полипептидов по сравнению с динеином, содержащимся в цитоплазме. Различные формы содержат один, два иди три моторных домена, и расположены в разных местах аксонемы. Смежные дуплеты микротрубочки связаны двумя наборами молекул динеина, которые называются внутренними и внешними ручками. Внешние ручки содержат только динеины с двумя или тремя головками, а белки с одной или двумя головками находятся во внутренних ручках.
Каким образом все эти связи обеспечивают подвижность жгутиков и задают соответствующий характер биения? Основной вопрос заключается в том, как функционируют динеины в этой структуре, поскольку они представляют собой моторы и с них должно начинаться движение. Для того чтобы выяснить роль динеина в обеспечении подвижности, необходимо отделить жгутики от клеток и удалить мембрану аксонемы. После этого аксонемы подвергают кратковременной обработке протеазой для того, чтобы разрушить нексиновые связи между внешними дуплетами микротрубочек. Если затем к препарату аксонем добавить АТФ, то микротрубочки сдвигаются по отношению к друг другу. Этот эффект вызывается динеинами, хвосты которых связаны с дублетом микротрубочек, генерирующих усилие в направлении от плюс- к минус-концу соседней пары. В интактной аксонеме динеин не может сдвигать внешние дублеты, поскольку они связаны между собой с помощью нексина. Поэтому усилие, генерируемое динеином, приводит к сгибанию жгутика.
Реснички и жгутики генерируют биение за счет распространения сгибательного движения по аксонеме. Сгибание начинается в основании реснички или жгутика и распространяется по направлению к дистальному концу. Сгибание происходит потому, что динеин активен только в пределах небольшого участка аксо-немы. Для того чтобы обеспечить сгибание, динеины активируются последовательно как вдоль длины, так и по периферии аксонемы. Активность динеинов контролируется центральными микротрубочками и спицами; жгутики мутантных клеток, у которых эти структуры отсутствуют, не способны совершать биение. У некоторых организмов центральные микротрубочки быстро вращаются, и в это время они могут передавать сигналы на спицы, которые, в свою очередь, активируют динеин. В центральных микротрубочках и в спицах находится несколько киназ и фосфатаз. Предполагается, что при вращении центральных трубочек запускается локальная система передачи сигнала, которая активирует динеины. Посредством быстрой локальной активации и инактивации специфических изоформ динеина аксонемы генерируют движения и регулируют силу и частоту биений.
В основании жгутика находится структура, которая называется базальным тельцем. Эти структуры обладают таким же строением, как и центриоли. Каждое базальное тело представляет собой цилиндр, состоящий из 9 триплетов микротрубочек, каждая из которых состоит или из 13 (трубочка А) или из 11 протофиламентов (трубочки В и С). Трубочки А и В служат матрицами при сборке 9 наружных дуплетов микротрубочек аксонемы. Базальное тело остается связанным с основанием аксонемы и служит для прикрепления последней к телу клетки.
Как происходит сборка жгутика, можно выяснить, удалив его с поверхности клетки и проследив образование нового. Жгутик регенерирует менее чем за час и в ходе этого процесса функционирует (т. е. осуществляет биения). Рост нового жгутика происходит на плюс-концах микротрубочек аксонемы, расположенных в дистальных участках каждого жгутика. Сборка жгутика предполагает, что необходимые компоненты аксонемы должны транспортироваться на концы структур и по мере роста собираться в аксонемы. Транспорт осуществляется в больших белковых комплексах, которые движутся в направлении кончика жгутика по наружной поверхности аксонемы, расположенной непосредственно под плазматической мембраной. Это движение называется интрафлагеллярным транспортом (IFT) и осуществляется с участием кинезина. Белковые комплексы также могут двигаться от вершины жгутика к его основанию (к минус-концам микротрубочек), однако функциональное значение этого направления транспорта неизвестно. IFT по направлению к телу клетки происходит за счет цитоплазматического динеина.
Использование наружного сегмента палочки в качестве светового сенсора может иллюстрировать широкое распространение такой возможности среди первичных ресничек. Возможно, что подобная функция первичных ресничек только начинает исследоваться. В других типах клеток содержатся более примитивные первичные реснички, в которых могут быть локализованы различные типы рецепторов. Локализация рецепторов в первичных ресничках может превратить их в некое подобие антенн, которые могут улавливать изменения в окружении и передавать эту информацию в клетку.
Некоторые редкие болезни человека связаны с мутациями, в результате которых реснички и жгутики утрачивают подвижность. Больные, которым эти мутации передаются по наследству, обычно страдают хроническими респираторными инфекциями, поскольку неподвижные жгутики не способны выводить из дыхательных путей слизь, а также захваченные патогенные микроорганизмы и раздражители. Больные мужчины часто оказываются бесплодными, так как сперматозоиды у них лишены подвижности. К числу широко известных заболеваний, связанных с неподвижностью ресничек и жгутиков, относится синдром Картагенера. Наряду с респираторными инфекциями и бесплодием, у половины больных отмечается извращенное расположение внутренних органов, при котором изменяется нормальная левосторонняя их асимметрия. Предполагается, что на одном из этапов раннего развития, до момента закладки внутренних органов, в результате биения жгутиков, в эмбрионе создается циркуляция жидкой среды, и возникает градиент секретирующих-ся веществ, способствующих морфогенезу, что определяет левостороннюю асимметрию. В отсутствие морфогенного градиента органы занимают случайное положение слева и справа. У мышей с мутациями по флагеллярным динеинам или моторам ответственным за IFT, также отмечается извращенное расположение внутренних органов. Это говорит о том, что мутации, влияющие на подвижность жгутиков или на их образование, могут приводить к порокам развития.
Биение реснички подразделяется на две фазы.
В момент удара ресничка полностью выпрямляется и проталкивает жидкость вдоль поверхности клетки.
В следующей затем фазе восстановления ресничка сгибается и возвращается в исходное положение для следующего удара. Процесс биения реснички, наблюдаемый под микроскопом.
Отмечается резкий перегиб реснички у основания, происходящий в фазе удара и ее постепенное разгибание при восстановлении. Процесс биения жгутика под микроскопом. Строение аксонемы. Показано упорядоченное расположение микротрубочек.
Микротрубочки соединены между собой различными связями, согласованное действие которых обеспечивает биение реснички.
Справа представлены электронные микрофотографии. На поперечном срезе видны внутренние и внешние динеиновые ручки,
связывающие внешние дублеты микротрубочек. Также видна одна из спиц с головкой (внизу слева). На кадрах анимации представлены два внешних дуплета (выделены золотистым цветом), соединенные динеином.
Первая часть анимации показывает, что происходит, когда из жгутиков выделяют дуплеты и селективно удаляют нексиновые линкеры.
Вторая часть показывает, что происходит в интактном жгутике. Под действием динеина и в присутствии нексиновых линкеров между дуплетами происходит перегиб жгутика. Очень длинная первичная ресничка, отрастающая от клеточной поверхности.
Внизу представлены клетки в поперечном сечении; мембраны показаны пунктиром.
Вздутие на ресничке может представлять собой карго, которое транспортируется между аксонемой и окружающей мембраной. Слева изображена зрительная палочка.
Показаны внутренний и внешний сегменты и тонкая перемычка между ними.
Справа представлена микрофотография области между двумя сегментами (выделенная на рисунке), сделанная в электронном микроскопе.
На небольшом расстоянии, вблизи от места, где она выходит из внутреннего сегмента,
перемычка выглядит как нормальная ресничка. Однако ее кончик погружен во внешний сегмент.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
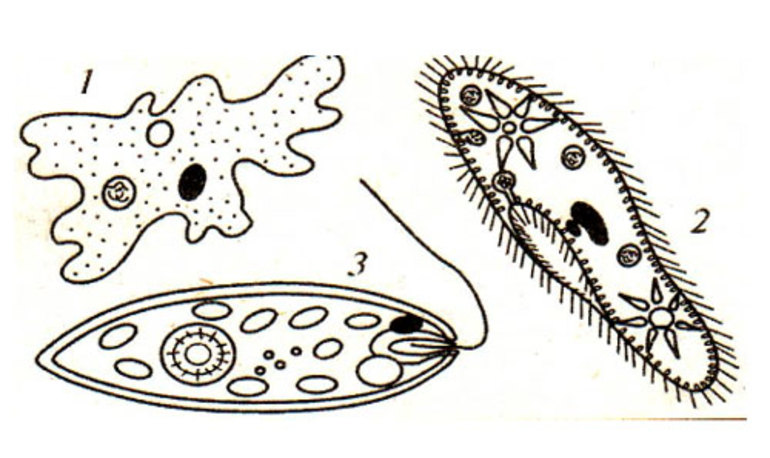
Органоиды движения — небольшие наросты на клеточной мембране, состоящие из системы микротрубочек. Они позволяют клеткам свободно перемещаться, что способствует росту живых организмов. Реснички, жгутики, псевдоподии и миофибриллы являются основными представителями органоидов движения. Строение и функции этих органелл изучаются на уроках биологии в 6 классе.
Краткая информация
Органоиды движения содержатся в растительных и животных клетках, входящих в состав многоклеточных организмов. Структура этих органелл формируется из молекул белков и фосфолипидов. Их средний размер составляет 0,25—100 мкм. В таблице перечислены основные особенности органоидов движения.
| Наименование органоида движения | Строение | Функции органоидов движения | Название одноклеточного организма |
| Жгутики | Цитоплазматические наросты, расположенные на поверхности мембраны | Передвижение клеток | Жгутиковые инфузории |
| Реснички | Тонкие выросты на эластичной структуре клетки | Очистка органов от пыли | Ресничные инфузории |
| Псевдоподии (ложноножки) | Выступы в цитоплазме клетки | Питание и передвижение организма | Саркодовые |
| Миофибриллы | Нити малой толщины | Сокращение мышц | Корненожки |
В человеческом организме присутствует большое количество ресничек и жгутиков. Они предназначены для очищения легких, защиты эпителия и стабильного функционирования репродуктивной системы. Принцип работы этих органоидов движения заключается в установлении прочных связей с клеточной мембраной.
Жгутики бактерий и архей
Жгутик — органоид движения эукариотов, обеспечивающий передвижение клеточных организмов в жидкой среде. Они содержатся в протистах, зооспорах и половых клетках. Эти органеллы представляют собой небольшие наросты, окруженные эластичной пленкой. Жгутики имеют цитоскелет, где осуществляется процесс гидролиза АТФ. Второстепенные функции жгутиков:
- формирование биологических пленок;
- обеспечение контакта клеточных организмов с субстратами;
- облегчение проникания симбиотических бактерий в клетки;
- включение защитных механизмов иммунной системы;
- предотвращение заражения клетки инфекционными вирусами.
Жгутик эукариотических клеток представляет собой комплексный структурный элемент. Он включает в себя 9 пар микротрубочек, соединенных нексиновыми мостиками. Между ними присутствует переходная зона эксонемы. В центральной части жгутика располагается ось с центриолями. На следующем рисунке описано строение органоида в разрезе.
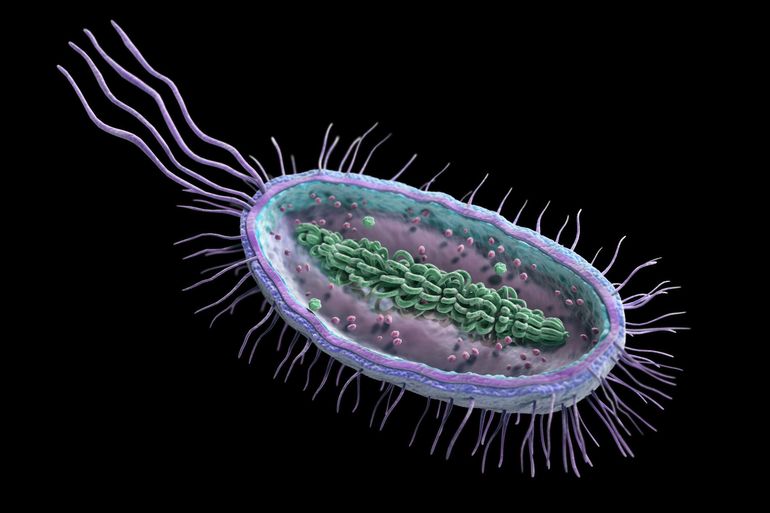
80% бактерий состоят из жгутиков. Они находятся на противоположных полюсах клеточного организма. Отличительной чертой жгутиков бактерий является их расположение в клетке. Они вмонтированы в оболочку клеточного организма. Вращение жгутика осуществляется при помощи энергии, получаемой при гидролизе АТФ. Органоид перемещается по часовой стрелке. Частотный диапазон вращения органеллы составляет от 200 до 1850 Гц. Бактериальный жгутик состоит из следующих компонентов:
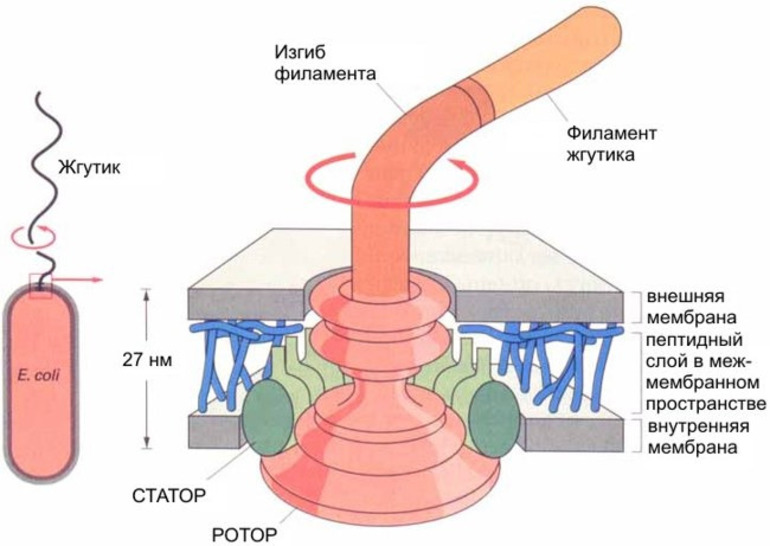
- Филамент. Представляет собой нитевидную структуру. Длина этого компонента составляет не более 14 мкм. Нить располагается за пределами цитоплазмы.
- Базальное тело. Представлено в виде муреинового чехла, окруженного мембранной оболочкой. Оно состоит из системы секреции и мотора.
- Крюк. Гибкий элемент, соединенный с филаментом и базальным телом. Его длина составляет 55 нм.
Основным рабочим элементом жгутика бактерий является филамент. Этот компонент объединяет несколько тысяч субъединиц фосфолипидов и белка. При вращении органоида филамент приобретает форму спирали, закрученной в левую сторону.
Базальное тело бактериального жгутика состоит из следующих частей:
- оси клеточного центра;
- L-кольца;
- P-кольца;
- MS-кольца;
- ротора;
- C-кольца.
Для определения характера передвижения жгутика требуется знать количество ресурсов, поставляемых основными компонентами базального тела.
Жгутики архей состоят из археллума, включающего в себя 7—13 различных генов. Структура этого органоида формируется едиными оперонами. Функцией жгутиков архей является перемещение клеточных организмов во влажной среде, но они не принимают участия в формировании биологических пленок.
Механизм работы ресничек
Реснички представляют собой тонкие органеллы в форме волоса. Находясь в неподвижном состоянии, эти органоиды выполняют роль рецепторов. Поверхность ресничек покрыта плотной цитоплазматической мембраной. В состав органеллы входит большое количество микротрубочек. В центральной части ресничек расположено базальтовое тело с центриолями.
В микротрубочках присутствуют белковые структуры, обеспечивающие стабильное скольжение ресничек. Во время передвижения органоиды совершают удары. Они предназначены для деполяризации клеточной мембраны. Характер и направление ударов зависят от процентного содержания ионов кальция в структуре ресничек.
Псевдоподии и миофибриллы
Псевдоподии представляют собой цитоплазматические выросты без плотной клеточной оболочки. В школьных учебниках эти органеллы также могут называться ложноножками. Псевдоподии сдержат простейшие организмы:
- амебы;
- арцеллы;
- фораминиферы.
Ложноножки предназначены для всасывания питательных веществ. Также к их функциям относится перемещение клеточных организмов. Скорость движения клеток составляет не более 0,2 мм/мин. Во время перемещения псевдоподии закрепляются в субстрате и захватывают частицы пищи. В результате этого процесса формируется пищеварительная вакуоль.
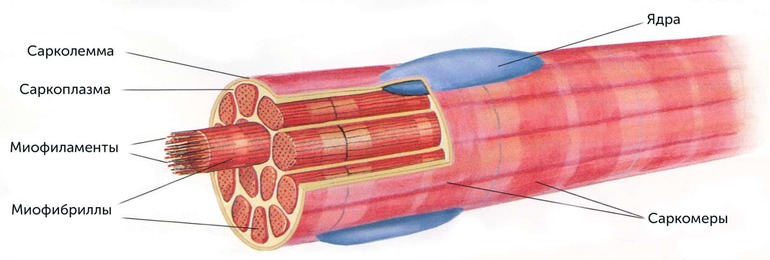
Миофибриллы представляют собой органоиды, состоящие из тонких белковых филаментов. Они располагаются в мышечном волокне и не имеют клеточной оболочки. Миофибриллы окружены саркоплазматическим ретикулумом, состоящим из саркомеров. Связь этих органелл с мышцами обеспечивается при помощи белковых нитей.
Миофибриллы обеспечивают стабильное сокращение мышечных волокон. При воздействии нервных импульсов эти органоиды начинают уменьшаться. В результате сокращения числа органоидов образуется энергия, определяющая силу мышцы. Во время силовых тренировок количество миофибрилл увеличивается. Этот процесс называется гипертрофией. Большие органоиды начинают делиться на несколько маленьких, иначе мышцы не смогут получать достаточное количество энергии.
После травмирования мышечных волокон число миофибрилл уменьшается. Этот процесс ускоряется при наличии гипса, фиксирующего мышцы в определенном положении. Это обусловлено разрушением саркомеров. В результате сокращения количества миофибрилл мышцы получают меньше энергии.
А жгутик это клеточная проекция в форме хлыста, которая участвует в перемещении одноклеточных организмов и в перемещении различных веществ в более сложных организмах.
Мы находим жгутики как в эукариотической, так и в прокариотической ветвях.Прокариотические жгутики представляют собой простые элементы, образованные единственной микротрубочкой, состоящей из субъединиц флагеллина, спирально расположенных, образующих полое ядро.
У эукариот конфигурация состоит из девяти пар микротрубочек тубулина и двух пар, расположенных в центральной области. Одним из типичных примеров жгутиков являются отростки сперматозоидов, которые придают им подвижность и позволяют оплодотворять яйцеклетку.
Реснички, другой тип удлинения клеток, имеют сходную структуру и функцию со жгутиками, но их не следует путать с жгутиками. Они намного короче и иначе двигаются.
Жгутики прокариот
У бактерий жгутики представляют собой спиральные нити, размеры которых составляют от 3 до 12 микрометров в длину и от 12 до 30 нанометров в диаметре. Они проще, чем те же элементы у эукариот.
Состав
В этих жгутиках мы можем выделить три основные части: длинную внешнюю нить, крючок, расположенный на конце нити, и базальное тело, прикрепленное к крючку.
Базальное тело имеет общие характеристики с аппаратом секреции факторов вирулентности. Это сходство может указывать на то, что обе системы унаследованы от общего предка.
Классификация
В зависимости от расположения жгутика бактерии делятся на разные категории. Если жгутик расположен на полюсах клетки как единая полярная структура на одном конце, он монотерический и если вы сделаете это с обоих концов, это амфибия.
Каждый из этих типов жгутиков также демонстрирует вариации в типах движений, которые выполняет жгутик.
Бактерии также имеют другие типы выступов на поверхности клетки. Один из них - пили, они более жесткие, чем жгутик, и бывают двух типов: короткие и обильные и длинные, участвующие в обмене. сексуальный.
Движение
Толчок или вращение бактериального жгутика является продуктом энергии, исходящей от протонно-движущей силы, а не непосредственно от АТФ.
Бактериальные жгутики не вращаются с постоянной скоростью. Этот параметр будет зависеть от количества энергии, которую клетка производит в любой момент времени. Бактерия может не только изменять скорость, но и изменять направление и движение жгутиков.
Когда бактерии направляются в определенную область, они, скорее всего, будут привлечены стимулом. Это движение известно как такси, и жгутик позволяет организму перемещаться в желаемое место.
Жгутики эукариот
Подобно прокариотическим организмам, эукариоты демонстрируют ряд процессов на поверхности мембраны. Жгутики эукариот состоят из микротрубочек и представляют собой длинные выступы, участвующие в движении и передвижении.
Более того, в эукариотических клетках может быть ряд дополнительных процессов, которые не следует путать с жгутиками. Микроворсинки - это продолжения плазматической мембраны, участвующие в абсорбции, секреции и адгезии веществ. Это также связано с моторикой.
Состав
Хотя это определение очень популярно в литературе, оно может вводить в заблуждение, поскольку в центре расположена только одна пара, а не две.
Строение микротрубочек
Микротрубочки - это белковые элементы, состоящие из тубулина. У этой молекулы есть две формы: альфа и бета тубулин. Они сгруппированы вместе, образуя димер, который будет формировать единицу микротрубочек. Компоненты полимеризуются и группируются в поперечном направлении.
Существуют различия между количеством протофиламентов, которые имеют микротрубочки, расположенные вокруг центральной пары. Один из них известен как каналец А или полный, потому что он имеет 13 протофиламентов, в отличие от канальца В, который имеет только 10-11 нитей.
Динеин и нексин
Каждая из микротрубочек прикреплена своим отрицательным концом к структуре, известной как базальное тельце или кинетосома, которая похожа по структуре на центриоль центросом с девятью триплетами микротрубочек.
Белок динеин, имеющий большое значение в движении жгутиков эукариот (АТФаза), связан двумя ветвями с каждым A-канальцем.
Нексин - еще один важный белок в составе жгутика. Он отвечает за соединение девяти пар внешних микротрубочек.
Движение
Движение происходит, когда внешние пары микротрубочек смещаются или смещаются. Динеин связан с канальцами как типа А, так и типа В. В частности, основание связано с канальцами А, а голова - с В. Нексин также играет роль в движении.
Есть несколько исследований, посвященных выяснению специфической роли динеина в движении жгутиков.
Различия между прокариотическими и эукариотическими жгутиками
Габаритные размеры
Жгутики прокариотических клонов меньше, достигают 12 мкм в длину и средний диаметр 20. Жгутики эукариот могут превышать 200 мкм в длину, а диаметр близок к 0,5 мкм.
Структурная конфигурация
Одной из наиболее ярких характеристик жгутиков эукариот является их организация микротрубочек 9 + 0 и конфигурация волокон 9 + 2. У прокариотических организмов такая организация отсутствует.
Прокариотические жгутики не покрыты плазматической мембраной, как в случае с эукариотами.
Состав жгутиков прокариот прост и включает только молекулы белка флагеллина. Состав жгутиков эукариот более сложный, он состоит из тубулина, динеина, нексина и дополнительного набора белков, а также других крупных биомолекул, таких как углеводы, липиды и нуклеотиды.
Энергия
Источником энергии жгутиков прокариот является не белок АТФаза, закрепленный в мембране, а движущая сила протона. Жгутик эукариот действительно обладает белком АТФазы: динеином.
Сходства и различия с ресничками
Сходства
Роль в передвижении
Часто встречается путаница между ресничками и жгутиками. Оба являются цитоплазматическими отростками, которые напоминают волосы и расположены на поверхности клеток. Функционально и реснички, и жгутики представляют собой выступы, которые способствуют перемещению клеток.
Состав
Оба возникают из базальных тел и имеют довольно похожую ультраструктуру. Точно так же химический состав обоих выступов очень похож.
Отличия
Длина
Решающее различие между этими двумя структурами связано с длиной: в то время как реснички имеют короткие выступы (от 5 до 20 мкм в длину), жгутики значительно длиннее и могут достигать длины более 200 мкм, что почти в 10 раз больше. чем реснички.
Количество
Когда в клетке есть реснички, их обычно бывает много. В отличие от клеток, у которых есть жгутики, у которых обычно один или два.
Движение
Кроме того, у каждой конструкции есть своеобразный механизм. Реснички двигаются мощными движениями, а жгутики - волнообразными, подобными хлыстам. Движение каждой реснички в клетке независимое, тогда как движение жгутиков скоординировано. Реснички прикреплены к волнообразной мембране, а жгутики - нет.
Сложность
Существует своеобразное различие между сложностью ресничек и жгутиков каждой структуры. Реснички - сложные проекции на все его длина, в то время как сложность жгутика ограничивается только основанием, где расположен двигатель, который управляет вращением.
Функция
Что касается их функции, то реснички участвуют в перемещении веществ в определенном направлении, а жгутики связаны только с движением.
У животных основная функция ресничек - это мобилизация жидкости, слизи или других веществ на поверхности.
Ссылки
- Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Роберт К. и Уолтер П. (2008). Молекулярная биология клетки. Наука Гарланд, Группа Тейлора и Фрэнсиса.
- Купер, Г. М., Хаусман, Р. Э. и Райт, Н. (2010). Клетка. Марбан.
- Хикман, С. П., Робертс, Л. С., Кин, С. Л., Ларсон, А., Айнсон, Х. и Эйзенхур, Д. Дж. (2008). Комплексные принципы зоологии. Нью-Йорк: Макгроу-Хилл. 14-е издание.
- Мэдиган, М. Т., Мартинко, Дж. М. и Паркер, Дж. (2004). Брок: Биология микроорганизмов. Pearson Education.
- Тортора, Дж. Дж., Функе, Б. Р., Кейс, К. Л., и Джонсон, Т. Р. (2004). Микробиология: введение (Том 9). Сан-Франциско, Калифорния: Бенджамин Каммингс.
Мавританский роман: происхождение, характеристика, представители и произведения
Читайте также: