
Реферат сцепленное наследование генов
Обновлено: 07.07.2024
Кроссинговер происходит в профазе I мейоза во время конъюгации гомологичных хромосом (рис. 3.10). В это время части двух хромосом могут перекрещиваться и обмениваться своими участками. В результате возникают качественно новые хромосомы, содержащие участки (гены) как материнских, так и отцовских хромосом. Особи, которые получаются из таких гамет с новым сочетанием аллелей, получили название… Читать ещё >
Сцепленное наследование. Хромосомная теория наследственности ( реферат , курсовая , диплом , контрольная )
Независимое комбинирование признаков (третий закон Менделя) осуществляется при условии, что гены, определяющие эти признаки, находятся в разных парах гомологичных хромосом. Следовательно, у каждого организма число генов, способных независимо комбинироваться в мейозе, ограничено числом хромосом. Однако в организме число генов значительно превышает количество хромосом.
Например, у кукурузы изучено более 500 генов, у мухи дрозофилы — более 1 тыс., а у человека — около 2 тыс. генов, тогда как хромосом у них 10,4 и 23 пары соответственно. Это дало основание предположить, что в каждой хромосоме локализовано множество генов. Гены, локализованные в одной хромосоме, образуют группу сцепления и наследуются вместе. Число групп сцепления соответствует гаплоидному набору хромосом, поскольку группу сцепления составляют две гомологичные хромосомы, в которых локализованы одинаковые гены. Закономерности сцепленного наследования генов были изучены Т. Х. Морганом и его учениками в начале 20-х годов XX века. Объектом для исследований являлась плодовая мушка дрозофила. У дрозофилы гены, контролирующие окраску тела и длину крыла, представлены следующими парами аллелей: серое тело — черное тело, длинные крылья — зачаточные (короткие). Серое тело и длинные крылья доминируют. Ожидаемое отношение фенотипов в F2 от скрещивания между гомозиготной с серым телом и длинными крыльями и гомозиготной с черным телом и зачаточными крыльями должно составить 9:3:3:1. Это указывало на обычное менделеевское наследование при дигибридном скрещивании, обусловленное случайным распределением генов, находящихся в разных, негомологичных хромосомах. Однако вместо этого в F2 были получены в основном родительские фенотипы в отношении примерно 3:1. Это можно объяснить, предположив, что, гены окраски тела и длины крыла локализованы в одной и той же хромосоме, т. е. сцеплены.
Способ наследования сцепленных генов отличается от наследования генов, локализованных в разных парах гомологичных хромосом. Так, если при независимом комбинировании дигибрид образует четыре типа гамет (АВ, Аb, аВ и аb) в равных количествах, то такой же дигибрид образует только два типа гамет: (АВ и аb) тоже в равных количествах. Последние повторяют комбинацию генов в хромосоме родителя. Было установлено, однако, что кроме обычных гамет возникают и другие — Аb и аВ — с новыми комбинациями генов, отличающимися от родительской гаметы. Причиной возникновения новых гамет является обмен участками гомологичных хромосом, или кроссинговер.
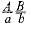
Кроссинговер происходит в профазе I мейоза во время конъюгации гомологичных хромосом (рис. 3.10). В это время части двух хромосом могут перекрещиваться и обмениваться своими участками. В результате возникают качественно новые хромосомы, содержащие участки (гены) как материнских, так и отцовских хромосом. Особи, которые получаются из таких гамет с новым сочетанием аллелей, получили название кроссинговерных или рекомбинантных.
Частота (процент) перекреста между двумя генами, расположенными в одной хромосоме, пропорциональна расстоянию между ними. Кроссинговер между двумя генами происходит тем реже, чем ближе друг к другу они расположены. По мере увеличения расстояния между генами все более возрастает вероятность того, что кроссинговер разведет их по двум разным гомологичным хромосомам (12, "https://referat.bookap.info").
Расстояние между генами характеризует силу их сцепления. Имеются гены с высоким процентом сцепления и такие, где сцепление почти не обнаруживается. Однако при сцепленном наследовании максимальная величина кроссинговера не превышает 50%. Если же она выше, то наблюдается свободное комбинирование между парами аллелей, не отличимое от независимого наследования.
Схема кроссинговера: I — отсутствие кроссинговера; 2 — кроссинговер на стадии двух хромосом; 3 — кроссинговер на стадии четырех хроматид Практически, однако, соотношение 3:1 никогда не наблюдается, а возникают все четыре фенотипа. Это объясняется тем, что полное сцепление встречается редко. В большинстве экспериментов по скрещиванию при наличии сцепления помимо мух с родительскими фенотипами обнаруживаются особи с новыми сочетаниями признаков.
Эти новые фенотипы называют рекомбинантными. Все это позволяет дать следующее определение сцепления: два или более генов называют сцепленными, если потомки с новыми генными комбинациями (рекомбинантны) встречаются реже, чем родительские фенотипы.
Основные положения хромосомной теории наследственности. Генетические механизмы формирования пола. Основные группы сцепления и карты хромосом у человека. Понятие о кроссинговере, его биологическое значение. Наследование признаков, контролируемых полом.
| Рубрика | Биология и естествознание |
| Вид | курсовая работа |
| Язык | русский |
| Дата добавления | 10.03.2019 |
| Размер файла | 291,7 K |
Студенты, аспиранты, молодые ученые, использующие базу знаний в своей учебе и работе, будут вам очень благодарны.
по учебной дисциплине / междисциплинарному курсу
Тема: Генетика пола, сцепленное наследование, кроссинговер
Актуальность работы. Генетика является одной из самых прогрессивных наук естествознания. Ее достижения изменили естественнонаучное и во многом философское понимание явлений жизни. Роль генетики для практики селекции и медицины очень велика. Значение генетики для медицины будет возрастать с каждым годом, ибо генетика касается самых сокровенных сторон биологии и физиологии человека. Благодаря генетике, ее знаниям, разрабатываются методы лечения ряда наследственных заболеваний, таких, как фенилкетонурия, сахарный диабет и другие. Здесь медико-генетическая работа призвана облегчить страдания людей от действия дефектных генов, полученных ими от родителей. Внедряются в практику приемы медико-генетического консультирования и пренатальной диагностики, что позволяет предупредить развитие наследственных заболеваний.
Цель и задачи исследования: Изучить генетику пола и наследование признаков, сцепленных с полом, рассмотреть сцепление генов и процесс кроссинговера. Для достижения этой цели были поставлены следующие задачи:
1. Изучить закономерности наследования пола и генов, локализованных в половых хромосомах;
2. Изучить закономерности наследования сцепленных генов;
3. Дать понятие о кроссинговере и охарактеризовать основные группы сцепления у человека;
Глава 1. Генетика пола
Пол - совокупность признаков, по которым производится специфическое разделение особей или клеток, основанное на морфологических и физиологических особенностях, позволяющее осуществлять в процессе полового размножения комбинирование в потомках наследственных задатков родителей.
Морфологические и физиологические признаки, по которым производится специфическое разделение особей, называется половым (Курчанов, Н. А., 2009).
Признаки, связанные с формированием и функционированием половых клеток, называется первичными половыми признаками. Это гонады (яичники или семенники), их выводные протоки, добавочные железы полового аппарата, копулятивные органы. Все другие признаки, по которым один пол отличается од другого, получили название вторичных половых признаков. К ним относят: характер волосяного покрова, наличие и развитие молочных желез, строение скелета, тип развития подкожной жировой клетчатки, строение трубчатых костей и др.
хромосомный наследственность генетический кроссинговер
1.1 Генетические механизмы формирования пола
Начало изучению генотипического определения пола было положено открытием американскими цитологами у насекомых различия в форме, а иногда и в числе хромосом у особей разного пола (Мак-Кланг, 1906, Уилсон, 1906) и классическими опытами немецкого генетика Корренса по скрещиванию однодомного и двудомного видов брионии. Уилсон обнаружил, что у клопа Lydaeus turucus самки имеют 7 пар хромосом, у самцов же 6 пар одинаковых с самкой хромосом, а в седьмой паре одна хромосома такая же, как соответствующая хромосома самки, а другая маленькая.
Пара хромосом, которые у самца и самки разные, получила название идио, или гетерохромосомы, или половые хромосомы. У самки две одинаковые половые хромосомы, обозначаемые как Х-хромосомы, у самца одна Х-хромосома, другая - Y-хромосома. Остальные хромосомы одинаковые у самца и у самки, были названы аутосомами. Таким образом, хромосомная формула у самки названного клопа запишется 12A + XX, у самца 2A + XY. У ряда других организмов, хотя и существует в принципе тот же аппарат для определения пола, однако гетерозиготны в отношении реализаторов пола не мужские, а женские организмы. Особи мужского пола имеют две одинаковые половые хромосомы ZZ, а особи женского пола - ZO или ZW. ZZ-ZW тип определения пола наблюдается у бабочек, птиц, ZZ-ZO - ящериц, некоторых птиц.
Совершенно другой механизм определения пола, называемый гаплодиплоидный, широко распространен у пчел и муравьев. У этих организмов нет половых хромосом: самки - это диплоидные особи, а самцы (трутни) - гаплоидные. Самки развиваются из оплодотворенныз яиц, а из неоплодотворенных развиваются трутни.
Человек в отношении определения пола относится к типу XX-XY. При гаметогенезе наблюдается типичное менделевское расщепление по половым хромосомам. каждая яйцеклетка содержит одну Х-хромосому, а другая половина - одну Y-хромосому. Пол потомка зависит от того, какой спермий оплодотворит яйцеклетку. Пол с генотипом ХХ называют гомогаметным, так как у него образуются одинаковые гаметы, содержащие только Х-хромосомы, а пол с генотипом XY-гетерогаметным, так как половина гамет содержит Х-, а половина - Y-хромосому. У человека генотипический пол данного индивидума определяют, изучая неделящиеся клетки. Одна Х-хромосома всегда оказывается в активном состоянии и имеет обычный вид. Другая, если она имеется, бывает в покоящемся состоянии в виде плотного темно-окрашенного тельца, называемого тельцем Барра (факультативный гетерохроматин). Число телец Барра всегда на единицу меньше числа наличных х-хромосом, т.е. в мужском организме их нет вовсе, у женщин (ХХ) - одно. У человека Y-хромосома является генетически инертной, так как в ней очень мало генов (Асланян, М.М., 2010). Однако влияние Y-хромосомы на детерминацию пола у человека очень сильное. Хромосомная структура мужчины 44A+XY и женщины 44A+XX такая же, как и у дрозофины, однако у человека особь кариотипом 44A+XD оказалась женщиной, а особь 44A+XXY мужчиной. В обоих случаях они проявляли дефекты развития, но все же пол определялся наличием или отсутствием y-хромосомы. Люди генотипа XXX2A представляют собой бесплодную женщину, с генотипом XXXY2A - бесплодных умственно отстающих мужчин. Такие генотипы возникают в результате нерасхождения половых хромосом, что приводит к нарушению развития (например, синдром Клайнфельтера (XXY). Нерасхождение хромосом изучаются как в мейозе, так и в нитозе. Нерасхождение может быть следствием физического сцепления Х-хромосом, в таком случае нерасхождение имеет место в 100% случаев (Иванов В.И., 2006).
Рис.1. Вид половых хромосом человека в метафазе митоза
Всем млекопитающим мужского пола, включая человека, свойственен так называемый H-Y антиген, находящийся на поверхности клеток, несущих Y-хромосому. Единственной функцией его считается дифференцировка гонад. Вторичные половые признаки развиваются под влиянием стероидных гормонов, вырабатываемых гонадами. Развитие мужских вторичных половых признаков контролирует тестостерон, воздействующий на все клетки организма, включая клетки гонад. Мутация всего одного Х-хромосомы, кодирующего белок-рецептор тестостерона, приводит к синдрому тестикумерной фелинизации особей XY. Клетки-мутанты не чувствительны в действию тестостерона, в результате чего взрослый организм приобретает черты, характерные для женского пола. При этом внутренние половые органы оказываются недоразвитыми и такие особи полностью стерильные. Таким образом, в определении и дифференцировке пола млекопитающих и человека взаимодействуют хромосомный и генный механизмы.
Несмотря на то, что женщины имеют две Х-хромосомы, а мужчины - только одну, экспрессия генов Х-хромосомы происходит на одном и том же уровне у обоих полов. Это объясняется тем, что у женщин в каждой клетке полностью инактивирована одна Х-хромосома (тельце Барра), о чем уже было сказано выше. Х-хромосома инактивируется на ранней стадии эмбрионального развития, соответствующей времени имплантации. при этом в разных клетках отцовская и материнская Х-хромосомы выключаются случайно. Состояние инактивации данной Х-хромосомы наследуется в ряду клеточных делений. Таким образом, женские особи, гетерозиготные по генам половых хромосом, представляют собой мозаики (пример, черепаховые кошки).
Таким образом, пол человека представляет собой менделирующий признак, наследуемый по принципу обратного (анализирующего) скрещивания. Гетерозиготой оказывается гетерогаметный пол (XY), который скрещивается с рецессивной гомозиготой, представленной гомогаметным полом (XX). В результате в природе обнаруживается наследственная дифференцировка организмов на мужской и женский пол и устойчивое сокращение во всех поколениях количественного равенства полов.
1.2 Наследование признаков, сцепленных с полом
Морган и его сотрудники заметили, что наследование окраски глаз у дрозофилы зависит от пола родительских особей, несущих альтернативные аллели. Красная окраска глаз доминирует над белой. При скрещивании красноглазого самца с белоглазой самкой в F1, получали равное число красноглазых самок и белоглазых самцов. Однако при скрещивании белоглазого самца с красноглазой самкой в F1 были получены в равном числе красноглазые самцы и самки (Асланян, М.М., 2010). При скрещивании этих мух F1, между собой были получены красноглазые самки, красноглазые и белоглазые самцы, но не было ни одной белоглазой самки. Тот факт, что у самцов частота проявления рецессивного признака была выше, чем у самок, наводил на мысль, что рецессивный аллель, определяющий белоглазость, находится в Х - хромосоме, а Y - хромосома лишена гена окраски глаз. Чтобы проверить эту гипотезу, Морган скрестил исходного белоглазого самца с красноглазой самкой из F1. В потомстве были получены красноглазые и белоглазые самцы и самки. Из этого Морган справедливо заключил, что только Х - хромосома несет ген окраски глаз. В Y - хромосоме соответствующего локуса вообще нет. Это явление известно под названием наследования, сцепленного с полом.
Гены, находящиеся в половых хромосомах, называют сцепленными с полом. В Х-хромосоме имеется участок, для которого в Y-хромосоме нет гомолога. Поэтому у особей мужского пола признаки, определяемые генами этого участка, проявляются даже в том случае, если они рецессивны. Эта особая форма сцепления позволяет объяснить наследование признаков, сцепленных с полом.
При локализации признаков как в аутосоме, так и в Х- b Y-хромосоме наблюдается полное сцепление с полом.
У человека около 60 генов наследуются в связи с Х-хромосомой, в том числе гемофелия, дальтонизм (цветовая слепота), мускульная дистрофия, потемнение эмали зубов, одна из форм агаммглобулинемии и другие. Наследование таких признаков отклоняется от закономерностей, установленных Г.Менделем. Х-хромосома закономерно переходит от одного пола к другому, при этом дочь наследует Х-хромосому отца, а сын Х-хромосому матери (Иванов В.И., 2006). Наследование, при котором сыновья наследуют признак матери, а дочери - признак отца получило, название крисс-кросс (или крест-накрест).
Известны нарушения цветового зрения, так называемая цветовая слепота. В основе появления этих дефектов зрения лежит действие ряда генов. Красно-зеленая слепота обычно называется дальтонизмом. Еще задолго до появления генетики в конце XVIII и в XIX в. было установлено, что цветовая слепота наследуется согласно вполне закономерным правилам. Так, если женщина, страдающая цветовой слепотой, выходит замуж за мужчину с нормальным зрением, то у их детей наблюдается очень своеобразная картина перекрестного наследования. Все дочери от такого брака получат признак отца, т.е. они имеют нормальное зрение, а все сыновья, получая признак матери, страдают цветовой слепотой (а-дальтонизм, сцепленный с Х-хромосомой)
В том же случае, когда наоборот, отец является дальтоником, а мать имеет нормальное зрение, все дети оказываются нормальными. В отдельных браках, где мать и отец обладают нормальным зрением, половина сыновей может оказаться пораженными цветовой слепотой. В основном наличие цветовой слепоты чаще встречается у мужчин. Э.Вильсон объяснил наследование этого признака, предположив, что он локализовал в Х-хромосоме и что у человека гетерогаметным (XY) является мужской пол. Становится вполне понятным, что в браке гомозиготной нормальной женщины (Ха Ха) с мужчиной дальтоником (Хаy) все дети рождаются нормальными. Однако при этом, все дочери становятся скрытыми носителями дальтонизма, что может проявиться в последующих поколениях Гринев В.В., 2006).
Другим примером наследования сцепленного с полом, может послужить рецессивный полулетальный ген, вызывающий несвертываемость крови на воздухе - гемофилию. Это заболевание появляется почти исключительно только у мальчиков. При гемофилии нарушается образование фактора VIII, ускоряющего свертывание крови. ген, детерминирующий синтех фактора VIII, находится в участке Х-хромосомы, недоминантным нормальным и рецессивным мутантным. Возможны следующие генотипы и фенотипы:
Дальнейшие исследования генетиков показали, что законы Менделя о независимом наследовании признаков при дигибридном скрещивании применимы лишь тогда, когда разные гены располагаются в разных парах гомологичных хромосом. В том случае, если два гена находятся в одной паре гомологичных хромосом, расщепление в потомстве гибридов будет другим . У любого организма генов значительно больше, чем хромосом. Например, у человека имеется около миллиона генов, а хромосом всего 23 пары. Следовательно, в одной хромосоме размещается в среднем несколько тысяч генов. Гены, расположенные в одной хромосоме, называют сцепленными. Все гены этой хромосомы образуют группу сцепления, которая при мейозе обычно попадает в одну гамету.
Значит, гены, входящие в одну группу сцепления, не подчиняются закону независимого наследования, а при дигибридном скрещивании вместо ожидаемого расщепления по фенотипу в соотношении 9:3:3:1 дают соотношение 3:1, как при моногибридном скрещивании .
Закономерности сцепленного наследования были установлены американским биологом Томасом Морганом (1866-1945). В качестве объекта он использовал плодовую муху дрозофилу. У дрозофилы окраску тела и длину крыльев определяют следующие пары аллелей: А - серое тело, а - черное тело, В - длинные крылья, b - зачаточные крылья. Гены, отвечающие за окраску тела и длину крыльев, находятся в одной паре гомологичных хромосом и наследуются сцепленно.
Схематично пары гомологичных хромосом и локализованных в них генов можно изобразить так: ; . Для сравнения представим запись генов, локализованных в разных парах гомологичных хромосом: ; .
При скрещивании дрозофилы с серым телом и длинными крыльями с дрозофилой, имеющей черное тело и зачаточные крылья, все гибриды первого поколения имели серую окраску тела и длинные крылья .
При дальнейшем скрещивании между собой гибридных мух первого поколения в F2 не произошло ожидаемого расщепления по фенотипу 9:3:3:1. Вместо этого в F2 были получены мухи с родительскими фенотипами в соотношении примерно 3:1. Появление в F2 двух фенотипов вместо четырех позволило сделать вывод, что гены окраски тела и длины крыльев дрозофил находятся в одной хромосоме. Так был установлен закон Т.Моргана: гены, расположенные в одной хромосоме, наследуются совместно - сцепленно, то есть наследуются преимущественно вместе.
Однако при дигибридном скрещивании при сцепленном наследовании признаков не всегда появляются особи только двух фенотипов. Иногда появляются особи еще двух фенотипов с перекомбинацией (новым сочетанием) родительских признаков: серое тело - зачаточные крылья, черное тело - длинные крылья. (Особей с такими фенотипами немного - около 8,5% каждого типа.) Почему же нарушается сцепление генов и появляются особи с новыми фенотипами? Было установлено, что сцепление генов может быть полным и неполным.
Полное сцепление наблюдается в том случае, если скрещиваются серый самец с длинными крыльями и самка с черным телом и зачаточными крыльями. Расщепление по фенотипу в этом случае будет 1:1, то есть наблюдается полное сцепление генов в одной хромосоме .
При скрещивании серой длиннокрылой самки с самцом, имеющим черное тело и зачаточные крылья, расщепление по фенотипу будет примерно 41,5:41,5:8,5:8,5, что характеризует неполное сцепление. Причина нарушения сцепления заключается в том, что в ходе мейоза происходит кроссинговер и гомологичные хромосомы обмениваются своими участками. В результате гены, расположенные в одной из гомологичных хромосом, оказываются в другой хромосоме. Возникают новые сочетания признаков.
У самцов дрозофил в мейозе кроссинговер не происходит, поэтому при скрещивании серого длиннокрылого самца дрозофилы с рецессивной самкой с черным телом и зачаточными крыльями сцепление будет полным. Неполное сцепление наблюдается в том случае, если самка гетерозиготна, а самец гомозиготен. В данном примере кроссинговер происходит примерно у 17% самок.
Таким образом, если не происходит перекреста хромосом и обмена генами, то наблюдается полное сцепление генов. При наличии кроссинговера сцепление генов бывает неполным. Благодаря перекресту хромосом возникают новые сочетания генов и признаков. Чем дальше друг от друга расположены гены в хромосоме, тем больше вероятность перекреста между ними и обмена участками хромосом.
Результатом исследований Т.Х.Моргана стало создание им хромосомной теории наследственности:
1. Гены располагаются в хромосомах; различные хромосомы содержат неодинаковое число генов, причем набор генов каждой из негомологичных хромосом уникален;
2. Каждый ген имеет определенное место (локус) в хромосоме; в идентичных локусах гомологичных хромосом находятся аллельные гены;
3. Гены расположены в хромосомах в определенной линейной последовательности;
4. Гены, локализованные в одной хромосоме, наследуются совместно, образуя группу сцепления; число групп сцепления равно гаплоидному набору хромосом и постоянно для каждого вида организмов;
5. Сцепление генов может нарушаться в процессе кроссинговера; это приводит к образованию рекомбинантных хромосом;
6. Частота кроссинговера является функцией расстояния между генами: чем больше расстояние, тем больше величина кроссинговера (прямая зависимость);
7. Каждый вид имеет характерный только для него набор хромосом - кариотип.
2. История генетики в России
ДОСТИЖЕНИЯ ГЕНЕТИКИ В 20-е - 40-е ГОДЫ
В 1926 году С.С.Четвериков публикует большую программную статью о связи теории эволюции и генетики. Как и в случае с Менделем, эта статья знаменовала собой рождение новой области - генетики популяций. Она включала ряд новых концепций, предсказаний и описание методов их проверки. Прежде всего это концепция "мутационного давления", процесса возникновения новых наследственных изменений (мутаций) - столь же неизбежного для пригородных видов, сколь неизбежен радиоактивный распад. Каждый вид "впитывает" в себя вновь возникающие мутации, они накапливаются в скрытом состоянии и могут служить источником эволюционных преобразований. Был сделан важный концептуальный вывод, что накопленное генное разнообразие должно выявляться в условиях изоляции и уже без всякого отбора приводить к различиям между популяциями и индивидами в природе. Четвериков создал понятие "генотипическая среда", а А.С.Серебровский ввел другое, столь же известное ныне, понятие "генофонд" - для сопоставления генных различий между популяциями. Таким образом, удалось связать теорию Дарвина с менделеевской генетикой.
Способность к матричному самовоспроизведению нуклеиновых кислот ДНК и РНК рассматривается ныне как основа жизни. Но именно Н.К.Кольцов в 1927 году выдвинул концепцию, что хромосомы представляют собой гигантские молекулы, способные к самовоспроизведению. Этот постулат уже в 30-е годы получил косвенное подтверждение в начатых Тимофеевым-Ресовским в Германии работах по радиационной генетике. Их цель была установить, с какой частотой возникают мутации под действием разных доз и видов облучения. В итоге, количественные расчеты привели к важному выводу, что повреждения, вызываемые облучением, являются не мульти- а мономолекулярными. Это хорошо гармонировало с идеей Кольцова о хромосоме как одной гигантской молекуле. На основе выдвинутого "принципа попаданий" удалось впервые определить примерный молекулярный объем гена.
Попытка выяснить строение гена была предпринята в серии работ на дрозофиле А.С.Серебровского и его учеников (Н.П.Дубинин, Б.Н.Сидоров, И.И.Агол, Н.И.Шапиро). Атака на ген оказалась успешной. Впервые был сделан вывод о делимости гена и его сложной линейной структуре. В середине 30-х был открыт и изучен "эффект положения" генов, когда нормальный ген, будучи искусственно перенесен в другое место хромосомы, менял характер своего проявления (Н. П. Дубинин, Б.Н.Сидоров, В.В.Хвостова, А.А.Прокофьева-Бельговская). Этот феномен, связанный с регуляторными отношениями между генами, является и ныне одной из горячих точек современной науки.
Из работ отечественных генетиков, наибольшее мировое признание, получили, пожалуй, работы академика Н.И.Вавилова и его коллег по ВИРу. Вавилов был одновременно генетиком, систематиком, эволюционистом, физиологом растений, выдающимся организатором науки и общественным деятелем, а также крупным географом-путешественником. Отметим здесь только три его новые концепции: 1) закон гомологических рядов в наследственной изменчивости, 2) учение о центрах происхождения культурных растений; 3) представление о сложной полиморфной структуре биологических видов. Закон Вавилова устанавливал определенные правила формообразования и позволял предсказывать у данного вида, еще не открытые, но возможные признаки (аналогия с системой Менделеева).
Широкую известность получили работы сподвижников Вавилова в области изучения хромосом. Так, Г.А.Левитский ввел в биологию термин "кариотип" - для описания базовых особенностей морфологии хромосом одного вида и сравнения их между собой у разных организмов и видов. В 1934 году он впервые у растений показал, как под действием облучения хромосомы распадаются на фрагменты и перестраиваются.
Ныне, в конце ХХ века у всех на слуху слова "генная инженерия". Между тем, еще в 20-е годы ученик Вавилова Г.Д.Карпеченко, работая в ВИРе, создал удивительный метод хромосомной инженерии. Его работы входят теперь во все учебники по генетике. Он показал возможность преодоления бесплодия отделенных гибридов за счет удвоения наборов хромосом обоих родителей. Таким путем впервые были получены гибриды между капустой и редькой, а затем созданы новые виды пшениц при их отдаленной гибридизации друг с другом и с сородичами. Этим методом широко пользовалась и природа, создавая новые виды растений. Впоследствии ученик Четверикова академик Б.Л.Астауров путем хромосомной инженерии впервые получил отдаленные гибриды у животных на примере тутового шелкопряда.
В 1932 г. под впечатлением успехов генетики в России было решено провести очередной Международный Генетический Конгресс. Но на это советские власти не дали разрешения. Надвигалась эпоха Лысенко. К началу 40-х годов Вавилов и его коллеги Левитский, Карпеченко, Л.Говоров были репрессированы.
РЕПРЕССИИ И ПОГРОМ ГЕНЕТИКИ
Где корни случившегося в 1948 году погрома генетики и воцарения Лысенко? Первое после революции десятилетие - период бурного роста и успехов русской генетики, возникшей на крепком биологическом фундаменте. Отношение власти к науке было амбивалентное. С одной стороны, естественные науки, и в их числе генетика, получали солидную государственную поддержку. Открывались новые вузы, кафедры, музеи, под которые нередко отдавались старинные особняки и дворцы. Сотрудники и студенты были полны оптимизма и энтузиазма. Политика государственного попечительства совпадала с интересами и устремлениями таких научных гигантов, как Н.И.Вавилов. Эта невиданная ранее для стран Европы государственная поддержка пропагандировалась, поражала и гипнотизировала большинство западных ученых.
Режим фетишизировал науку, но одновременно низводил ее на роль служанки ("наука на службе социализма") в социалистической "перестройке" общества.
Все, что не соответствовало этим целям - подавлялось. Поэтому, одновременно с ростом естественных, уже в первые годы революции были просто разгромлены социальные науки: история, философия и те течения общественной мысли, которые хоть в малейшей мере оппонировали или выходили за рамки марксистской догмы. Наука попала в золотую клетку. С 1929 г. с началом Великого Перелома возрастает роль репрессивных органов. Одной из первых жертв стал профессор С.С.Четвериков и его лаборатория. По нелепому доносу его арестовывают и без суда и следствия ссылают в Свердловск. Он уже никогда не возвращается в Москву. Лаборатория распадается, ряд ее членов также подвергаются ссылке. Другие, спасаясь от репрессий, уезжают из Москвы.
Кольцова глубоко интересовала генетика человека. В своем ИЭГ он начал исследования по изучению близнецов и основал в 1922 г. "Русский евгенический журнал". В опубликованной в 1923 г. в этом журнале статье "Генетический анализ психических особенностей человека" Кольцов наметил программу исследования на десятилетия. В 1932 году по его инициативе был создан Медико-Биологический институт, работавший с таким энтузиазмом и энергией, что за 4 года вышло 4 тома оригинальных работ, многие из которых до сих пор не утратили актуальности. Однако, в 1936 году институт в одночасье был закрыт, а его директор С.Г.Левит вскоре расстрелян. Все работы по генетике человека и медицинской генетике были прерваны на четверть века. В итоге целые поколения врачей остались без столь необходимых генетических знаний.
Истории воцарения и господства Лысенко в 1948 г. посвящены многие книги. Отметим здесь главное. Боевая операция по разгрому генетики на Сессии ВАСХНИЛ в августе 1948 года, которую проводил Лысенко, была лично одобрена Сталиным.
.. Были сразу уволены десятки и сотни ведущих профессоров и преподавателей. Из библиотек изымались и уничтожались по спискам биологические книги, основанные на менделевской генетике. Пламя погрома перекинулось на цитологию, эмбриологию, физиологию и достигло даже таких отдаленных областей, как квантовая химия.
После смерти Сталина в 1953 г., в период "оттепели", усиливается противостояние лысенковскому обскурантизму. Начиная с 1953 г. известный эволюционист проф. А.А.Любищев и вернувшийся из лагеря генетик В.П.Эфроимсон посылают в ЦК партии, в журналы, ведущим биологам серии критических статей о монополии Лысенко в биологии, анализируя большой урон со стороны лысенковщины сельскому хозяйству, медицине, экономике. В 1955 г. в ЦК партии было направлено знаменитое "письмо трехсот", подписанное ведущими биологами, затем к нему присоеденились письмо ряда академиков-физиков. В 1956 г. проф. М.Е.Лобашев начинает читать курс классической генетики на возглавляемой им кафедре генетики в Ленинградском университете. В это же время в Институте биофизики и Институте атомной энергии создаются генетические лаборатории, а затем в 1957 г. Институт цитологии и генетики в Сибирском отделении АН СССР (Академгородок, Новосибирск).
Вместе с тем, еще в декабре 1958 г. была разогнана редакция "Ботанического журнала" во главе с акад. В.Н.Сукачевым за публикацию серии критических статей об идеях Лысенко. В 1963 г. такая же участь постигла журнал "Нева" за яркую и смелую статью генетиков В.С.Кирпичникова и Ж.А.Медведева "Перспективы советской генетики". Однако явное падение Лысенко началось лишь вслед за падением Н.С.Хрущева в 1964 г. В сентябре 1965 года на заседании Президиума АН под руководством акад. М.В.Келдыша впервые, наконец, открыто подверглись критике методы и результаты деятельности Лысенко. В 1965 г. он был снят с поста директора академического Института генетики, который он занимал целых четверть века лет после ареста Вавилова, навязывая через систему государственных учреждений свои бредни.
В этом кратком очерке можно назвать лишь наиболее важные работы отечественных генетиков последней трети ХХ века. К ним, в первую очередь, относится, сделанное И.А.Рапопортом открытие супермутагенов - веществ, в десятки и сотни раз повышающих частоту возникновения мутаций у самых разных организмов. С использованием супермутагенов сделаны важные работы в теории мутаций, получены новые штаммы антибиотиков и новые сорта растений (Рапопорт останется в истории генетики и как единственный биолог, который в 1948 году открыто отказался признать лысенкоизм).
Несомненным достижением отечественной генетики является открытие у животных на примере дрозофилы "прыгающих генов" и свидетельства того, что эти гены вызывают вспышки нестабильных мутаций в лаборатории и природе и связаны с адаптивными преобразованиями генетической системы клетки. Оригинальные результаты, полученные в рамках этого направления российскими генетиками, включая мировые исследования по этой проблеме, были обобщены в замечательной сводке Р.Б.Хесина "Непостоянство генома". Эта сводка несомненно войдет в золотой фонд российской науки. В ней обосновано положение о потенциальном единстве генофонда земных организмов за счет горизонтального переноса генов вирусами и другими подвижными элементами. С именем Р.Б.Хесина, ученика А.С.Серебровского, связано зарождение и развитие молекулярной генетики в стране, которое происходило под крышей Института атомной энергии.
Блестящий цитолог и генетик В.В.Прокофьева-Бельговская, ученица Ю.А.Филипченко, создала школу цитогенетиков, изучающих поведение и структуру хромосом человека в норме и патологии ("хромосомные болезни"). Вместе с другим генетиком, В.П.Эфроимсоном, они возродили исследования по медицинской генетике. Однако влияние идеологических запретов на изучение наследственности человека оказалось столь велико, что книга В.П.Эфроимсона "Генетика гениальности" более 20 лет не могла пробиться в печать и вышла лишь в 1998 году.
Если век 19-й по праву вошел в историю мировой цивилизации как Век Физики, то стремительно завершающемуся веку 20-му, в котором нам счастливилось жить, по всей вероятности, уготовано место Века Биологии, а может быть, и Века Генетики.
Действительно, за неполных 100 лет после вторичного открытия законов Г. Менделя генетика прошла триумфальный путь от натурфилосовского понимания законов наследственности и изменчивости через экспериментальное накопление фактов формальной генетики к молекулярно-биологическому пониманию сущности гена, его структуры и функции. От теоретических построений о гене как абстрактной единице наследственности - к пониманию его материальной природы как фрагмента молекулы ДНК, кодирующего аминокислотную структуру белка, до клонирования индивидуальных генов, создания подробных генетических карт человека, животных, идентификации генов, мутации которых сопряжены с тяжелыми наследственными недугами, разработки методов биотехнологии и генной инженерии, позволяющих направленно получать организмы с заданными наследственными признаками, а также проводить направленную коррекцию мутантных генов человека, т.е. генотерапию наследственных заболеваний. Молекулярная генетика значительно углубила наши представления о сущности жизни, эволюции живой природы, структурно-функциональных механизмов регуляции индивидуального развития. Благодаря ее успехам начато решение глобальных проблем человечества, связанных с охраной его генофонда.
1. Г.Д. Бердышев, В.А. Ратнер “Код наследственности”. Новосибирское книжное издательство. 1963 год.
2. Н. П. Дубинин. “Генетика и человек”. М.,”Просвещение”, 1978.
3. Е.Ф. Давиденкова, А.Б. Чухловин. “О наследственности”. М.,”Медицина”, 1975.
4. Ф. Полканов. “Мы и ее величество ДНК”. М., “Детская литература”, 1968.
* Данная работа не является научным трудом, не является выпускной квалификационной работой и представляет собой результат обработки, структурирования и форматирования собранной информации, предназначенной для использования в качестве источника материала при самостоятельной подготовки учебных работ.
1. Генетика пола. 2
2.1. Генетические механизмы формирования пола. 3
2.2. Наследование признаков, сцепленных с полом. 6
2.3. Наследование признаков, контролируемых полом. 11
3. Сцепленное наследование признаков. 11
3.1. Хромосомная теория наследственности. 12
3.2. Механизм сцепления. 13
3.3. Кроссинговер. 14
3.4. Группы сцепления и карты хромосом у человека. 18
4. Заключение. 19
5. Библиографический список. 20
Введение.
Генетика является одной из самых прогрессивных наук естествознания. Ее достижения изменили естественнонаучное и во многом философское понимание явлений жизни. Роль генетики для практики селекции и медицины очень велика. Значение генетики для медицины будет возрастать с каждым годом, ибо генетика касается самых сокровенных сторон биологии и физиологии человека. Благодаря генетике, ее знаниям, разрабатываются методы лечения ряда наследственных заболеваний, таких, как фенилкетонурия, сахарный диабет и другие. Здсь медико-генетическая работа призвана облегчить страдания людей от действия дефектных генов, полученных ими от родителей. Внедряются в практику приемы медико-генетического консультирования и прентальной диагностики, что позволяет предупредить развитие наследственных заболеваний.
1. Генетика пола.
Пол - совокупность признаков, по которым производится специфическое разделение особей или клеток, основанное на морфологических и физиологических особенностях, позволяющее осуществлять в процессе полового размножения комбинирование в потомках наследственных задатков родителей.
Морфологические и физиологические признаки, по которым производится специфическое разделение особей, называется половым.
Признаки, связанные с формированием и функционированием половых клеток, называется первичными половыми признаками. Это гонады (яичники или семенники), их выводные протоки, добавочные железы полового аппарата, копулятивные органы. Все другие признаки, по которым один пол отличается од другого, получили название вторичных половых признаков. К ним относят: характер волосяного покрова, наличие и развитие молочных желез, строение скелета, тип развития подкожной жировой клетчатки, строение трубчатых костей и др.
2.1. Генетические механизмы формирования пола.
Начало изучению генотипического определения пола было положено открытием американскими цитологами у насекомых различия в форме, а иногда и в числе хромосом у особей разного пола (Мак-Кланг, 1906, Уилсон, 1906) и классическими опытами немецкого генетика Корренса по скрещиванию однодомного и двудомного видов брионии. Уилсон обнаружил, что у клопа Lydaeus turucus самки имеют 7 пар хромосом, у самцов же 6 пар одинаковых с самкой хромосом, а в седьмой паре одна хромосома такая же, как соответствующая хромосома самки, а другая маленькая.
Пара хромосом, которые у самца и самки разные, получила название идио, или гетерохромосомы, или половые хромосомы. У самки две одинаковые половые хромосомы, обозначаемые как Х-хромосомы, у самца одна Х-хромосома, другая - Y-хромосома. Остальные хромосомы одинаковые у самца и у самки, были названы аутосомами. Таким образом, хромосомная формула у самки названного клопа запишется 12A + XX, у самца 2A + XY. У ряда других организмов, хотя и существует в принципе тот же аппарат для определения пола, однако гетерозиготны в отношении реализаторов пола не мужские, а женские организмы. Особи мужского пола имеют две одинаковые половые хромосомы ZZ, а особи женского пола - ZO или ZW. ZZ-ZW тип определения пола наблюдается у бабочек, птиц, ZZ-ZO - ящериц, некоторых птиц.
Совершенно другой механизм определения пола, называемый гаплодиплоидный, широко распространен у пчел и муравьев. У этих организмов нет половых хромосом: самки - это диплоидные особи, а самцы (трутни) - гаплоидные. Самки развиваются из оплодотворенныз яиц, а из неоплодотворенных развиваются трутни.
Человек в отношении определения пола относится к типу XX-XY. При гаметогенезе наблюдается типичное менделевское расщепление по половым хромосомам. каждая яйцеклетка содержит одну Х-хромосому, а другая половина - одну Y-хромосому. Пол потомка зависит от того, какой спермий оплодотворит яйцеклетку. Пол с генотипом ХХ называют гомогаметным, так как у него образуются одинаковые гаметы, содержащие только Х-хромосомы, а пол с генотипом XY-гетерогаметным, так как половина гамет содержит Х-, а половина - Y-хромосому. У человека генотипический пол данного индивидума определяют, изучая неделящиеся клетки. Одна Х-хромосома всегда оказывается в активном состоянии и имеет обычный вид. Другая, если она имеется, бывает в покоящемся состоянии в виде плотного темно-окрашенного тельца, называемого тельцем Барра (факультативный гетерохроматин). Число телец Барра всегда на единицу меньше числа наличных х-хромосом, т.е. в мужском организме их нет вовсе, у женщин (ХХ) - одно. У человека Y-хромосома является генетически инертной, так как в ней очень мало генов. Однако влияние Y-хромосомы на детерминацию пола у человека очень сильное. Хромосомная структура мужчины 44A+XY и женщины 44A+XX такая же, как и у дрозофины, однако у человека особь кариотипом 44A+XD оказалась женщиной, а особь 44A+XXY мужчиной. В обоих случаях они проявляли дефекты развития, но все же пол определялся наличием или отсутствием y-хромосомы. Люди генотипа XXX2A представляют собой бесплодную женщину, с генотипом XXXY2A - бесплодных умственно отстающих мужчин. Такие генотипы возникают в результате нерасхождения половых хромосом, что приводит к нарушению развития (например, синдром Клайнфельтера (XXY). Нерасхождение хромосом изучаются как в мейозе, так и в нитозе. Нерасхождение может быть следствием физического сцепления Х-хромосом, в таком случае нерасхождение имеет место в 100% случаев.
Рис.1. Вид половых хромосом человека в метафазе митоза.
Всем млекопитающим мужского пола, включая человека, свойственен так называемый H-Y антиген, находящийся на поверхности клеток, несущих Y-хромосому. Единственной функцией его считается дифференцировка гонад. Вторичные половые признаки развиваются под влиянием стероидных гормонов, вырабатываемых гонадами. Развитие мужских вторичных половых признаков контролирует тестостерон, воздействующий на все клетки организма, включая клетки гонад. Мутация всего одного Х-хромосомы, кодирующего белок-рецептор тестостерона, приводит к синдрому тестикумерной фелинизации особей XY. Клетки-мутанты не чувствительны в действию тестостерона, в результате чего взрослый организм приобретает черты, характерные для женского пола. При этом внутренние половые органы оказываются недоразвитыми и такие особи полностью стерильные. Таким образом, в определении и дифференцировке пола млекопитающих и человека взаимодействуют хромосомный и генный механизмы.
Несмотря на то, что женщины имеют две Х-хромосомы, а мужчины - только одну, экспрессия генов Х-хромосомы происходит на одном и том же уровне у обоих полов. Это объясняется тем, что у женщин в каждой клетке полностью инактивирована одна Х-хромосома (тельце Барра), о чем уже было сказано выше. Х-хромосома инактивируется на ранней стадии эмбрионального развития, соответствующей времени имплантации. при этом в разных клетках отцовская и материнская Х-хромосомы выключаются случайно. Состояние инактивации данной Х-хромосомы наследуется в ряду клеточных делений. Таким образом, женские особи, гетерозиготные по генам половых хромосом, представляют собой мозаики (пример, черепаховые кошки).
Таким образом, пол человека представляет собой менделирующий признак, наследуемый по принципу обратного (анализирующего) скрещивания. Гетерозиготой оказывается гетерогаметный пол (XY), который скрещивается с рецессивной гомозиготой, представленной гомогаметным полом (XX). В результате в природе обнаруживается наследственная дифференцировка организмов на мужской и женский пол и устойчивое сокращение во всех поколениях количественного равенства полов.
2.2. Наследование признаков, сцепленных с полом.
Морган и его сотрудники заметили, что наследование окраски глаз у дрозофилы зависит от пола родительских особей, несущих альтернативные аллели. Красная окраска глаз доминирует над белой. При скрещивании красноглазого самца с белоглазой самкой в F1, получали равное число красноглазых самок и белоглазых самцов. Однако при скрещивании белоглазого самца с красноглазой самкой в F1 были получены в равном числе красноглазые самцы и самки. При скрещивании этих мух F1, между собой были получены красноглазые самки, красноглазые и белоглазые самцы, но не было ни одной белоглазой самки. Тот факт, что у самцов частота проявления рецессивного признака была выше, чем у самок, наводил на мысль, что рецессивный аллель, определяющий белоглазость, находится в Х - хромосоме, а Y - хромосома лишена гена окраски глаз. Чтобы проверить эту гипотезу, Морган скрестил исходного белоглазого самца с красноглазой самкой из F1. В потомстве были получены красноглазые и белоглазые самцы и самки. Из этого Морган справедливо заключил, что только Х - хромосома несет ген окраски глаз. В Y - хромосоме соответствующего локуса вообще нет. Это явление известно под названием наследования, сцепленного с полом.
Гены, находящиеся в половых хромосомах, называют сцепленными с полом. В Х-хромосоме имеется участок, для которого в Y-хромосоме нет гомолога. Поэтому у особей мужского пола признаки, определяемые генами этого участка, проявляются даже в том случае, если они рецессивны. Эта особая форма сцепления позволяет объяснить наследование признаков, сцепленных с полом.
При локализации признаков как в аутосоме, так и в Х- b Y-хромосоме наблюдается полное сцепление с полом.
У человека около 60 генов наследуются в связи с Х-хромосомой, в том числе гемофелия, дальтонизм (цветовая слепота), мускульная дистрофия, потемнение эмали зубов, одна из форм агаммглобулинемии и другие. Наследование таких признаков отклоняется от закономерностей, установленных Г.Менделем. Х-хромосома закономерно переходит от одного пола к другому, при этом дочь наследует Х-хромосому отца, а сын Х-хромосому матери. Наследование, при котором сыновья наследуют признак матери, а дочери - признак отца получило, название крисс-кросс (или крест-накрест).
Известны нарушения цветового зрения, так называемая цветовая слепота. В основе появления этих дефектов зрения лежит действие ряда генов. Красно-зеленая слепота обычно называется дальтонизмом. Еще задолго до появления генетики в конце XVIII и в XIX в. было установлено, что цветовая слепота наследуется согласно вполне закономерным правилам. Так, если женщина, страдающая цветовой слепотой, выходит замуж за мужчину с нормальным зрением, то у их детей наблюдается очень своеобразная картина перекрестного наследования. Все дочери от такого брака получат признак отца, т.е. они имеют нормальное зрение, а все сыновья, получая признак матери, страдают цветовой слепотой (а-дальтонизм, сцепленный с Х-хромосомой)
Р Х а Х а х Х а y
F1 Х а Х а , Х а y
В том же случае, когда наоборот, отец является дальтоником, а мать имеет нормальное зрение, все дети оказываются нормальными. В отдельных браках, где мать и отец обладают нормальным зрением, половина сыновей может оказаться пораженными цветовой слепотой. В основном наличие цветовой слепоты чаще встречается у мужчин. Э.Вильсон объяснил наследование этого признака, предположив, что он локализовал в Х-хромосоме и что у человека гетерогаметным (XY) является мужской пол. Становится вполне понятным, что в браке гомозиготной нормальной женщины (Х а Х а ) с мужчиной дальтоником (Х а y) все дети рождаются нормальными. Однако при этом, все дочери становятся скрытыми носителями дальтонизма, что может проявиться в последующих поколениях.
Другим примером наследования сцепленного с полом, может послужить рецессивныйполулетальный ген, вызывающий несвертываемость крови на воздухе - гемофилию. Это заболевание появляется почти исключительно только у мальчиков. При гемофилии нарушается образование фактора VIII, ускоряющего свертывание крови. ген, детерминирующий синтех фактора VIII, находится в участке Х-хромосомы, недоминантным нормальным и рецессивным мутантным. Возможны следующие генотипы и фенотипы:
Читайте также: