
Роль нейтрофилов при воспалении реферат
Обновлено: 05.07.2024
а) Воспаление. При повреждении ткани независимо от его причины (бактерии, травма, химические агенты, тепло или любое другое явление) выделяется множество веществ, вызывающих существенные вторичные изменения в окружающих неповрежденных тканях. Весь комплекс этих тканевых изменений называют воспалением.
Воспаление характеризуется:
(1) расширением местных кровеносных сосудов с последующим избыточным местным кровотоком;
(2) увеличением проницаемости капилляров, что способствует утечке большого количества жидкости в интерстициальное пространство;
(3) часто — свертыванием жидкости в интерстициальном пространстве в связи с избытком фибриногена и других белков, вытекающих из капилляров;
(4) миграцией большого числа гранулоцитов и моноцитов в ткани;
(5) отеком тканевых клеток.
К тканевым продуктам, вызывающим эти реакции, относятся гистамин, брадикинин, серотонин, простагландины, несколько разных продуктов реакции системы комплемента, продукты реакции свертывающей системы крови и множество веществ, называемых лимфокинами, которые выделяются активированными Т-клетками (частью иммунной системы). Некоторые из этих веществ мощно активируют макрофагальную систему, и в течение нескольких часов макрофаги начинают уничтожать поврежденные ткани. Но иногда макрофаги повреждают еще живые тканевые клетки.
Интенсивность воспалительного процесса обычно пропорциональна степени повреждения ткани. Например, когда стафилококки проникают в ткани, они выделяют смертельно опасные клеточные токсины. В результате воспаление развивается быстро, гораздо быстрее, чем сами стафилококки могут размножиться и распространиться. Следовательно, для локальной стафилококковой инфекции характерно быстрое отгораживание, предохраняющее от распространения ее по телу.
Стрептококки, напротив, не вызывают такого интенсивного локального разрушения тканей. Следовательно, процесс отгораживания развивается медленно, на протяжении многих часов, в течение которых стрептококки успевают размножиться и мигрировать. В результате стрептококки часто проявляют гораздо более выраженную тенденцию к распространению в организме и развитию смертельных состояний, чем стафилококки, несмотря на то, что стафилококки гораздо более губительны для тканей.
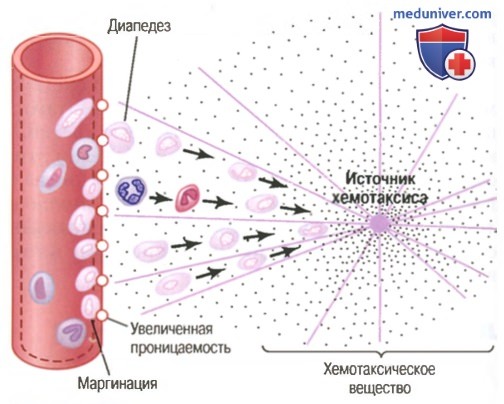
Движение нейтрофилов путем диапедеза через поры капилляров и путем хемотаксиса по направлению к области повреждения ткани
Реакции макрофагов и нейтрофилов во время воспаления
(1) изменяют внутреннюю поверхность эндотелия капилляров, способствуя прилипанию нейтрофилов к стенкам капилляров в воспаленной области. Этот эффект, показанный на рисунке выше, называют маргинацией (или краевым стоянием лейкоцитов),
(2) ведут к разрыхлению сцеплений между эндотелиальными клетками капилляров и небольших венул, обеспечивая появление достаточно больших отверстий для прохождения нейтрофилов путем диапедеза непосредственно из крови в тканевые пространства;
(3) другие продукты воспаления вызывают хемотаксис нейтрофилов в направлении поврежденных тканей, как объяснялось ранее.
Таким образом, в течение нескольких часов после начала повреждения тканей область обильно заполняется нейтрофилами. Поскольку нейтрофилы крови — уже зрелые клетки, они готовы немедленно начать свои очистительные функции для уничтожения бактерий и удаления инородного материала.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021

Сологуб Т. В., Романцова М. Г., Кремень Н. В., Александрова Л. М., Аникина О. В., Суханов Д. С., Коваленко А. Л., Петров А. Ю., Ледванов М. Ю., Стукова Н. Ю., Чеснокова Н. П., Бизенкова М. Н., Понукалина Е. В., Невважай Т. А.,
3.5. Общая характеристика и механизмы развития сосудистых реакций в очаге острого воспаления
Как известно, острое воспаление характеризуется определенной последовательностью сосудистых изменений, проявляющихся развитием спазма сосудов, артериальной, венозной гиперемии и стаза.
Спазм сосудов - реакция кратковременная. Спазм может длиться от нескольких секунд (при легком повреждении ткани) до нескольких минут (при тяжелом повреждении). Такая реакция сосудов в очаге воспаления не является строго обязательной и не проявляется в ряде случаев при повреждении паренхиматозных органов. Возникновение спазма сосудов в зоне воспаления обусловлено сокращением гладкомышечных элементов сосудов в ответ на их повреждение, а также возбуждением вазоконстрикторов под влиянием сильного, быстродействующего альтерирующего фактора. Кратковременность этой стадии определяется быстрой инактивацией вазоконстрикторных медиаторов - норадреналина, адреналина под влиянием моноаминоксидазы. Возможно, что в механизмах спазма сосудов в зоне воспаления принимает участие серотонин, освобождающийся в большом количестве при повреждении тканей из тромбоцитов и лаброцитов. Внешним проявлением спазма сосудов является побледнение участка ткани, где развивается воспалительный процесс.
Стадия артериальной гиперемии характеризуется умеренным расширением мелких артерий, артериол, капилляров, венул, а также функционированием резервных капилляров (феномен новообразования капилляров). Расширение сосудов вызывает увеличение кровенаполнения ткани, усиление лимфообразования и лимфооттока. Приток крови может несколько превышать ее отток, что способствует возрастанию гидродинамического давления в сосудах, увеличению линейной и объемной скорости движения крови, снижению артериовенозной разницы по кислороду. На этой стадии формируются такие внешние признаки воспаления, как покраснение и повышение температуры. Усиление оксигенации ткани активирует обмен веществ в клетках, способствует появлению активных форм кислорода, которые, с одной стороны, могут оказывать повреждающее действие на интактные клетки и соединительную ткань, а с другой - становятся факторами защиты организма от инфекционных патогенных агентов, а в дальнейшем оказывают стимулирующее влияние на процессы пролиферации в зоне воспаления. По мере развития артериальной гиперемии к очагу воспаления усиливается приток гуморальных факторов защиты - комплемента, пропердина, фибронектина, интерферона, церулоплазмина и др. Интенсивный кровоток обеспечивает вымывание токсических продуктов обмена, факторов патогенности, продуцируемых микроорганизмами, биологически активных веществ. В ряде случаев, при снижении реактивности организма, за счет интенсивного кровотока могут произойти диссеминация инфекции и развитие септического состояния. Артериальная гиперемия может продолжаться в течение нескольких часов и суток, она наиболее выражена по периферии зоны альтерации. В основе развития артериальной гиперемии лежат несколько механизмов [50, 53]:
1. Нейрогенный - за счет активации аксонрефлекса, усиления холинергических и гистаминергических нервных влияний на сосудистую стенку, а также повреждения сенсорных пептидергических нервных волокон, участвующих в ноцицепции и выделении сенсорных нейропептидов - субстанции Р, пептида гена, родственного кальцитонину, пептида протеинового гена. Указанные нейропептиды обладают выраженным сосудорасширяющим действием.
2. Нейропаралитический - за счет пареза симпатических вазоконстрикторов и снижения чувствительности адренорецепторов сосудов.
3. Миопаралитический - за счет снижения базального тонуса сосудов под влиянием умеренных концентраций вазоактивных соединений (гистамин, кинины, лейкотриены и др.), а также за счет Н+- и К+-гиперионии. По мере нарастания воспалительного процесса артериальная гиперемия сменяется венозной.
Венозная гиперемия характеризуется дальнейшим расширением сосудов, снижением скорости кровотока, полнокровием ткани, феноменом краевого стояния лейкоцитов и их эмиграцией, нарушением реологических свойств крови, усилением процессов экссудации.
Факторы, влияющие на переход артериальной гиперемии в венозную, можно разделить на две группы: внутрисосудистые и внесосудистые [50].
К внутрисосудистым факторам, вызывающим развитие венозной гиперемии, относятся набухание эндотелиальных клеток, краевое стояние лейкоцитов, активация системы гемостаза, сладжирование эритроцитов, сгущение крови, повышение ее вязкости, образование микротромбов.
Из внесосудистых факторов наибольшее значение имеют отек ткани и сдавление венул, мелких вен, лимфатических сосудов экссудатом. Усиливают венозную гиперемию избыточное накопление в очаге воспаления медиаторов с сосудорасширяющим действием, ферментов лизосом и ионов водорода, а также нарушение околокапиллярного соединительнотканного скелета и десмосом в зоне первичной и вторичной альтерации.
Внешними проявлениями венозной гиперемии являются отек ткани, цианоз, снижение обменных процессов и как следствие - снижение температуры в очаге воспаления. Усиление гипоксии и ацидоза стимулирует развитие соединительнотканных элементов по периферии очага воспаления и тем самым обеспечивает формирование барьера, отделяющего воспалительный очаг от здоровой ткани. В образовании барьера принимают участие также нейтрофилы и мононуклеары.
Ограничение кровотока в участке венозной гиперемии и образование барьеров способствуют уменьшению резорбции из очага воспаления продуктов распада, токсических факторов, а также снижают риск распространения инфекционных агентов. Однако при развитии воспаления в паренхиматозных органах длительная венозная гиперемия может привести к снижению их специализированных функций и развитию склерозирования клеток [7, 8].
Исходом венозной гиперемии является стаз - полная остановка кровотока в сосудах. Распространенный стаз характерен для острого, быстроразвивающегося воспаления (гиперергического). Продолжительность и исход стаза могут быть различны. Стаз может длиться от нескольких часов до нескольких дней; он может быть обратимым и необратимым [71].
Механизмы эмиграции лейкоцитов. Роль лейкоцитов в очаге воспаления
Как уже было отмечено, важнейшим признаком венозной гиперемии является эмиграция лейкоцитов из сосудов в воспаленную ткань. Последовательность выхода лейкоцитов получила название закона Мечникова, согласно которому спустя несколько часов (1, 5-2 часа) с момента действия альтерирующего фактора интенсивно эмигрируют нейтрофилы и другие сегментоядерные лейкоциты, а затем моноциты и лимфоциты.
В настоящее время известно несколько классов молекул клеточной адгезии:
1. Селектины - лектиновые молекулы клеточной адгезии, опосредующие самую раннюю стадию взаимодействия лейкоцитов и сосудистой стенки. Селектины экспрессируются рано и не обеспечивают прочного прикрепления лейкоцитов к эндотелию сосудов. Выделяют три вида селектинов: L-селектины синтезируются всеми лейкоцитами,
Е-селектины синтезируются эндотелиоцитами. Указанные селектины обеспечивают феномен краевого стояния лейкоцитов. Источником
P-селектинов являются тромбоциты и эндотелий сосудов, помимо участия в процессах маргинации лейкоцитов, они способствуют адгезии тромбоцитов.
2. Интегрины - это гетеродимерные белки, экспрессируемые различными лейкоцитами и клетками гемопоэтического ряда. Эндотелий экспрессирует лишь некоторые интегрины. В настоящее время идентифицировано пять видов интегринов. Наиболее важными для миграционного и фагоцитарного процесса являются три гетеродимера. Указанные интегрины ответственны за поздние стадии взаимодействия лейкоцитов, тромбоцитов и эндотелия сосудов.
3. Адгезивные молекулы суперсемейства иммуноглобулинов (ICAM) - трансмембранные протеины с пятью внеклеточными доменами, экспрессируемые преимущественно эндотелием. Некоторые из этих молекул синтезируются неактивированными клетками и определяют интенсивность физиологической миграции лейкоцитов. В зоне воспаления под влиянием ИЛ-1, ФНО, гамма-интерферона активируются синтез и экспрессия молекул ICAM, что обеспечивает отсроченную фазу адгезии и эмиграции лейкоцитов. В группу белков, подобных иммуноглобулинам, включены лимфоцитарные рецепторы - СD2, СD58 (LFA-3). Они экспрессируются на Т-лимфоцитах и антигенпредставляющих клетках, участвуют в презентации антигена и контакте цитотоксических лимфоцитов с клетками-мишенями.
4. Адрессины - белки эндотелиальных клеток венул лимфоидных органов. Имеют большое значение в миграции лимфоцитов в очаги хронического и аутоиммунного воспаления. При остром воспалении адрессины менее значимы.
5. Хрящевые соединительные белки. Экспрессируются всеми видами лейкоцитов, распознают гиалуроновую кислоту, что обеспечивает перемещение лейкоцитов в основном веществе соединительной ткани [16, 18].
При недостаточном образовании селектинов и интегринов или выработке антител к мембранным рецепторам снижаются адгезивные свойства лейкоцитов, нарушается их способность к эмиграции, накоплению в очаге воспаления и фагоцитозу, что может способствовать частым гнойным инфекциям и развитию сепсиса.
Важная роль в процессах адгезии и эмиграции принадлежит устранению отрицательного заряда эндотелиальных клеток и лейкоцитов за счет накопления в очаге воспаления одновалентных и двухвалентных ионов (Н, К, Са, Mn, Mg) , а также катионных белков, выделяемых лейкоцитами. Факторами инициации адгезии лейкоцитов к стенке сосуда являются комплемент (С5а), лейкотриены, фибронектин, иммуноглобулины (Fc-фрагмент IgG), гистамин.
После адгезии происходят перемещение лейкоцита по поверхности эндотелиоцита к межэндотелиальной щели, которая в очаге воспаления значительно расширена, а затем и образование ложноножки и передвижение лейкоцита через межэндотелиальную щель в подэндотелиальное пространство и базальную мембрану сосуда. Таким образом лейкоциты оказываются за пределами сосуда.
В большинстве случаев острого воспаления внутрисосудистые перемещения лейкоцитов и их эмиграция занимают несколько часов. Как правило, первыми в очаг воспаления выходят нейтрофилы, они обнаруживаются в воспаленной ткани уже через 6-24 часа. Несколько позднее эмигрируют моноциты и лимфоциты (24-48 часов). Такая асинхронность эмиграции клеток обусловлена неодновременным появлением молекул адгезии и хемотаксических факторов, специфичных для разных лейкоцитов.
Нужно отметить, что временные интервалы эмиграции лейкоцитов и последовательность их выхода весьма относительны и определяются типом сосуда, видом воспаления и стадией воспаления.
Так, в зоне воспалительного процесса, инициируемого возбудителями туберкулеза, листериоза, хламидиоза, токсоплазмоза, вирусной инфекцией, первоначально в очаге воспаления доминируют мононуклеары, при аллергических реакциях - эозинофилы.
Направленное движение лейкоцитов обеспечивается хемоаттрактантами, концентрация которых по мере развития вторичной альтерации в очаге воспаления нарастает, а также наличием рецепторов к различным хемоаттрактантам на поверхности лейкоцитов.
При взаимодействии рецепторов и хемоаттрактантов возникает гиперполяризация мембраны лейкоцита, увеличивается ее проницаемость для ионов Са, инициируется синтез функционально активных фосфолипидов и циклических нуклеотидов, растет число внутриклеточных органелл, функционирование которых обеспечивает перемещение лейкоцита и секрецию содержимого гранул. Помимо ориентированного движения лейкоцитов, хемоаттрактанты индуцируют адгезию лейкоцитов к эндотелию, способствуют краевому стоянию лейкоцитов и их агрегации в просвете микрососудов. Хемотаксис опосредуется различными группами веществ:
1. Цитотаксигенами, которые, не являясь хемоаттрактантами, генерируют факторы хемотаксиса.
2. Цитотаксинами, которые оказывают прямое воздействие на лейкоциты. Некоторые цитотаксины специфичны для нейтрофилов (например, экстракт культуры Е.coli) , а другие цитотаксины индуцируют миграцию гранулоцитов и моноцитов [35].
Хемоаттрактанты могут иметь эндогенное и экзогенное происхождение. Экзогенными хемоаттрактантами являются пептиды бактерий, особенно содержащие N-формиловые группы. Свойствами эндогенных хемоаттрактантов обладают компоненты комплемента, иммуноглобулины, иммунные комплексы, лимфокины и монокины, кинины, лейкотриен В4, продукты липоксигеназного пути превращения арахидоновой кислоты, фактор, активирующий тромбоциты, фактор Хагемана, лизосомальные ферменты и другие.
Роль лейкоцитов в очаге воспаления
Эмигрировавшие в зону воспаления нейтрофилы являются активными фагоцитами, которые очищают зону воспаления от инфекционных возбудителей. Адгезия нейтрофилов к объекту фагоцитоза ускоряется благодаря опсонинам - активным белковым молекулам, прикрепляющимся к объекту и облегчающим распознавание объекта фагоцитирующими клетками. Одновременно с процессами направленного движения лейкоцитов и фагоцитозом в них происходит респираторный взрыв - резкое увеличение потребления кислорода для образования бактерицидных свободных кислородных радикалов (синглетный кислород, гидроксильный радикал, перекись водорода, супероксидный анион). Образование активных форм кислорода происходит с участием ферментов миелопероксидазы, супероксиддисмутазы и каталазы. Освобождающиеся в очаге воспаления активные формы кислорода являются высокотоксичными факторами для бактерий, грибов, микоплазм, вирусов, хламидий и других возбудителей, они нарушают структуру и функции мембран микробных клеток, ограничивают их жизнедеятельность или вызывают гибель микроорганизмов. Помимо антимикробной активности, усиление свободнорадикальных процессов вызывает повреждение интактных паренхиматозных клеток, эндотелиальных клеток сосудов и элементов соединительной ткани в очаге воспаления, что способствует дальнейшей альтерации ткани.
Кроме вновь синтезирующихся факторов оксидантной системы, в гранулах нейтрофилов содержатся лизоцим, лактоферрин, катионные белки, щелочная и кислая фосфатазы, рибонуклеаза, дезоксирибонуклеаза, гиалуронидаза, b-глюкуронидаза, эластаза, коллагеназа, ФАТ, кинины, лейкоцитарный пироген, хемотаксические факторы.
Таким образом, накопление нейтрофилов в очаге воспаления и освобождение ими указанных биологически активных веществ вызывают гибель или ограничение жизнедеятельности микроорганизмов, разрушение и лизис омертвевших тканей, очищение зоны повреждения [57]. За счет нейтрофилов вокруг очага воспаления формируется нейтрофильный защитный барьер, который отграничивает зону повреждения (совместно с моноцитарным и фибробластическим) от здоровой ткани и препятствует распространению инфекции и токсических повреждающих факторов.
Высокоактивные медиаторы нейтрофилов участвуют в развитии вторичной альтерации, стимулируют выход биологически активных веществ из других клеток, способствуют расширению сосудов, увеличению их проницаемости, экссудации плазмы и эмиграции лейкоцитов. Поступая в системный кровоток, они вызывают проявление некоторых системных эффектов воспалительного процесса.
Эмигрировавшие в зону воспаления эозинофилы также обладают способностью фагоцитировать бактерии, грибы, иммунные комплексы. Они содержат примерно такой же набор ферментов, как и нейтрофилы (кроме лизоцима). Однако в фагоцитозе участвует меньший процент эозинофилов, и он протекает менее интенсивно, чем у нейтрофилов.
В очаге воспаления эозинофилы выполняют две основные функции: они становятся модуляторами реакций гиперчувствительности и главным механизмом защиты от личиночных стадий паразитарных инфекций.
Основными факторами, стимулирующими процесс дегрануляции эозинофилов, являются иммуноглобулины, иммунные комплексы, комплемент. Продукты секреции эозинофилов ингибируют выделение лаброцитами гистамина и участвуют в его инактивации за счет гистаминазы, эозинофильный катионный протеин связывает и нейтрализует гепарин, высокие концентрации арилсульфатазы инактивируют хемотаксические белки и медленно реагирующую субстанцию анафилаксии, фосфолипазы В и D инактивируют тромбоцитактивирующий фактор. Под влиянием эндотоксинов и в процессе фагоцитоза эозинофилы продуцируют и выделяют эндогенный пироген, обеспечивающий развитие лихорадочной реакции, сопровождающей воспалительный процесс.
Медиаторы эозинофилов, так же как и нейтрофилов, могут участвовать в реакциях повреждения ткани и распространении вторичной альтерации.
Базофильные лейкоциты, как и другие гранулоциты, обладают способностью к фагоцитозу, но их фагоцитарная активность довольно низка. В зоне инфекционного и аллергического воспаления возникает реакция дегрануляции базофилов с выделением медиаторов воспаления - гистамина, гепарина, фактора активации тромбоцитов, лейкотриенов, калликреина, эозинофильного хемотаксического фактора, ферментов.
При наличии этиологического агента в виде бактериального возбудителя моноциты/макрофаги являются антигенпрезентирующими клетками, обеспечивающими запуск иммунного ответа организма. В процессе представления антигена активированные макрофаги секретируют и освобождают такие цитокины, как ФНО, интерлейкины (ИЛ-1, ИЛ-3, ИЛ-6, ИЛ-8, ИЛ-10, ИЛ-12, ИЛ-15), компоненты комплемента, хемотаксические факторы [44]. Выделение зрелых форм интерлейкинов из клеток начинается уже через 2 часа после антигенной стимуляции и продолжается в течение 24-48 часов. Моноциты/макрофаги в зоне воспаления являются источником лизосомальных протеаз - коллагеназы, эластазы, ангиотензин конвертазы, активатора плазминогена; адгезивных веществ - фибронектина, тромбоспондина, протеогликанов; факторов роста - колониестимулирующих факторов, фактора роста фибробластов; эндогенных пирогенов.
Под действием пусковых медиаторов макрофагов в очаге воспаления происходит активация различных типов лейкоцитов, клеток соединительной ткани, эндотелия сосудов, паренхиматозных клеток, что приводит к синтезу и секреции биологически активных соединений, оказывающих повреждающие, защитные и системные эффекты.
Интенсивная эмиграция лимфоцитов в зону воспаления в большинстве случаев осуществляется после эмиграции нейтрофилов и моноцитов. Стимулированные лимфоциты выделяют биологически активные вещества - лимфокины, обеспечивающие развитие иммунного ответа, аллергических реакций, процессов пролиферации и репарации.
Все лейкоциты в зоне воспаления довольно быстро подвергаются жировой дегенерации, превращаются в гнойные тельца и удаляются вместе с гноем. В очаге воспаления возможны два пути гибели лейкоцитов.
Первый - грубое повреждение мембраны, нарушение целостности клетки и ее некроз, что возможно в очаге первичной и вторичной альтерации под влиянием самого повреждающего агента и различных цитотоксических медиаторов воспаления.
Второй - более мягкий, запрограммированный путь клеточной гибели, или апоптоз. При апоптозе наблюдаются уменьшение ядра лейкоцита, конденсация ядерного хроматина, фрагментация ДНК, подавление транскрипции, снижение синтеза рибосомной РНК и белков. Стареющие нейтрофилы теряют способность секретировать лизосомальные ферменты, и это исключает дальнейшее повреждение ткани. Макрофаги активно фагоцитируют апоптозные нейтрофилы. Поглощение апоптозных нейтрофилов обеспечивается взаимодействием тромбоспондина, секретируемого макрофагом и рецепторами нейтрофила [62, 67]. Путем апоптоза могут удаляться из очага воспаления не только нейтрофилы, но и эозинофилы и лимфоциты.

1. Зайчик А.Ш. Патологическая физиология. Том 2. Патохимия [Текст]: учеб. / А.Ш. Зайчик, Л.П. Чурилов. – 3-е изд. – СПб.:ЭЛБИ-СПб, 2007. – 688 с.
3. Лекция 4. Особенности структуры, функции и метаболизма В- и Т- систем лимфоцитов / Н.П. Чеснокова, Е.В. Понукалина, Т.Н. Жевак [и др.] // Международный журнал фундаментальных и прикладных исследований. 2015. № 4. – С. 293 -297.
4. Патологическая физиология [Текст]: учеб. / под общ. ред. В.В.Моррисона, Н.П. Чесноковой. – 4-е изд. – Саратов: Изд-во Сарат. гос. мед. ун-та, 2009. – 679 с.
6. Типовые патологические процессы как основа патогенеза болезней различной этиологии [Текст]: монография / под общ. ред. В.М. Попкова, Н.П. Чесноковой. – Саратов: Изд–во Сарат. гос. мед. ун-та, 2017. – 536 с.
7. Типовые реакции иммунной системы на действие антигенов-аллергенов [Текст]: учеб. пособие / под ред. Н.П. Чесноковой. – Саратов: Изд-во Сарат. гос. мед. ун-та, 20014. – 156 с.
8. Цитокины: биологическая роль в развитии реакций адаптации и повреждения в условиях нормы и патологии различного генеза [Текст]: монография / под общ. ред. В.М. Попкова, Н.П. Чесноковой. – Саратов: Изд-во Сарат. гос. мед. ун-та, 2016. – 448 с.
Воспаление – типовой патологический процесс, составляющий основу патогенеза болезней с разнообразной локализацией патологии и симптоматикой. Воспаление может развиваться остро или хронически. При остром развитии воспалительного процесса возникает стереотипный комплекс сосудистых изменений в виде спазма сосудов, артериальной и венозной гиперемии.
Несколько опережая во времени развитие сосудистых реакций, а затем и параллельно с сосудистыми изменениями формируются тканевые расстройства в виде альтерации, экссудации, пролиферации. Указанные сосудистые и тканевые изменения имеют место при различных заболеваниях воспалительной природы – стоматитах, гингивитах, глосситах, паро- и перидонтитах, эзофагитах, энтеритах, миокардитах и так далее, то есть развиваются не зависимо от локализации воспаления [1, 4, 5, 6, 8].
Касаясь характеристики фазы альтерации, следует отметить морфологические и биохимические аспекты ее развития.
Альтерация – обобщающий термин, означающий повреждение. С точки зрения морфологии, это зона некроза, дистрофии, апоптоза, отека тканей и других деструктивных процессов. С метаболической, патохимической, функциональной сторон, это стадия освобождения медиаторов альтерации и проявления их биологических эффектов. По времени развития различают первичную и вторичную альтерацию.
Первичная альтерация формируется в момент воздействия патогенного фактора на организм, ее границы соответствуют зоне первичного действия патогенов.
Вторичная альтерация отсрочена во времени, связана с диффузией из зоны первичной альтерации в окружающую ткань токсических и ферментных факторов патогенности возбудителей бактериальной, вирусной, паразитарной природы, а также медиаторов альтерации гуморального и клеточного происхождения, предсуществующих или вновь синтезируемых.
К основным источникам предсуществующих медиаторов альтерации относятся тучные клетки, а также базофилы, в процессе дегрануляции которых освобождается гистамин, гепарин, серотонин, факторы активации тромбоцитов, хемотаксиса эозинофилов, нейтрофилов, а затем, в процессе антигенной стимуляции, продуцируются интерлейкины и другие биологически активные вещества.
В момент действия альтерирующих факторов терминалями нервных волокон освобождаются нейропептиды, такие, как субстанция P, пептид гена родственного кальцитонину, формирующие чувство боли.
В процессе дегрануляции тромбоцитов, высвобождаются катехоламины, серотонин, лизосомальные гидролазы, факторы гемостаза и фибринолиза.
К числу предсуществующих медиаторов альтерации клеточного происхождения относятся лизосомальная гидролаза, а также лизоцим, катионные белки, лактоферрин. Основными источниками которых являются клетки моноцитарно-макрофагальной системы и гранулоциты.
Типовой реакцией на повреждение в зоне воспаления, в том числе в тканях пародонта, является повышение проницаемости биологических мембран, в частности цитоплазматических, митохондриальных, лизосомальных и др.
Альтерирующие факторы различной биологической, а также, физической и химической природы инициируют активацию таких ферментов в зоне повреждения, как липоксигеназа и циклооксигеназа, обеспечивающих развитие каскада реакций образования, соответственно, лейкотриенов и простогландинов. Последние обладают мощным вазоактивным действием, регулирует тонус и проницаемость сосудистой стенки, гемостатические реакции, вызывая развитие экссудации и эмиграции лейкоцитов на стадии венозной гиперемии.
Важная роль в развитии воспаления принадлежит вновь синтезируемым медиаторам таким как лимфокины и монокины, вызывающим развитие синдрома системного воспалительного ответа, направленного в значительной мере на зобилизацию защитно-приспособительных реакций за счет вовлечения иммунной системы, гормонального баланса, метаболических изменений [1, 4, 5, 6, 7].
В настоящее время известно более 100 биологически активных веществ, секретируемых моноцитами/макрофагами, классификация которых приведена ниже:
1) протеазы: активатор плазминогена, коллагеназа, эластаза, ангиотензин-конвертаза;
2) медиаторы воспаления и иммуномодуляции:ФНО, ИЛ-1, ИЛ-3, ИЛ-6, ИЛ-8, ИЛ-10, ИЛ-12, ИЛ-15, ИФН, лизоцим, фактор активации нейтрофилов, компоненты комплемента;
3) факторы роста: КСФ-ГМ, КСФ-Г, КСФ-М, фактор роста фибробластов, трансформирующий фактор роста;
4) факторы свертывающей системы и ингибиторы фибринолиза: V,VII, IX, X, ингибиторы плазминогена, ингибиторы плазмина;
5) адгезивные вещества: фибронектин, тромбоспондин, протеогликаны.
Лимфокины – это биологически активные вещества, которые продуцируются сенсибилизированными лимфоцитами при специфической антигенной стимуляции [2, 3, 7, 8].
Следует отметить, что часть цитокинов (ИЛ-2, ИЛ-3, ИЛ-6, ИЛ-8, ИЛ-10 и др.) могут продуцироваться лимфоцитами, а также клетками моноцитарномакрофагальной системы. В связи с этим они могут быть отнесены как к группе лимфокинов, так и к группе монокинов.
Лимфокины проявляют многообразную биологическую активность, обеспечивая бластогеный и митогенный эффекты на лимфоидную и моноцитарно-макрофагальную систему, развитие хемотаксиса нейтрофилов, макрофагов, лимфоцитов, вызывая антивирусные и антимикробные эффекты.
К числу вновь синтезируемых медиаторов в зоне воспаления клеточного происхождения относят свободные радикалы, такие как супероксид-анион-радикал, гидроксильный радикал, перекись водорода, оксид азота, имеющие как внутриклеточное, так и внеклеточное происхождение в условиях формирования гипоксии, ишемии, тромбоза, эмболии в зоне воспаления, а также при различных стрессорных ситуациях.
Свободные радикалы взаимодействуют со структурными белками клеток, с липидными компонентами биологических мембран с образованием эндоперекисей с последующей дестабилизацией биосистемы.
Особую роль вразвитии воспаления отводят оксиду азота, который является мощным вазодилятатором, ингибитором агрегации тромбоцитов, нейротрансмиттером неадренергически-нехолинергических нейронов, вызывающих релаксацию гладкой мускулатуры ряда органов и тканей, в частности половых органов.
При остром воспалении свободные радикалы вызывают разрушение межклеточного матрикса, оказывают повреждающее действие на фибробласты, в то же время они могут оказывать стимулирующее влияние на процессы пролиферации [1, 4, 5, 6, 7].
К числу факторов антиоксидантной защиты тканей относятся ферменты: каталаза, супероксиддисмутаза, глутатионпероксидаза, а также витамин К, α-токоферол, метионин и др.
Помимо клеточных медиаторов воспаления большую роль в развитии вторичной альтерации играют плазменные медиаторы: кинины, компоненты системы комплемента, факторы обертывания крови.
Относительно механизмов активации медиаторов альтерации гуморального происхождения следует отметить, что деструкция эндотелия сосудов в зоне альтерации приводит к обнажению субэндотелиальной выстилки с выраженными прокоагулянтными свойствами за счет наличия коллагена, эндотелина, простагландинов F2α, тромбоксанов, фибронектина и других компонентов. При этом возникает инициация внешнего и внутреннего механизмов активации протромбиназы и соответственно систем гемостаза, фибринолиза, комплемента, калликреин-кининовой системы, тромбоцитарного звена гемостаза.
Одновременно с тканевыми изменениями в зоне воспаления в тканях пародонта развивается комплекс сосудистых реакций. При кратковременном действии достаточно сильного повреждающего фактора первоначально возникает спазм сосудов вследствие раздражения вазоконстрикторных нервных окончаний и гладкомышечных элементов сосудистых стенок. Спазм сосудов очень быстро сменяется артериальной гиперемией – увеличением кровенаполнения воспалительного очага за счет усиления притока крови в условиях накопления умеренного количества биологически активных веществ – медиаторов воспаления, активации аксон-рефлекса и повреждения вазоконстрикторных нервных окончаний. Артериальная гиперемия характеризуется умеренным расширением артериол, капилляров и венул, увеличением скорости кровотока, феноменом новообразования капилляров, активацией обменных процессов в тканях [1, 4, 5].
В последующем артериальная гиперемия сменяется венозной вследствие нарушения оттока крови из очага воспаления. Этому способствует уменьшение просвета сосудов за счет набухания эндотелия, краевого стояния лейкоцитов, сгущения крови, агрегации тромбоцитов, а также сдавление венул и лимфатических сосудов экссудатом. Для венозной гиперемии характерно значительной расширение сосудов микроциркуляторного русла, замедление скорости кровотока, снижение оксигенации тканей очага воспаления. На стадии венозной гиперемии создаются условия для развития последующих тканевых изменений: экссудации и эмиграции.
Экссудация (от лат. ex-sudare – потеть) – выпотевание белоксодержащей жидкой части крови через сосудистую стенку в воспаленную ткань.
В основе развития экссудации при воспалении лежат следующие механизмы:
1) повышение проницаемости сосудов в результате воздействия медиаторов воспаления, ионов водорода, лизосомальных ферментов;
2) возрастание гидродинамического давления в сосудах очага воспаления вследствие увеличенного кровенаполнения;
3) возрастание осмотического и онкотического давления в воспаленной ткани в результате альтерации;
4) снижение онкотического давления крови из-за потери белков в условиях повышенной проницаемости сосудистых;
5) диффузия жидкой части крови через микропоры в эндотелиальных клетках (трансцеллюлярные каналы), а также активный перенос микровезикул при цитопемсисе.
Ведущим фактором экссудации является повышение проницаемости сосудов. К медиаторам воспаления, влияющим на проницаемость сосудистой стенки, относятся гистамин, серотонин, брадикинин, С5а и С3а, лейкотриены С4 и Д4, которые влияют непосредственно на эндотелиальные клетки, вызывая их контракцию, а также С5а de Arg, лейкотриен В4, интерлейкин-1, фактор активирующий тромбоциты [4, 5, 6, 7, 8].
Повышение проницаемости сосудов при воспалении является двухфазным. Первая фаза, или немедленная, возникает вслед за действием альтерирующего агента, достигает максимума на протяжении нескольких минут и продолжается в течение 5-30 минут. Вторая фаза, замедленная, развивается постепенно, достигает максимума через 4-6 часов и длится иногда до 10 часов в зависимости от вида и интенсивности воспаления.
Степень повышения проницаемости сосудов при воспалении определяет не только интенсивность экссудации, но и качественный состав экссудата, содержащего большее, чем в транссудате, количество белка, биологически активных веществ. В зависимости от качественного состава выделяют следующие виды экссудатов:
Эмиграция – выход форменных элементов белой крови за пределы сосудистого русла в очаг воспаления. Последовательность выхода лейкоцитов в зону воспаления получила название закона И. И. Мечникова, согласно которому через 1,5-2 часа с момента действия альтерирующего агента интенсивно эмигрируют нейтрофильные лейкоциты, которые выходят через поры между эндотелиальными клетками. Через 18–20 часов в зоне воспаления появляются моноциты, а затем лимфоциты, которые эмигрируют путем цигопемсиса и частично через поры между эндотелиальными клетками. При поражении тканей пародонта значительно усиливается эмиграция гранулоцитов и мононуклеаров не только в очаг воспаления, но и в ротовую полость. Выход форменных элементов в полость рта осуществляется через зубодесневой желобок из посткапиллярных венул маргинальной части десен и межзубных сосочков, расположенных непосредственно под эпителием [4, 5, 6, 7, 8].
Эмиграция лейкоцитов обусловлена, во-первых, действием хемоаттрактантов, обусловливающих развитие положительного хемотаксиса. В роли хемоаттрактантов могут выступать продукты специфических реакций в организме: компоненты комплемента (С3а, С5а, С567), лимфокины, цитофильные антитела, иммунные комплексы. В другую группу входят неспецифические эндогенные цитотаксины, выделяемые из поврежденных, инфицированных или активированных клеток, – калликреин, фибринопептид В, коллаген, лейкоэгрезин, цАМФ и др. Третью группу составляют экзогенные хемоаттрактанты, выделяющиеся в процессе жизнедеятельности различными микроорганизмами.
Миграции из кровотока и положительному хемотаксису лейкоцитов в полость рта способствует наличие мощных хемотаксических агентов бактериальных зубных бляшек.
Формирование в очаге воспаления положительного потенциала альтерации способствует эмиграции лейкоцитов под действием электрокинетических сил.
Выходу лейкоцитов способствует высокая проницаемость сосудов и процесс экссудации.
Эмигрировавшие элементы белой крови в зоне воспаления выполняют ряд функций. Нейтрофильные лейкоциты и моноциты обеспечивают фагоцитоз, являющийся одним из важнейших механизмов неспецифической резистентности организма. В то же время лейкоциты в зоне воспаления являются источником целого ряда бактерицидных факторов и биологически активных веществ – миелопероксидазы, лизоцима, лактоферрина, катионных белков, кининов, простагландинов, лизосомальных ферментов, хемотаксических факторов. Активные гранулоциты и моноциты продуцируют эндогенные пирогенные факторы, вызывающие развитие лихорадки, обеспечивающей реализацию комплекса защитно-приспособительных реакций в организме. Моноциты, относящиеся к категории макрофагов, принимают участие в инициировании иммунного ответа при воспалении, обеспечивая подготовку антигенов переработку их в иммуногенную форму. Таким образом, эмигрировавшие в зону воспаления моноциты принимают участие в специфических механизмах реагирования на действие патогенного фактора. Лимфоциты являются источником лимфокинов и принимают непосредственное участие в иммунологических механизмах защиты [2, 7, 8].
Исходом венозном гиперемии является развитие престаза и стаза – полной остановки кровотока в микроциркуляторном русле.
Пролиферация является завершающей фазой развития воспаления, обеспечивающей репаративную регенерацию тканей на месте очага альтерации. Процессу пролиферации предшествует формирование нейтрофильного и моноцитарного барьеров, фагоцитоз инфекционных возбудителей воспаления клеточного детрита. По периферии очага воспаления формируется фибробластический барьер, за счет которого зона воспаления инкапсулируется и возникает постоянная миграция клеточных и неклеточных элементов соединительной ткани и формирование соединительнотканного остова на месте первичной и вторичной альтерации. Одновременно в ряде тканей происходит размножение и органоспецифических элементов с последующим полным или частичным восстановлением структуры и функции поврежденной ткани [4, 5, 6, 7, 8].
Локальные расстройства кровоснабжения и обменных процессов в зоне воспаления, как правило, сочетаются с комплексом метаболических и функциональных расстройств на уровне целостного организма.
Синдром системного воспалительного ответа
Синдром системного воспалительного ответа – это комплекс цитокин-опосредованных системных морфологических, функциональных и биохимических изменений в организме, возникающих на фоне развития воспалительно-деструктивных процессов инфекционной, аллергической, онкологической, травматической природы, . стрессорного воздействия.
Основными признаками ССВО, помимо гормонального дисбаланса, являются: увеличение содержания в крови белков острой фазы, развитие лейкоцитоза более 12000 мкл, в тяжелых случаях возможна лейкопения – менее 4000 мкл.
Одновременно развивается температурная реакция выше 38 °С под влиянием провоспалительных цитокинов – вторичных эндопирогенов (ИЛ1, ИЛ6, ИЛ8, ФНОα, КСФ). Активация симпатоадреналовой системы приводит к развитию тахикардии, параллельно возникает тахипноэ.
Наиболее достоверным и хорошо изученным признаком острой фазы ответа является изменение белкового состава крови.
Белки острой фазы синтезируются главным образом в печени и делятся на две группы:
1. Позитивные маркеры (α1-антихемотрипсин, С-реактивный белок, фракция С3 комплемента, церулоплазмин, фибриноген, гаптоглобин, орозомукоид, плазминоген, антитромбин III), уровень которых возрастает.
2. Негативные маркеры (альбумин, трансферин), уровень которых снижается.
Уровень позитивных маркеров острой фазы нарастает в течение 24-48 часов после воздействия чрезвычайного альтерирующего фактора, причем в случае травматического повреждения степень увеличения позитивных маркеров в крови коррелирует с тяжестью повреждающего воздействия. При неосложненном течении патологии уровень позитивных маркеров острой фазы нормализуется в течение 72-96 часов. Однако в случае развития септических заболеваний, хронических воспалительных процессов, у больных с осложненным послеоперационным периодом уровень позитивных маркеров острой фазы длительное время может оставаться повышенным. В этих случаях большую прогностическую ценность представляет мониторинг позитивных маркеров острой фазы.
Наиболее чувствительным критерием системного действия на организм острой воспалительной реакции является возрастание концентрации в крови С-реактивного белка, в связи с чем наиболее распространен в клинической практике мониторинг С-реактивного белка. Последний является полипептидом с ММ 120 кД, обладает способностью связываться с полисахаридом С на поверхности одной из разновидностей стрептококка. Увеличение продукции С-реактивного белка печенью сочетается с последующим развитием комплекса системных реакций, в частности, индуцируемой этим белком активацией комплемента, подавлением функции тромбоцитов и лимфоцитов, торможением ретракции сгустка, стимуляцией фагоцитарной активности нейтрофилов.

Студент медицинского факультета УЛГУ. Интересы: современные медицинские технологии, открытия в области медицины, перспективы развития медицины в России и за рубежом.
- Запись опубликована: 25.11.2020
- Время чтения: 1 mins read
Фагоцитоз – это способность определенных клеток (фагоцитов) удерживать и переваривать плотные частицы. Это явление было открыто И. Мечниковым.
Фагоцитоз осуществляется микрофагами (нейтрофилами) и системой мононуклеарных макрофагов.
Мононуклеарные макрофаги
Система (MMS) включает следующие элементы:
- промоноциты (костный мозг);
- моноциты (кровь);
- тканевые макрофаги;
- гистиоциты соединительной ткани;
- печеночные клетки Куппера;
- легочные альвеолярные макрофаги;
- свободные и фиксированные макрофаги лимфатической ткани;
- плевру и перитонеальные макрофаги.
Клетки мононуклеарной системы объединяет общее происхождение гемопоэтических клеток и саморегуляция.
Моноцитопоэз
Моноцитопоэз – это стимулирующий фактор колонии фибробластов и фактор роста макрофагов, но он же подавляет (отрицательная саморегуляция) интерферон фибробластов и лейкоцитов и т. д.
Процесс фагоцитоза делится на четыре стадии:
- Приближение.
- Адгезия.
- Оседание;
- Стадия пищеварения.
Фаза приближения
Фагоцит приближается к объекту – бактериям, мервому клеточному элементу, инородному объекту. При движении под действием хемотаксиса цитоплазма фагоцита образует удлинения (псевдоподии).
Адгезия
Способствует образованию аминополисахаридов на поверхности фагоцитов и перекрытию фагоцитарного объекта белками сыворотки, особенно иммуноглобулинами. Последний механизм по сути является опсонизацией – бактерии и поврежденные клетки перекрываются с IgM, IgG и компонентами комплемента (C3, C5 и др.), что облегчает адгезию к фагоциту.

Адгезия
Поверхность фагоцита заряжена отрицательно, поэтому адгезия лучше, если объект, подлежащий фагоцитозу, заряжен положительно. Менее фагоцитарны отрицательно заряженные объекты, такие как опухолевые клетки.
Оседание
Этап фагоцитации объекта – путь инвагинации. Сначала фагоцит образует углубление, а затем фагосому – вакуоль, содержащую объект, подлежащий фагоцитозу. НАДН-зависимая оксидаза в мембране фагоцитов активируется до образования фагосом; в результате O2 превращается в O 2 ~ (супероксид-анион) и образуется H 2 O 2.
Эти продукты обладают бактерицидным действием, а также вызывают образование свободных радикалов. Под действием пероксидаз и каталаз H 2 O 2 расщепляется и высвобождается молекулярный O 2. Свободные радикалы и активный молекулярный O 2 действуют на мембрану фагоцита и объект, подлежащий фагоцитозу, активируя перекисное окисление липидов.
Липопероксиды и свободные радикалы неустойчивы к лизосомным мембранам и способствуют высвобождению лизосомальных ферментов.
Стадия пищеварения
Лизосомы присоединяются к фагосоме, содержащей фагоцитарный объект. Эти органеллы содержат все ферменты, необходимые для расщепления углеводов, белков, жиров и нуклеиновых кислот. В еще неактивной форме они попадают в вакуоль фагоцитов.
Пищеварительная вакуоль образуется при pH около 5,0, близком к оптимальному для лизосомальных ферментов. Активируются лизосомальные ферменты, и фагоцитарный объект постепенно переваривается. Во время стадии пищеварения проницаемость мембраны фагосомы увеличивается, содержимое фагосомы ускользает в цитоплазму, и микрофаг умирает (этому процессу способствует ацидоз). В этом случае фагоцитарный микроорганизм также может сохранять жизнеспособность.
Роль макрофагов в воспалительном процессе
Макрофаги начинают участвовать в фагоцитозе позже микрофагов. Макрофаги также более устойчивы к гипоксии и ацидозу, например, моноциты жизнеспособны даже при pH 5,5.
В очаге воспаления макрофаги выполняют несколько функций:
- Фагоцитируют бактериальные остатки, оставшиеся после эвакуации гноя и продуктов распада тканей – очищающая функция;
- Высвобождают лизосомальные ферменты – гиалуронидазу, аминопептидазу и др;
- Синтезируют компоненты системы комплемента и простагландины.
В то же время воспаленная тканевая среда также стимулирует образование фибробластов и фиброцитов. Постепенно появляется новая, богатая сосудами грануляционная ткань.
Фагоцитоз стимулируется продуктами повреждения тканей – внутриклеточные белки, ферменты, полипептиды, аминокислоты, электролиты и т. д., биологически активными веществами, половыми гормонами, тироксином, адреналином, лихорадкой. Но подавляется недостатком стимулирующих факторов, глюкокортикоидов, гликолортикоидов, ацетилхолина, ацетилхолина.

Фагоцитоз
Считается, что клетки плаценты и злокачественные опухоли способны секретировать вещество, которое подавляет функцию макрофагов, иммунологические реакции лимфоцитов и эмиграцию лейкоцитов, что приводит к значительному ослаблению или даже прекращению воспаления.
Объекты, которые фагоциты не могут переваривать, остаются в этих клетках в течение длительного времени и покрываются тонкой пленкой аминополисахаридов. После гибели фагоцитов они повторно фагоцитируются или выводятся из организма. Процесс, при котором фагоцит после переваривания высвобождает часть продуктов своего фагоцитоза в окружающую среду, называется экструзией.
Фагоцитоз – не единственный в организме механизм борьбы с воспалением. Большинство микроорганизмов погибают в условиях ацидоза, а также от ферментов, высвобождаемых во время гибели клеток и функционирования иммунокомпетентной системы.
Нейтрофилы во внеклеточном пространстве секретируют катионные белки, которые могут убивать ферменты без ферментов и фагоцитоза. Таким образом, воспалительные очаги постепенно избавляются от микроорганизмов и мертвых клеток.
Асептическое и острое воспаление
Дальнейшее течение воспаления зависит от того, является ли воспаление асептическим или бактериальным.
В асептических воспалительных условиях, например, вокруг хирургического шва, инородного тела, стенка микрофагов (нейтрофилов) начинает формироваться в течение нескольких часов, достигая максимума в течение дня.
Позже появляется следующий вал макрофагов, который достигает максимума через 2-3 раза. в день. Эмигрировавшие лейкоциты постепенно становятся неподвижными, больше не могут делиться и погибают в течение 3-5 дней. 2-3. на 5 сутки начинает формироваться стенка фибробластов, а на 5 сутки – соединительнотканная капсула.
Таким образом, в случае острого воспаления патогенный агент в организме преобразует белки, которые участвуют в реакции антиген-антитело, медиаторы и модуляторы воспаления, систему фагоцитов и миграцию клеток. В результате заканчивается острое воспаление. Однако, если инфекционные агенты попадают в участок асептического воспаления, например, в результате травмы, это воспаление становится септическим (бактериальным) воспалением.
Бактериальные воспалительные состояния бактерий и токсинов, не вызывающие гиперемии, экссудации и эмиграции лейкоцитов. Между клетками воспаленных тканей и особенно вокруг кровеносных сосудов накапливается все больше и больше микрофагов. Лизосомы микрофагов содержат множество активных ферментов, и эти клетки также начинают процесс фагоцитоза. При пальпации воспаленная ткань в это время кажется плотной, поэтому это называется стадией воспалительной инфильтрации.
По мере прогрессирования воспаления лейкоциты и те тканевые клетки, которые претерпели необратимые изменения во время воспаления, погибают. В этих клетках высвобождаются лизосомальные ферменты, которые расщепляют тканевые белки, белковые и липидные комплексы и другие структуры. Это стадия гнойного размягчения воспаления.
Эти стадии наблюдаются, например, у пациентов с гнойным воспалением перикарда (фурункул), гнойным воспалением апокринных потовых желез (гидраденит) и гнойным воспалением соединительной ткани (флегмона). При вдыхании воспаленная ткань выглядит мягкой, с характерным раскачиванием – флюктуацией. Образуется замкнутое скопление гноя – абсцесс.

Гнойное воспаление перикарда
Гной разрывается в направлении наименьшего сопротивления (либо наружу, либо внутри тела). Если гной попадает в кровоток (пемия), в организме может развиться множество очагов гноя, но под действием микроорганизмов и токсических веществ – угрожающее общее заболевание (сепсис, септикопиемия). Поэтому важно диагностировать накопление гноя и обеспечить хирургический дренаж гноя наружу.
Читайте также: