
Регуляция пола у растений реферат
Обновлено: 07.07.2024
а) Эмбриогенез - процесс формирования зародыша внутри материнского растения из оплодотворенной яйцеклетки (зиготы) за счет асимметричного деления и дифференцировки клеток. Эмбриогенез сопровождается формированием тканей и органов будущего растительного организма и завершается формированием семени. Все процессы эмбриогенеза у покрытосеменных растений происходят в семязачатке, внутри которого развивается женский гаметофит, происходят оплодотворение, формирование зародыша и семени.
Эмбриогенез может быть разделен на два этапа. На первом этапе формируется зародыш (эмбрион) — задается общий план строения тела растительного организма, создаются оси симметрии, формируются апикальные меристемы и ряд других тканей. Второй этап эмбриогенеза (постэмбриональное развитие) связан с формированием семени: дальнейшей дифференцировкой клеток и тканей, их обезвоживанием и синтезом биополимеров (липидов, крахмала и белков), необходимых в период прорастания и роста проростков.
Формирование зародыша может происходить 2мя путями:
1) Типичный половой процесс: две сливающиеся гаметы формируетзиготический зародышàсемя àформирование плода
2) Апомиксис: зародыш формируется не из зиготы, а из неоплодотворенной яйцеклетки, которая может быть гаплоидной или из-за аномалий гаметогенеза остается диплоидной. Поэтому при апомиксисе могут формироваться как гаплоидные, так и диплоидные зародыши. В случае апомиксиса зародыш может развиваться и из других гаплоидных или диплоидных клеток, ассоциированных с яйцеклеткой
Регуляция: у цветковых растений выделяют две группы генов, контролирующих эмбриогенез:
· гены, которые отвечают за формирование зародыша в целом;
· гены, которые контролируют программы дифференцировки и образования будущих органов растения.
б) Явление покоя очень важно для растений, поскольку позволяет им переждать неблагоприятные условия среды. Различают покой физиологический и вынужденный. Причиной вынужденного покоя являются факторы внешней среды, которые препятствуют прорастанию семян. Физиологический покой зависит от физиологического состояния растения и определяется соотношением гормонов ингибиторов и активаторов. Переход растений от активного роста к физиологическому покою определяется балансом эндогенных ингибиторов (АБК) и активаторов роста (гиббереллины, цитокинины). Переход в состояние покоя семян сопровождается значительным увеличением содержания АБК. Выход из этого состояния связан с уменьшением содержания АБК и повышением уровня гиббереллинов и (или) цитокининов.
2. Вегетативный этап:
включает прорастание семени или органов вегетативного размножения (клубней, луковиц) и формирование вегетативных органов (листьев, стеблей, корней). На этом этапе онтогенеза происходит новообразование клеток, тканей и органов, их активный рост и быстрое увеличение в размерах. Вегетативный период иногда делят на два этапа: прорастания семени; ювенильную стадию.
Прорастание семян начинается при благоприятных условиях увлажнения, температуры и освещения. Этот процесс начинается с набухания семени и заканчивается проклевыванием зародышевого корешка сквозь семенную оболочку. Выход из покоя и инициация прорастания семян обусловлены увеличением уровня оводненности осевых частей зародыша и изменением баланса фитогормонов, в первую очередь таких как АБК, гиббереллины и ауксины.
Ювенильный этап развития включает период от первого листа до заложения цветка и связан с накоплением вегетативной массы растения. В этот период развития растение не способно к цветению (т. е. ювенильно) и размножению.
Развитие апикальных меристем àразвитие листа и корняàдифференциация сосудов
3. Генеративный этап:
Генеративный этап развития растения охватывает период от закладки органов размножения до образования гаметофитов, оплодотворения и образования зиготы. К этому моменту онтогенеза растительного организма накапливается вегетативная масса, достаточная для формирования цветков, семян и плодов.
Органом полового размножения у покрытосеменных растений является цветок. Растения начинают формировать генеративные органы и приобретают способность к размножению при определенных условиях внешней среды после того, как накопят необходимую вегетативную массу. После инициации генеративного этапа развития в апикальных меристемах вместо примордиев листьев начинают закладываться цветочные почки, содержащие зачатки органов цветка.
Факторы инициации цветения делятся на эндогенные (внутренние) и экзогенные (внешние). Внешними факторами, индуцирующими переход к цветению, являются фотопериод (относительная продолжительность светлого и темного времени суток) и температура. К эндогенным факторам цветения относятся возраст растения, вегетативная масса, содержание фитогормонов (в первую очередь гиббереллинов) и сахаров.
Растения, у которых цветение индуцируется "коротким днем", называют короткодневными. В том случае, когда зацветанию благоприятствуют "длинные дни", растения называют длиннодневными. Виды растений, у которых "длина дня" не оказывает существенного влияния на цветение, называют нейтральными.
Установлено так же, что гиббереллины способны стимулировать превращение генеративной меристемы в цветковую. Гиббереллин-зависимый путь инициации цветения становится основным в условиях короткого дня. Гиббереллин индуцирует цветение, активируя транскрипцию гена LFY, который является ключевым на втором этапе развития цветка — на этапе детерминации флоральной меристемы. Завершается процесс инициации цветения эвокацией — необратимыми процессами, происходящими в апикальной меристеме побега, которые приводят к формированию репродуктивных органов растения.
Последующие этапы генеративного развития растения включают: формирование цветковых меристем; формирование органов цветка; гаметогенез.
4. Сенильный этап:
Последний этап развития растения — сенильный (этап старости и отмирания) — включает период от полного прекращения плодоношения до естественного отмирания растений. Рост в этом случае обнаруживается очень редко.Старение выражается в прогрессирующем нарушении синтеза макромолекул и систем регуляции организма, накоплении токсичных и инертных в химическом отношении продуктов, постепенном угасании отдельных физиологических функций. Старение представляет собой серию упорядоченных цитологических и биохимических событий. Нарушается функционирование одних органелл (хлоропласты), в то время как другие (ядро) остаются структурно и функционально активными до самых последних стадий старения. В стареющих тканях идут процессы катаболизма, которые требуют синтеза denovo гидролитических ферментов.Существование и развитие растительного организма связано с постоянной гибелью части составляющих его клеток. Программированная гибель клеток (у животных этот процесс называют апоптозом) играет важную роль в процессах морфогенеза и при повреждениях. За счет отмирания отдельных тканей и клеток в растении формируются сосуды флоэмы и ксилемы, защитные структуры типа корневого чехлика, некротические пятна при поражении патогеном.
Определение пола у растений:
Под определением (детерминацией) пола у растений понимается формирование признаков пола у клеток, органов или особей под воздействием, как генетических факторов, так и условий внешней и внутренней среды. По наличию и степени развития генеративных органов цветки делят на обоеполые (гермафродитные) и однополые (раздельнополые). Последние бывают пестичными (женскими) или тычиночными (мужскими).
Установлено, что гены, ответственные за детерминацию пола у растений располагаются не только в половых хромосомах, но и в других хромосомах (аутосомах). Под действием различных причин может происходить даже полное превращение одного пола в другой. Это связано с регуляторными процессами, обусловленными действием факторов внешней среды, и внутренними изменениями метаболического характера.
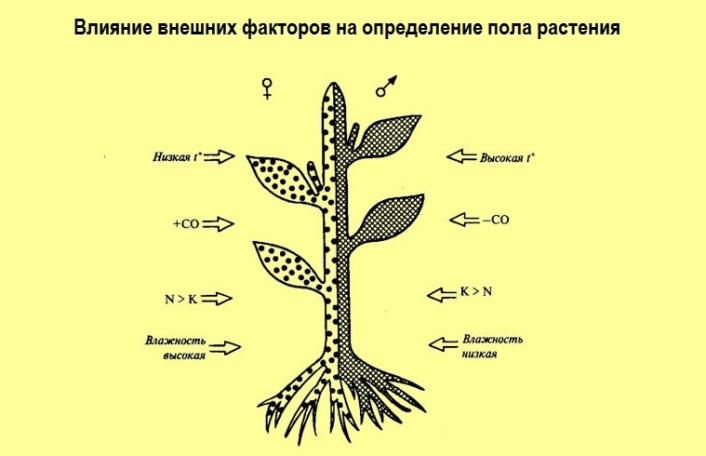
Появлению женских цветков и формированию женских растений у двудомных видов (женская сексуализация) способствуют низкие температуры, высокая влажность, хорошее азотное питание. Дифференцировке мужских цветков (мужская сексуализация) — высокие температуры, низкая влажность, калийное питание. Показано, что короткий день приводит к появлению большего количества женских особей КДР хмеля и конопли. Длинный свет действует противоположным образом. Красный (длинноволновой) свет усиливает проявление мужских признаков, в синий (коротковолновой) — женских. Экспериментальные данные дают основание полагать, что действие спектрального состава на проявление пола у растений осуществляется через фитохромную систему и связано с содержанием и активностью фитогормонов.Исследования, проведенные на двудомных растениях (конопля, шпинат) и на однодомных с раздельнополыми цветками (кукуруза, огурцы) показали роль гиббереллинов в формировании признаков мужского пола, а цитокининов, ауксинов и этилена — женских признаков. Уровень фитогормонов в растении определяется не только действием внешней среды, но и коррелятивными взаимосвязями между органами. В этой связи были проведены эксперименты по влиянию удаления части побегов или обрезки корней на формирование признаков пола. Удаление корней у конопли и шпината способствовало увеличению числа мужских растений. Введение в среду выращивания таких растений цитокинина приводило к образованию женских цветков. Удаление листьев способствовало возрастанию количества женских растений, а введение гиббереллинов увеличивало число мужских экземпляров. Предполагают, что цитокинины, образующиеся в корнях, передвигаясь в апекс, включают программу, обусловливающую проявление женского пола (пестичные цветки), тогда как гиббереллины, образующиеся в листьях, включают программу, связанную с мужской сексуализацией (тычиночные цветки).
Вопрос о формировании пола у растений относится к самым сложным и наименее изученным проблемам биологии развития растений. Вместе с тем, его изучение имеет непосредственное значение для разработки приемов повышения урожайности многих сельскохозяйственных культур, особенно у тех, которые образуют раздельнополые цветки.

Определение пола у растений
Под определением пола у растений понимается формирование признаков пола у клеток, органов или особей под воздействием как генетических факторов, так и условий внешней и внутренней среды. По наличию и степени развития генеративных органов цветки делят на обоеполые (гермафродитные) и однополые (раздельнополые). Последние бывают пестичными (женскими) или тычиночными (мужскими). Если на одном растении находятся цветки разных типов, то такие растения называют однодомными. Растения, имеющие на одних экземплярах пестичные цветки, а на других — тычиночные, называются двудомными. Интересно отметить, что только 4-5% видов растений двудомны. Представителями двудомных растений являются крапивные, хмель, облепиха, тополь, ива, щавель кислый, шпинат и др.
Процесс формирования пола происходит у растений в разные периоды его развития и определяется в основном влиянием внешних условий. Дело в том, что в отличие от животных организмов процесс формирования пола у растений подвержен большим колебаниям. Под действием различных причин может происходить даже полное превращение одного пола в другой. Это связано с регуляторными процессами, обусловленными действием факторов внешней среды, и внутренними изменениями метаболического характера.
Влияние внешних факторов на определение пола у растений
Многочисленные факты свидетельствуют о влиянии на проявление признаков пола у растений таких внешних воздействий, как влажность почвы и воздуха, температура, спектральный состав света, условия минерального питания. Так, появлению женских цветков и формированию женских растений у двудомных видов способствуют низкие температуры, высокая влажность, хорошее азотное питание. Появлению мужских цветков — высокие температуры, низкая влажность, калийное питание. Определение пола зависит также от длины дня. Доказано, что короткий день приводит к появлению большего количества женских особей. Длинный световой день действует противоположным образом.
Влияние фитогормонов на определение пола у растений
Исследования, проведенные на двудомных растениях (крапивные) и на однодомных с раздельнополыми цветками (кукуруза, огурцы) показали роль фитогормонов в формировании пола растения. Основной парой фитогормонов, контролирующих проявление пола у растений, являются гиббереллины и цитокинины. Гиббереллины принимают участие в формировании признаков мужского пола, а цитокинины и ауксины — женских признаков.
Уровень фитогормонов в растении определяется не только действием внешней среды, но и коррелятивными взаимосвязями между органами. Были проведены эксперименты по влиянию удаления части побегов или обрезки корней на формирование признаков пола. Удаление корней у крапивных способствовало увеличению числа мужских растений. Введение цитокинина в среду выращивания этих же растений приводило к образованию женских цветков. Предполагают, что цитокинины, образующиеся в корнях, передвигаясь в апекс (прим.: верхушка побега), включают программу, обусловливающую проявление женского пола, тогда как гиббереллины, образующиеся в листьях, включают программу, связанную с мужской сексуализацией. Однако если половая дифференцировка клеток уже началась, то воздействие фитогормонами не приведет к изменению направленности в формировании пола, предопределенного генетически. Развитие растений осуществляется по определенной генетической программе. Развертывание этой программы, в том числе проявление пола, старение и смерть организма, может в определенной мере контролироваться внешними условиями, через внутреннюю гормональную систему регуляции. Таким образом, развитие растений происходит при взаимодействии генетических потенций и факторов внешней среды.
Опыление
Процесс переноса пыльцы с пыльника (мужской цветок) на рыльце пестика (женский цветок) называется опылением.
Однодомные растения имеют и мужские и женские цветки на одном растении. Например, кукуруза – мужские цветки собраны на верхушке метелкой, а женские — на стволе початками. Опыление у кукурузы происходит при помощи ветра.
Томаты, баклажаны и многие зерновые культуры относятся к самоопыляемым растениям. Например, цветки томата имеют и пестики, и тычинки. Тычинки срослись так, что в большинстве случаев пестик оплодотворяется собственной пыльцой (факультативное самоопыление).
Двудомные растения устроены так, что на мужских растениях имеются только мужские цветки с тычинками и пыльцой, а на женских – женские цветки с пестиками. Если перенос пыльцы осуществляется между цветками разных особей, такое опыление называется перекрестным. Перекрестное опыление обусловливает обмен генами, поддерживает высокий уровень генетического разнообразия популяций и создает широкое поле для естественного отбора. Для перекрестного опыления, как правило, требуется участие посредника – насекомого, животного, человека. Поэтому выращивая двудомные растения в условиях закрытого грунта, придется процесс опыления взять на себя.

Искусственное опыление растений в условиях закрытого грунта
Шаг 1. Сбор пыльцы
Если вы разводите растения для себя, в небольших масштабах, то для получения семян будет достаточно пыльцы с одной веточки мужского растения. Во избежание случайного или преждевременного опыления изолируйте мужское растение как можно скорее при появлении на нем пыльника.
Перед сбором пыльцы наденьте на ветку чистый пакет (бумажный или целлофановый). Перевяжите пакет так, чтобы он плотно прилегал к ветке, и пыльца не выпадала из него. Оставьте пакет в таком состоянии на нескольких дней. Перед снятием пакета, когда пыльцы будет собрано достаточно, легонько потрясите веточку, тогда все остатки пыльцы опадут в пакет. Аккуратно срежьте ветку и только после этого снимите пакет.
Шаг 2. Хранение пыльцы
Если опыление женских растений будет производиться не сразу, то пыльцу надо умело сохранить, т. к. в естественных условиях она хранится недолго. Ее легко уничтожить высокой температурой и влажностью. Но в морозилке пыльца может храниться несколько месяцев. Поэтому собранную пыльцу аккуратно достаем из пакета и высыпаем на бумагу. Для удаления растительных остатков можно просеять пыльцу через сито (под сито также кладется бумага). Затем предварительно стерилизованным скребком собираем пыльцу с бумаги, помещаем ее в стерильную емкость и убираем в морозилку.

Шаг 3. Процесс опыления
Если процесс опыления производится сразу после сбора пыльцы, то можно просто надеть пакет с пыльцой на женское растение и слегка потрясти. Для надежности результата можно оставить пакет на женском растении на сутки. Убедитесь, что при опылении нет сквозняков, и пыльца не рассыпалась. Если в оранжерее есть растения, не предназначенные для опыления – изолируйте их, либо проводите опыление в отдельном месте.
ВАЖНО: перед каждой новой процедурой опыления делайте уборку там, где опыляете. Перед тем, как относить женские растения после оплодотворения обратно в оранжерею, обрызгайте их водичкой, чтобы смыть и уничтожить остатки пыльцы. Это позволит предотвратить опыление остальных растений.
Если процесс опыление производится спустя какое-то время и пыльца хранилась в морозилке, то опылять придется вручную кисточкой. Для этого окунаем кисть в контейнер с пыльцой и аккуратно и легко на¬носим кистью пыльцу на пестики. Здесь следует быть внимательным: пыльца не должна случайно стряхнуться с кисточки. Данный способ всем хорош, однако семян в итоге получается не так много, как в первом случае.
Несомненно, закрытый грунт имеет свои особенности и некоторые сложности воспроизводства условий окружающей среды для успешного развития растений. Но
современный уровень науки и знаний позволяют получать отличные урожаи и даже вывести потомство в виде семян.
Вопрос о формировании пола у растений относится к самым сложным и наименее изученным проблемам биологии развития растений. Вместе с тем, его изучение имеет непосредственное значение для разработки приемов повышения урожайности многих сельскохозяйственных культур, особенно у тех, которые образуют раздельнополые цветки.
Определение пола у растений
Под определением пола у растений понимается формирование признаков пола у клеток, органов или особей под воздействием как генетических факторов, так и условий внешней и внутренней среды. По наличию и степени развития генеративных органов цветки делят на обоеполые (гермафродитные) и однополые (раздельнополые). Последние бывают пестичными (женскими) или тычиночными (мужскими). Если на одном растении находятся цветки разных типов, то такие растения называют однодомными. Растения, имеющие на одних экземплярах пестичные цветки, а на других — тычиночные, называются двудомными. Интересно отметить, что только 4-5% видов растений двудомны. Представителями двудомных растений являются крапивные, хмель, облепиха, тополь, ива, щавель кислый, шпинат и др.
Процесс формирования пола происходит у растений в разные периоды его развития и определяется в основном влиянием внешних условий. Дело в том, что в отличие от животных организмов процесс формирования пола у растений подвержен большим колебаниям. Под действием различных причин может происходить даже полное превращение одного пола в другой. Это связано с регуляторными процессами, обусловленными действием факторов внешней среды, и внутренними изменениями метаболического характера.
Влияние внешних факторов на определение пола у растений
Многочисленные факты свидетельствуют о влиянии на проявление признаков пола у растений таких внешних воздействий, как влажность почвы и воздуха, температура, спектральный состав света, условия минерального питания. Так, появлению женских цветков и формированию женских растений у двудомных видов способствуют низкие температуры, высокая влажность, хорошее азотное питание. Появлению мужских цветков — высокие температуры, низкая влажность, калийное питание. Определение пола зависит также от длины дня. Доказано, что короткий день приводит к появлению большего количества женских особей. Длинный световой день действует противоположным образом.
Влияние фитогормонов на определение пола у растений
Исследования, проведенные на двудомных растениях (крапивные) и на однодомных с раздельнополыми цветками (кукуруза, огурцы) показали роль фитогормонов в формировании пола растения. Основной парой фитогормонов, контролирующих проявление пола у растений, являются гиббереллины и цитокинины. Гиббереллины принимают участие в формировании признаков мужского пола, а цитокинины и ауксины — женских признаков.
Уровень фитогормонов в растении определяется не только действием внешней среды, но и коррелятивными взаимосвязями между органами. Были проведены эксперименты по влиянию удаления части побегов или обрезки корней на формирование признаков пола. Удаление корней у крапивных способствовало увеличению числа мужских растений. Введение цитокинина в среду выращивания этих же растений приводило к образованию женских цветков. Предполагают, что цитокинины, образующиеся в корнях, передвигаясь в апекс (прим.: верхушка побега), включают программу, обусловливающую проявление женского пола, тогда как гиббереллины, образующиеся в листьях, включают программу, связанную с мужской сексуализацией. Однако если половая дифференцировка клеток уже началась, то воздействие фитогормонами не приведет к изменению направленности в формировании пола, предопределенного генетически. Развитие растений осуществляется по определенной генетической программе. Развертывание этой программы, в том числе проявление пола, старение и смерть организма, может в определенной мере контролироваться внешними условиями, через внутреннюю гормональную систему регуляции. Таким образом, развитие растений происходит при взаимодействии генетических потенций и факторов внешней среды.
Опыление
Процесс переноса пыльцы с пыльника (мужской цветок) на рыльце пестика (женский цветок) называется опылением.
Однодомные растения имеют и мужские и женские цветки на одном растении. Например, кукуруза – мужские цветки собраны на верхушке метелкой, а женские — на стволе початками. Опыление у кукурузы происходит при помощи ветра.
Томаты, баклажаны и многие зерновые культуры относятся к самоопыляемым растениям. Например, цветки томата имеют и пестики, и тычинки. Тычинки срослись так, что в большинстве случаев пестик оплодотворяется собственной пыльцой (факультативное самоопыление).
Двудомные растения устроены так, что на мужских растениях имеются только мужские цветки с тычинками и пыльцой, а на женских – женские цветки с пестиками. Если перенос пыльцы осуществляется между цветками разных особей, такое опыление называется перекрестным. Перекрестное опыление обусловливает обмен генами, поддерживает высокий уровень генетического разнообразия популяций и создает широкое поле для естественного отбора. Для перекрестного опыления, как правило, требуется участие посредника – насекомого, животного, человека. Поэтому выращивая двудомные растения в условиях закрытого грунта, придется процесс опыления взять на себя.
Искусственное опыление растений в условиях закрытого грунта
Шаг 1. Сбор пыльцы
Если вы разводите растения для себя, в небольших масштабах, то для получения семян будет достаточно пыльцы с одной веточки мужского растения. Во избежание случайного или преждевременного опыления изолируйте мужское растение как можно скорее при появлении на нем пыльника.
Перед сбором пыльцы наденьте на ветку чистый пакет (бумажный или целлофановый). Перевяжите пакет так, чтобы он плотно прилегал к ветке, и пыльца не выпадала из него. Оставьте пакет в таком состоянии на нескольких дней. Перед снятием пакета, когда пыльцы будет собрано достаточно, легонько потрясите веточку, тогда все остатки пыльцы опадут в пакет. Аккуратно срежьте ветку и только после этого снимите пакет.
Шаг 2. Хранение пыльцы
Если опыление женских растений будет производиться не сразу, то пыльцу надо умело сохранить, т. к. в естественных условиях она хранится недолго. Ее легко уничтожить высокой температурой и влажностью. Но в морозилке пыльца может храниться несколько месяцев. Поэтому собранную пыльцу аккуратно достаем из пакета и высыпаем на бумагу. Для удаления растительных остатков можно просеять пыльцу через сито (под сито также кладется бумага). Затем предварительно стерилизованным скребком собираем пыльцу с бумаги, помещаем ее в стерильную емкость и убираем в морозилку.
Шаг 3. Процесс опыления
Если процесс опыления производится сразу после сбора пыльцы, то можно просто надеть пакет с пыльцой на женское растение и слегка потрясти. Для надежности результата можно оставить пакет на женском растении на сутки. Убедитесь, что при опылении нет сквозняков, и пыльца не рассыпалась. Если в оранжерее есть растения, не предназначенные для опыления – изолируйте их, либо проводите опыление в отдельном месте.
ВАЖНО: перед каждой новой процедурой опыления делайте уборку там, где опыляете. Перед тем, как относить женские растения после оплодотворения обратно в оранжерею, обрызгайте их водичкой, чтобы смыть и уничтожить остатки пыльцы. Это позволит предотвратить опыление остальных растений.
Если процесс опыление производится спустя какое-то время и пыльца хранилась в морозилке, то опылять придется вручную кисточкой. Для этого окунаем кисть в контейнер с пыльцой и аккуратно и легко на¬носим кистью пыльцу на пестики. Здесь следует быть внимательным: пыльца не должна случайно стряхнуться с кисточки. Данный способ всем хорош, однако семян в итоге получается не так много, как в первом случае.
Несомненно, закрытый грунт имеет свои особенности и некоторые сложности воспроизводства условий окружающей среды для успешного развития растений. Но
современный уровень науки и знаний позволяют получать отличные урожаи и даже вывести потомство в виде семян.
Проблема регуляции пола вытекает из необходимости увеличения продукции животноводства за счет преимущественного получения особей одного вида, дающих более высокий выход молока, мяса, шерсти, яиц и т. д. Так, в молочном скотоводстве более желательно рождение телочек, а в мясном — бычков, так как они быстрее растут. От высокоценных племенных быков и коров целесообразно получать мужских потомков для более быстрого размножения их генотипов. В яичном птицеводстве экономически более выгодно получение курочек. В связи с этими практическими потребностями исследователи не только стремятся познать механизмы определения пола, но и изучают возможности искусственного регулирования пола. Необходимо отметить, что в отношении крупных животных с внутриутробным развитием плодов эта проблема еще не решена. Регуляция соотношения полов у млекопитающих может быть достигнута путем разделения спермы на две фракции: первую — содержащую в спермиях Х-хромосому и вторую — содержащую Y-хромосому. Оплодотворение самок одной из этих фракций будет давать приплод одного пола. Проводились эксперименты по разделению спермы на указанные фракции центрифугированием, электрофорезом и седиментацией (осаждением) с помощью аминокислого гистидина. Осеменение самок крольчих, например, более легкой и более подвижной фракцией приводило к сдвигу в сторону мужского пола. Однако полного сдвига в соотношении полов сделано не было. Разрабатывается метод количественного определения ДНК в спермиях путем измерения интенсивности флуоресценции ядер. Полученные результаты, как считают авторы этого метода, могут стать предпосылкой для успешного разделения спермиев у млекопитающих на несущие X-или Y-хромосому.
Партеногенез. Это развитие организма без оплодотворения. Получение особей одного пола может быть достигнуто при развитии эмбрионов из отцовских (андрогенез) или материнских (гиногенез) гамет. Так, под руководством Б. Л. Астаурова были проведены эксперименты по андрогенезу у тутового шелкопряда. Неоплодотворенные яйца шелкопряда подвергали тепловому шоку и облучали рентгеном, тем самым разрушали их ядра, не повредив цитоплазму. Затем эти яйца осеменяли. Зигота формировалась путем слияния ядер двух проникших в яйцо спермиев, развившиеся из нее особи имели признаки только отцовского вида.
В другом варианте экспериментов неоплодотворенные и еще не прошедшие редукционного деления яйца нагревали, останавливая тем самым мейоз и сохраняя диплоидность набора хромосом. Из таких яиц без оплодотворения (партеногенетически) развивались только самки, унаследовавшие признаки матери. Девственное развитие (партеногенез) в естественных или спонтанных условиях встречается у птиц. И. В. Кудрявцев, 3. А. Ощепкова, А. К. Голубев и др. на основании экспериментов, проведенных на курах разных пород, пришли к выводу, что существуют генетические предпосылки селекции на получение жизнеспособных особей — партеногенов и создание линий с высокой предрасположенностью к партеногенезу. При этом все вылупляющиеся цыплята оказываются петушками. Отбор на повышение частоты партеногенеза, проведенный в двух линиях индеек, дал следующие результаты. Способность яиц к партеногенезу возросла с 1,1 до 18,6 % в первой линии и с 4 до 21,1% во второй. Значение партеногенеза не только в том, что он позволяет получить потомство одного пола, но и в том, что использование этого явления дает возможность получить особей, идентичных генотипу одной из родительских форм.
ЗАКЛЮЧЕНИЕ
Проблема происхождения половых различий, механизмов определения пола животных организмов очень важна и для теоретической биологии, и для практики. Достаточно сказать, что возможность искусственного регулирования пола животных была бы исключительно полезна для сельского хозяйства.
Половое размножение свойственно почти всем организмам. Благодаря скрещиванию и комбинации генов возникают генотипы, способствующие успешному приспособлению индивида к меняющимся условиям внешней среды. Половой процесс обеспечивает широкое распространение внутри вида любого гена, особенно имеющего приспособительное значение.
Пол у животных определяется, как правило, в момент оплодотворения яйцеклетки. В этом случае важнейшая роль в генетическом определении пола принадлежит хромосомному набору зиготы. В диплоидных наборах растений и животных содержатся парные гомологичные хромосомы, одинаковые по форме, с одинаковыми генами. Хромосомы, одинаковые у мужских и женских организмов, называются аутосомами. Но по одной паре хромосом женские и мужские особи различаются. В этой паре хромосомы не одинаковы по форме или размерам и по находящимся в них генам. Хромосомы, по которым мужской и женский пол отличаются друг от друга, называются половыми или гетерохромосомами.
Регуляция пола, имеет важное практическое значение. Так, в яичном птицеводстве желательно получать больше курочек, а в мясном - петушков. В молочном скотоводстве нужны телочки, в мясном - бычки. У тутового шелкопряда самцы дают на 25-30% больше шелка, чем самки, поэтому их преимущество очевидно.
У животных можно получать организмы желательного пола с использованием биотехнологических приемов, таких как трансплантация и клонирование. Ученые в настоящее время разработали способ разделения эмбрионов на части, У домашних животных эмбрион можно разделить на 4, 8 или большее количество частей. При этом одну из частей эмбриона подсаживают в матку самки, а остальные замораживают и сохраняют длительное время в жидком азоте. После рождения потомка можно делать пересадки и остальных частей разделенного эмбриона. В этом случае пол потомков будет такой же, как и у рожденного животного.
Описанное Морганом сцепленное наследование признаков было обнаружено у многих видов животных и человека. У кур сцепленно с полом наследуется поперечно-полосатая окраска оперения. Это явление используется в птицеводстве для разделения по полу цыплят в суточном возрасте. При скрещивании петушков, имеющих сплошную окраску оперения, с поперечно- полосатыми курами в потомстве петушки будут поперечно-полосатыми, а курочки - со сплошной окраской оперения.
Список литературы
Фотопериодическая реакция и действие фитохрома обусловливают только общую подготовленность растения к зацветанию и образованию цветков. Но что же конкретно вызывает переход растений к цветению? На этот счет в разные периоды существовали сменявшие одна другую гипотезы. Последней по времени и принятой в настоящее время является теория гормональной регуляции цветения. Впервые она была разработана М. X. Чайлахяном в 1937 г., а затем получила широкое распространение и всеобщее признание.
Отправным моментом теории считается наличие в растениях специфического гормона цветения — флоригена. Он состоит из двух компонентов. Один из них — гиббереллин — вещество, реально существующее, идентифицированное химически, присутствующее во всех растениях. Второй компонент — антезйн — до сих пор не выделенное и не идентифицированное вещество, и о его присутствии судят только по производимому им эффекту зацветания.
Цветение растения может произойти только при достаточном накоплении этих двух компонентов. У фотопериодически нейтральных растений их образование находится под генным контролем и не зависит от длины дня. Следовательно, у них существует автоматическая регуляция образования флоригена, т. е. его физиологический градиент в растениях. При достижении определенной концентрации фитогормона растение зацветает. У фотопериодически чувствительных видов этот градиент не имеет значения, а цветение растений определяется физиологическим состоянием листьев. Они получают свет определенной продолжительности и качества и только тоща вырабатывают фйтогормоны, точнее, два его компонента. При неблагоприятной длине дня растение вырабатывает только один компонент и к зацветанию перейти не может. Так, длиннодневные растения при коротком дне вырабатывают только антезин и не цветут. Но если в растения ввести дополнительно экзогенный гиббереллин, они бь!стро образуют цветки и зацветут. То же наблюдается и у ко- рстсодневных растений: на длинном дне они образуют гиббереллин, вытягиваются, но не дают цветков. При прививке к ним цветущих побегов растений того же вида они получают дополнительно антезин и нормально зацветают.
Теория гормональной регуляции пола у растений
Большая часть видов цветковых растений имеет обоеполые цветки, однако существует достаточно много видов с однополыми мужскими и женскими цветками. У одних растений эти два рода цветков располагаются на одном и том же экземпляре (однодомные растения), у других — на разных экземплярах (двудомные растения). Соотношение мужских и женских цветков у однодомных или мужских и женских экземпляров у двудомных имеет практическое значение, так как от этого зависит продуктивность растения. Поэтому важно знать, как регулируется это соотношение и как его можно изменить. В настоящее время это объясняют с позиций теории гормональной регуляции пола, которая разработана М. X. Чайлахяном.
Установлено, что многие синтетические аналоги фитогормонов вызывают изменение соотношения мужских и женских растений у таких двудомных видов, как шпинат и конопля. Обработка этих растений гиббереллином обусловливает маскулинизацию (проявление мужского пола), а обработка ауксином, цитокининами, абс- цизовой кислотой — феминизацию (сдвиг в сторону женского пола). Однако в растении присутствуют одновременно все фйтогормоны. Как же происходит регуляция пола в организме? в этом процессе важную роль играют различные органы растения, так как они вырабатывают неодинаковые фйтогормоны. Так, роль корней, синтезирующих в основном цитокинины, заключается в смещении признаков пола в женскую сторону. Роль листьев, вырабатывающих преимущественно гиббереллины, состоит в смещении признаков пола в мужскую сторону. Таким образом, соотношение полов в растениях определяется в основном соотношением цитокининов и гиббереллинов. Участие других фитогормонов — ауксинов и абсцизовой кислоты — проявляется более слабо и в связи с этим недостаточно изучено. Описанные выше закономерности были подтверждены М. X. Чайлахяном экспериментально в очень простых и изящных опытах с различными однодомными и двудомными растениями в водных культурах. Если у растений подрезать корневую систему, то уменьшится синтез цитокининов, вследствие чего гиббереллины будут относительно преобладать. Это изменит равновесие между фитогормонами и сдвинет соотношение в пользу мужского пола (маскулинизация). Если же, напротив, удалить часть листьев, то уменьшится синтез гиббереллинов, а в растениях будут относительно преобладать цитокиничы. Тогда соотношение изменится в сторону преобладания женского пола (феминизация). Полученные данные дают возможность проводить такую регуляцию в производственных условиях. Разумеется, это может быть достигнуто не удалением части листьев и корней, а обработкой растений аналогами фитогормонов. Перспективными для практического использования могут быть различные цитокинины. В настоящее время в селекционно-семеноводческой практике в основном применяют гиббереллин — для стимулирования образования мужских цветков, необходимых для опыления у партено- карпических сортов огурца.
Читайте также: