
Реферат по теме произвольные движения
Обновлено: 04.07.2024
Рассмотрим общую схему управления движениями. Решающим фактором поведения является полезный результат. Для его достижения в нервной системе формируется группа взаимосвязанных нейронов — функциональная система. Деятельность ее включает следующие процессы: обработка всех сигналов, поступающих из внешней и внутренней среды организма — так называемый афферентный синтез; принятие решения о цели и задачах действия; создание представления об ожидаемом результате и формирование конкретной программы движений; анализ полученного результата и внесение в программу поправок — сенсорных коррекций.
В процессах афферентного синтеза участвуют глубокие внутренние процессы — побуждение к действию (мотивация) и его замысел, извлекаются из памяти моторные следы (навыки) и выученные тактические комбинации. У человека на их основе создается определенный план и конкретная программа движения. Эти процессы отражаются в изменениях электрической активности мозга. На уровне спинного мозга процессы преднастройки отражаются повышением возбудимости спиральных мотонейронов, в мышцах — повышением чувствительности проприорецепторов скелетных мышц. Сенсорная информация о результате выполнения движения, получаемая по каналам обратной связи, используется нервными центрами для уточнения временных, пространственных и силовых характеристик двигательных актов, внесения поправок в команды. Такие поправки называются сенсорными коррекциями.
В живом организме имеет место рефлекторное кольцевое регулирование и программное управление движениями.
В двигательной деятельности человека различают произвольные движения (сознательно управляемые целенаправленные действия) и непроизвольные движения, котрые происходят без участия сознания и представляют собой либо безусловные реакции, либо автоматизированные двигательные навыки. В основе управления произвольными движениями человека лежат два различных физиологических механизма: рефлекторное кольцевое регулирование и программное управление по механизму центральных команд.
Замкнутая система рефлекторного кольцевого регулирования характерна для осуществления различных форм двигательных действий и позных реакций, не требующих быстрого двигательного акта. Это позволяет нервным центрам получать информацию о состоянии мышц и результатах их действий по различным афферентным путям и вносить поправки в моторные команды по ходу действия.
Программное управление по механизму центральных команд — это механизм регуляции движений, независимый от афферентных проприоцептивных влияний. Такое управление используется в случае выполнения кратковременных движений (прыжков, бросков, ударов ), когда организм не успевает использовать информацию от проприорецепторов мышц и других рецепторов. Вся программа должна быть готова еще до начала двигательного акта. При этом отсутствует замкнутое кольцо регуляции. Управление производится по так называемой открытой петле, а активность во многих произвольно сокращающихся мышцах возникает раньше, чем регистрируется обратная афферентная импульсация. Например, при выполнении прыжковых движений электрическая активность в мышцах, направленная на амортизацию удара, возникает раньше, чем происходит соприкосновение с опорой, т. е. она носит предупредительный характер.
Такие центральные программы создаются согласно сформированному в мозге (главным образом — в ассоциативной передне-лобной области коры) образу двигательного действия и цели движения. В дальнейшей конкретной разработке моторной программы принимают участие мозжечок (латеральная область его коры) и базальные ядра (полосатое тело и бледный шар). Информация от них поступает через таламус в моторную и премоторную области коры и далее — к исполнительным центрам спинного мозга и скелетным мышцам.
Механизм кольцевого регулирования является филогенетически более древним и возникает раньше в процессе индивидуального развития. Примерно к трем годам достаточное развитие получают зрительные обратные связи, осуществляющие текущий зрительно-моторный контроль, а с 5-6 лет происходит переход к текущему контролю движений с участием проприоцептивных обратных связей. Этот механизм достигает значительного совершенства к 7-9 летнему возрасту, после чего начинается переход к формированию механизма центральных команд. К 10-11 годам повышение скорости произвольных движений обеспечивается достаточным развитием процессов предварительного программирования их пространственных и временных параметров. С этого возраста представлены оба механизма управления произвольными движениями, дальнейшее совершенствование которых продолжается вплоть до 17-19 лет.
Существует три основных функциональных блока мозга, выделяемые в многоэтажных системах нервных центрах: блок регуляции тонуса, уровня бодрствования; блок приема, переработки и хранения информации; блок программирования, регуляциии контроля двигательной деятельности.
К первому функциональному блоку относятся неспецифические отделы нервной системы, в частности ретикулярная формация ствола мозга, которые модулируют функциональное состояние вышележащих и нижележащих отделов, вызывая состояния сна, бодрствования, повышенной активности, увеличивая или уменьшая мощность двигательных реакций.
Третий функциональный блок расположен в передних отделах больших полушарий. В его состав входят первичные (моторные) и вторичные (премоторные) поля, а высшим отделом являются ассоциативные передне-лобные (или префронтальные) области (передние третичные поля). Этот блок с участием речевых функций выполняет универсальную реакцию общей регуляции поведения, формируя намерения и планы, программы произвольных движений и контроль их выполнения.
8.2.2 Регуляция позно-тонических реакций различными отделами центральной нервной системы (ЦНС)
Мышечная деятельность включает в себя процессы осуществления двигательных актов и процессы поддержания позы тела. Эти процессы регулируются различными отделами ЦНС.
Рассмотрим роль спинного мозга в регуляции позно-тонических реакций. Мышечный тонус по своей природе является рефлекторным актом. Для его возникновения достаточна рефлекторная деятельность спинного мозга. При длительном растяжении мышц в поле силы тяжести возникает постоянное раздражение их проприорецепторов, потоки импульсов от которых проходят по толстым афферентным волокнам в спинной мозг, где передаются непосредственно (без участия вставочных нейронов) на альфа-мотонейроны передних рогов и вызывают тоническое напряжение мышц. Такие двухнейронные (или моносинаптические) рефлекторные дуги лежат в основе тонических сухожильных (с рецепторов сухожилий) и миостатических рефлексов на растяжение (с рецепторов мышечных веретен). Это рефлексы активного противодействия мышцы ее растяжению. В произвольной двигательной деятельности человека иногда требуется подавление этих рефлексов, например, при выполнении шпагата.
Степень тонического напряжения мышцы зависит от частоты импульсов; посылаемых к ней альфа-мотонейронами. Однако, потоки этих импульсов могут регулироваться вышележащими этажами нервной системы, в частности, неспецифическими отделами ствола мозга с помощью так называемой гамма-регуляции. Разряды гамма-мотонейронов спинного мозга под влиянием ретикулярной формации повышают чувствительность рецепторов мышечных веретен. В результате при той же длине мышцы увеличивается поток импульсов от рецепторов к альфа-мотонейронам и далее к мышце, повышая ее тонус.
В регуляции тонуса участвует также медленная часть пирамидной системы и различные структуры экстрапирамидной системы головного мозга:подкорковые ядра; красные ядра и черная субстанция среднего мозга; мозжечок; ретикулярная формация ствола мозга; вестибулярные ядра продолговатого мозга.
Неспецифическая система вызывает общее изменение тонуса различных мышц: усиление тонуса осуществляет активирующий отдел ретикулярной формации среднего мозга, а угнетение — тормозящий отдел продолговатого мозга. В отличие от нее специфическая система (моторные центры коры больших полушарий и ствола мозга) действует избирательно, т. е. на отдельные группы мышц-сгибателей или разгибателей. Усиление тонуса мышц-сгибателей вызывают корковые влияния, передающиеся непосредственно к нейронам спинного мозга по корково-спинномозговой (пирамидной) системе, а также через красные ядра (по корково-красноядерно-спинномозговой системе) и частично через ретикулярную формацию (по ретикуло-спинномозговой системе). В противоположность им, влияния, передающиеся через вестибулярные ядра продолговатого мозга к вставочным и двигательным нейронам передних рогов спинного мозга (по вестибуло-спинномозговым путям), вызывают тоническое повышение возбудимости мотонейронов мышц-разгибателей, что обеспечивает повышение тонуса этих мышц.
Мозжечок формирует правильное распределение тонуса скелетных мышц: через красные ядра среднего мозга он повышает тонус мышц-сгибателей, а через вестибулярные ядра продолговатого мозга — усиливает тонус мышц-разгибателей. В поддержании позы и равновесия тела, регуляции тонуса мышц основное значение имеет медиальная продольная зона мозжечка — кора червя. При мозжечковых расстройствах падает тоническое напряжение мышц (атония) и вследствие ненормального распределения тонуса мышц конечностей возникает нарушение походки (атаксия).
Бледный шар угнетает тонус мышц, а полосатое тело снижает его угнетающее действие.
Высший контроль тонической активности мышц осуществляет кора больших полушарий, в частности ее моторные, премоторные и лобные области. С ее участием происходит выбор наиболее целесообразной для данного момента позы тела, обеспечивается ее соответствие двигательной задаче. Непосредственное отношение к регуляции тонуса мышц имеют медленные пирамидные нейроны положения. Корковые влияния натонические реакции мышц передаются через медленную часть пирамидного тракта и через экстрапирамидную систему.
Специальная группа рефлексов способствует сохранению позы — это так называемые установочные рефлексы. К ним относятся статические и статокинетические рефлексы, в осуществлении которых большое значение имеют продолговатый и средний мозг.
Статические рефлексы возникают при изменении положения тела или его частей в пространстве в нескольких случаях. При изменениях положения головы в пространстве возникают лабиринтные рефлексы (при раздражении рецепторов вестибулярного аппарата). Шейные рефлексы возникают с проприорецепторов мышц шеи при изменении положения головы по отношению к туловищу, выпрямительные рефлексы — с рецепторов кожи, вестибулярного аппарата и сетчатки глаза. Например, при отклонении головы назад повышается тонус мышц-разгибателей спины, а при наклоне вперед — тонус мышц-сгибателей (лабиринтный рефлекс). С помощью выпрямительного рефлекса происходят последовательные сокращения мышц шеи и туловища, а затем и конечностей. Этот рефлекс обеспечивает вертикальное положение тела теменем кверху. У человека он проявляется, например, при нырянии.
Статокинетические рефлексы компенсируют отклонения тела при ускорении или замедлении прямолинейного движения (лифтный рефлекс), а также при вращениях (отклонения головы, тела и глаз в сторону, противоположную движению). Перемещение глаз со скоростью вращения тела, но в противоположную сторону, и быстрое их возвращение в исходное положение — нистагм глаз — обеспечивает сохранение изображения внешнего мира на сетчатке глаз и тем самым зрительную ориентацию.
8.2.3 Регуляция движений различными отделами ЦНС
Спинной мозг обеспечивает протекание многих элементарных двигательных рефлексов, включение которых в сложные двигательные акты и регуляция по мощности, пространственной ориентации и моменту включения осуществляется вышележащими отделами головного мозга под контролем коры больших полушарий.
Рассмотрим роль спинного мозга и подкорковых отделов ЦНС в регуляции движений. Спинной мозг осуществляет ряд элементарных двигательных рефлексов: рефлексы на растяжение (миостатические и сухожильные рефлексы, например, коленный рефлекс), кожные сгиба тельные рефлексы (например, защитный рефлекс отдергивания конечности при уколах, ожогах), разгиба тельные рефлексы (рефлекс отталкивания от опоры, лежащий в основе стояния, ходьбы, бега), перекрестные рефлексы и др.
Элементарные двигательные рефлексы включаются в более сложные двигательные акты — регуляцию деятельности мышц-антагонистов, ритмических и шагательных рефлексов, лежащих в основе локомоций и других движений.
Для сгибательного движения в суставе необходимо не только сокращение мышц-сгибателей, но и одновременное расслабление мышц-разгибателей. При этом в мотонейронах мышц-сгибателей возникает процесс возбуждения, а в мотонейронах мышц-разгибателей — торможение. При разгибании сустава, наоборот, тормозятся центры сгибателей и возбуждаются центры разгибателей. Такие координационные взаимоотношения между спинальными моторными центрами названы реципрокной (взаимосочетанной) иннервацией мышц-антагонистов. Однако реципрокные отношения между центрами мышц-антагонистов в необходимых ситуациях (например, при фиксации суставов, при точностных движениях) могут сменяться одновременным их возбуждением.
Нейроны промежуточной продольной зоны коры мозжечка согласуют позные реакции с движениями. Они выполняют также точные расчеты по ходу движений, необходимые для коррекции ошибок и адаптации моторных программ к текущей ситуации. Программирование каждого последующего шага осуществляется ими на основе анализа предыдущего. Кроме того производится согласование движений рук и ног, и особенно — регуляция активности мышц-разгибателей, обеспечивающих опорную фазу движения. Значение мозжечка в четком поддержании темпа ритмических движений объясняют геометрически правильным чередованием рядов эфферентных клеток Пуркинье и походящих к ним афферентных волокон.
К управлению ритмическими движениями непосредственное отношение имеют активирующие и угнетающие отделы ретикулярной формации, которые влияют на силу и темп сокращения мышц, а также подкорковые ядра, которые организуют автоматическое их протекание и содружественные движения конечностей. Включение древних форм ритмических движений (циклоидных) в акт письма позволяет человеку перейти от отдельного начертания букв к обычной письменной скорописи. То же самое происходит при освоении акта ходьбы — с переходом от отдельных шагов к ритмической походке. Плавность ритмических движений, четкое чередование реципрокных сокращений мышц обеспечивают премоторные отделы коры.
В высшей регуляции произвольных движений важнейшая роль принадлежит передне-лобным областям (передним третичным полям). Здесь помимо обычных вертикальных колонок нейронов существует принципиально новый тип функциональной единицы — в форме замкнутого нейронного кольца. Циркуляция импульсов в этой замкнутой системе обеспечивает кратковременную память. Она сохраняет в коре возбуждение между временем прихода сенсорных сигналов и формированием ответной эфферентной команды. Такой механизм служит основой сенсомоторной интеграции при программировании движений, при осуществлении зрительно-двигательных реакций.
Функцией передне-лобной (третичной) области коры является сознательная оценка текущей ситуации и предвидение возможного будущего, выработка цели и задачи поведения, программирование произвольных движений, их контроль и коррекция. Соответствие выполняемых действий поставленным задачам придает движениям человека определенную целесообразность и осмысленность. При поражении лобных долей движения человека становятся бессмысленными.
Необходимо рассмотреть и речевую регуляцию движений. Спецификой регуляции движений у человека является то, что они подчинены речевым воздействиям, т. е. могут программироваться лобными долями в ответ на поступающие извне словесные сигналы, а также благодаря участию внешней или внутренней речи (мышления) самого человека. В этой функции принимают участие расположенные в левом полушарии человека сенсорный центр речи Вернике и моторный центр речи — центр Брока. Считают, что афферентная импульсация от речевой мускулатуры является важным ориентиром, дополняющим проприоцептивные сигналы от работающих мышц, а формирующиеся на речевой основе избирательные связи в коре облегчают составление моторных программ.
Эта управляющая система еще не развита у ребенка 2-3 лет. Она появляется лишь к 3-4 годам. Внешняя речь, сменяясь постепенно шепотом и переходя затем во внутреннюю речь, становится важным регулятором моторных действий взрослого человека.
Основные принципы и схема управления движениями. Характеристика основных функциональных блоков мозга (мозжечка, коры и ствола головного мозга). Роль различных отделов центральной нервной системы в регуляции движений. Сущность речевой регуляции движений.
| Рубрика | Медицина |
| Вид | реферат |
| Язык | русский |
| Дата добавления | 21.02.2011 |
| Размер файла | 307,7 K |
Студенты, аспиранты, молодые ученые, использующие базу знаний в своей учебе и работе, будут вам очень благодарны.
1.Основные принципы организации движений
1.1 Общая схема управления движениями
1.2 Рефлекторное кольцевое регулирование и программное управление движениями
1.3 Три основных функциональных блока мозга
2. Роль различных отделов ЦНС в регуляции познотонических реакций
2.1 Роль спинного мозга
2.2 Роль коры головного мозга, мозжечка и ствола мозга
3. Рефлексы поддержания позы (установочные)
4. Роль различных отделов ЦНС в регуляции движений
4.1 Роль спинного мозга и подкорковых отделов ЦНС в регуляции движений
4.2 Роль различных отделов коры больших полушарий
4.3 Речевая регуляция движений
4.4 Нисходящие моторные системы
1. Основные принципы организации движений
Выполнение двигательных актов осуществляется обширным комплексом нейронов, расположенных в различных отделах ЦНС. Такая функциональная система управления движениями является многоэтажной и многоуровневой.
1.1 Общая схема управления движениями
1.2 Рефлекторное кольцевое регулирование и программное управление движениями
В двигательной деятельности человека различают произвольные движения - сознательно управляемые целенаправленные действиям непроизвольные движения, происходящие без участия сознания и представляющие собой либо безусловные реакции, либо автоматизированные двигательные навыки. В основе управления произвольными движениями человека лежат два различных физиологических механизма: 1) рефлекторное кольцевое регулирование и 2) программное управление по механизму центральных команд.Замкнутая система рефлекторного кольцевого регулирования характерна для осуществления различных форм двигательных действий и позных реакций, не требующих быстрого двигательного акта. Это позволяет нервным центрам получать информацию о состоянии мышц и результатах их действий по различным афферентным путям и вносить поправки в моторные команды по ходу действия.Программное управление по механизму центральных команд -- это механизм регуляции движений, независимый от афферентных проприоцептивных влияний. Такое управление используется в случае выполнения кратковременных движений (прыжков, бросков, ударов, метаний), когда организм не успевает использовать информацию от проприорецепторов мышц и других рецепторов. Вся программа должна быть готова еще до начала двигательного акта. При этом отсутствует замкнутое кольцо регуляции. Управление производится по так называемой открытой петле, а активность во многих произвольно сокращающихся мышцах возникает раньше, чем регистрируется обратная афферентная импульсация. Например, при выполнении прыжковых движений электрическая активность в мышцах, направленная на амортизацию удара, возникает раньше, чем происходит соприкосновение с опорой, т. е. она носит предупредительный характер.Такие центральные программы создаются согласно сформированному в мозге (главным образом -- в ассоциативной переднелобной области коры) образу двигательного действия и цели движения. В дальнейшей конкретной разработке моторной программы принимают участие мозжечок (латеральная область его коры) и базальные ядра (полосатое тело и бледное ядро). Информация от них поступает через таламус в моторную и премоторную области коры и далее -- к исполнительным центрам спинного мозга и скелетным мышцам.Механизм кольцевого регулирования является более древним филогенетически и возникает раньше в процессе индивидуального развития. Примерно к трем годам достаточное развитие получают зрительные обратные связи, осуществляющие текущий зрительно-моторный контроль, а с 5-6 лет происходит переход к текущему контролю движений с участием проприоцептивных обратных связей. Этот механизм достигает значительного совершенства к 7-9 летнему возрасту, после чего начинается переход к формированию механизма центральных команд. К 10-11 годам повышение скорости произвольных движений обеспечивается достаточным развитием процессов предварительного программирования их пространственных и временных параметров. С этого возраста представлены оба механизма управления произвольными движениями, дальнейшее совершенствование которых продолжается вплоть до 17-19лет.
1.3 Три основных функциональных блока мозга
Среди многоэтажных систем нервных центров обобщенно можно выделить три основных функциональных блока (Лурия А. Р., 19 7 3):
1. блок регуляции тонуса, уровня бодрствования;
2. блок приема, переработки и хранения информации;
3. блок программирования, регуляции и контроля двигательной деятельности.
2. Роль различных отделов ЦНС в регуляции позно-тонических реакций
Мышечная деятельность включает в себя процессы осуществления двигательных актов и процессы поддержания позы тела. Эти процессы регулируются различными отделами ЦНС.
2.1 Роль спинного мозга
Мышечный тонус является по своей природе рефлекторным актом. Для его возникновения достаточна рефлекторная деятельность спинного мозга. При длительном растяжении мышц в поле силы тяжести возникает постоянное раздражение их проприорецепторов, потоки импульсов от которых проходят по толстым афферентным волокнам в спинной мозг, где передаются непосредственно (безучастия вставочных нейронов) на альфа-мотонейроны передних рогов и вызывают тоническое напряжение мышц. Такие двухнейронные (или моносинаптические) рефлекторные дуги лежат в основе тонических сухожильных (с рецепторов сухожилий) и миотатических рефлексов на растяжение (с рецепторов мышечных веретен). Это рефлексы активного противодействия мышцы ее растяжению. В произвольной двигательной деятельности человека иногда требуется подавление этих рефлексов, например, при выполнении шпагата. Степень тонического напряжения мышцы зависит от частоты импульсов, посылаемых к ней альфа-мотонейронами. Однако, потоки этих импульсов могут регулироваться вышележащими этажами нервной системы, в частности, неспецифическими отделами ствола мозга с помощью так называемой гамма -- регуляции. Разряды гамма-мотонейронов спинного мозга под влиянием ретикулярной формации повышают чувствительность рецепторов мышечных веретен. В результате при той же длине мышцы увеличивается поток импульсов от рецепторов к альфа-мотонейронам и далее к мышце, повышая ее тонус.
2.2 Роль коры головного мозга, мозжечка и ствола мозга
В регуляции тонуса участвует также медленная часть пирамидной системы и различные структуры экстрапирамидной системы (подкорковые ядра, красные ядра и черная субстанция среднего мозга, мозжечок, ретикулярная формация ствола мозга, вестибулярные ядра продолговатого мозга).Неспецифическая система вызывает общее изменение тонуса различных мышц: усиление тонуса осуществляет активирующий отдел ретикулярной формации среднего мозга, а угнетение -- тормозящий отдел продолговатого мозга. В отличие от нее специфическая система (моторные центры коры больших полушарий и ствола мозга) действует избирательно, т. е. на отдельные группы мышц-сгибателей или разгибателей. Усиление тонуса мышц-сгибателей вызывают корковые влияния, передающиеся непосредственно к нейронам спинного мозга но корково-спинномозговой (пирамидной) системе, а также через красные ядра (по корково-красноядерно-спинномозговой системе) и частично через ретикулярную формацию (по ретикуло-спинномозговой системе). В противоположность им, влияния, передающиеся через вестибулярные ядра продолговатого мозга к вставочным и двигательным нейронам передних рогов спинного мозга (по вестибуло-спинномозговым путям), вызывают тоническое повышение возбудимости мотонейронов мышц-разгибатслей. что обеспечивает повышение тонуса этих мышц. Мозжечок формирует правильное распределение тонуса скелетных мышц: через красные ядра среднего мозга он повышает тонус мышц-сгибателей, а через вестибулярные ядра продолговатого мозга -- усиливает тонус мышц-разгибателей. В поддержании позы и равновесия тела, регуляции тонуса мышц основное значение имеет медиальная продольная зона мозжечка -- кора червя. При мозжечковых расстройствах падает тоническое напряжение мышц (атония) и вследствие ненормального распределения тонуса мышц конечностей возникает нарушение походки (атаксия).Бледное ядро угнетает тонус мышц, а полосатое тело снижаем его угнетающее действие. Высший контроль тонической активности мышц осуществляет кора больших полушарий, в частности ее моторные, премоторные и лобные области. С ее участием происходит выбор наиболее целесообразной для данного момента позы тела, обеспечивается ее соответствие двигательной задаче. Непосредственное отношение к регуляции тонуса мышц имеют медленные пирамидные нейроны положения. Корковые влияния на тонические реакции мышц передаются через медленную часть пирамидного тракта и через экстрапирамидную систему.
3. Рефлексы поддержания позы (установочные)
Специальная группа рефлексов способствует сохранению позы -- это так называемые установочные рефлексы. К ним относятся статические и стато-кинетические рефлексы, в осуществлении которых большое значение имеют продолговатый и средний мозг. Статические рефлексы возникают при изменении положения тела или его частей в пространстве: 1) при изменениях положения головы в пространстве -- лабиринтные рефлексы, возникающие при раздражении рецепторов вестибулярного аппарата. 2) шейные рефлексы - возникающие с проприорецепторов мышц шеи при изменении положения головы по отношению к туловищу, и 3) выпрямительные рефлексы -- с рецепторов кожи, вестибулярного аппарата и сетчатки глаза. Например, при отклонении головы назад повышается тонус мышц-разгибателей спины, а при наклоне вперед -- тонус мышц-сгибателей (лабиринтный рефлекс). С помощью выпрямительного рефлекса происходят последовательные сокращения мышц шеи и туловища, а затем и конечностей. Этот рефлекс обеспечивает вертикальное положение тела теменем кверху. У человека он проявляется, например, при нырянии.
Стато-кинетические рефлексы компенсируют отклонения тела при ускорении и замедлении прямолинейного движения (лифтный рефлекс), а также при вращениях (отклонения головы, тела и глаз в сторону, противоположную движению). Перемещение глаз со скоростью вращения тела, но в противоположную сторону, и быстрое их возвращение в исходное положение -- нистагм глаз -- обеспечивает сохранение изображения внешнего мира на сетчатке глаз и тем самым зрительную ориентацию.
4. Роль различных отделов ЦНС в регуляции движений
Спинной мозг обеспечивает протекание многих элементарных двигательных рефлексов, включение которых в сложные двигательные акты и регуляция по мощности, пространственной ориентации и моменту включения осуществляется вышележащими отделами головного мозга под контролем коры больших полушарий.
4.1 Роль спинного мозга и подкорковых отделов ЦНС в регуляции движений
К управлению ритмическими движениями непосредственное отношение имеют активирующие и угнетающие отделы ретикулярной формации, влияющие на силу и темп сокращения мышц, а также подкорковые ядра, которые организуют автоматическое их протекание и содружественные движения конечностей. Включение древних форм ритмических движений (циклоидных) в акт письма позволяет человеку перейти от отдельного начертания букв к обычной письменной скорописи. То же самое происходит при освоении акта ходьбы -- с переходом от отдельных шагов к ритмической походке. Плавность ритмических движений, четкое чередование реципрокных сокращений мышц обеспечивают премоторные отделы коры.
4.2 Роль различных отделов коры больших полушарий
4.3 Речевая регуляция движений
Спецификой регуляции движений у человека является то, что они подчинены речевым воздействиям, т. е. могут программироваться лобными долями в ответ на поступающие извне словесные сигналы, а также благодаря участию внешней или внутренней речи (мышления) самого человека. В этой функции принимают участие расположенные в левом полушарии человека сенсорный центр речи Вернике и моторный центр речи -- центр Брока. Считают, что афферентная импульсация от речевой мускулатуры является важным ориентиром, дополняющим проприоцептивные сигналы от работающих мышц, а формирующиеся на речевой основе избирательные связи в коре облегчают составление моторных программ.Эта управляющая система еще не развита у ребенка 2-3 лет. Она появляется лишь к 3-4 годам. Внешняя речь, сменяясь постепенно шепотом и переходя затем во внутреннюю речь, становится важным регулятором моторных действий взрослого человека.
4.4 Нисходящие моторные системы
Высшие отделы головного мозга осуществляют свои влияния па деятельность нижележащих отделов, в том числе спинного мозга, через нисходящие пути, которые группируют обычно в две основные нисходящие системы -- пирамидную и экстрапирамидную.В настоящее время предлагают подразделять основные нисходящие пути, исходя из расположения нервных окончаний в спинном мозге и функциональных различий, на следующие 2 системы: более молодую латеральную, волокна которой оканчиваются в боковых (латеральных) частях спинного мозга и связанную преимущественно с мускулатурой дистальных звеньев конечностей (сюда относят корково-спинномозговую и красноядерно-спинномозговую системы), и древнюю медиальную, волокна которой оканчиваются во внутренних (медиальных) частях белого вещества, связанную главным образом с мускулатурой туловища и проксимальных звеньев конечностей, состоящую из вестибуло-спинномозговой и ретикуло-спинномозговой систем.
Пирамидная система выполняет 3 основные функции:
· посылает мотонейропам спинного мозга импульсы -- команды к движениям (пусковые влияния);
· изменяет проведение нервных импульсов во вставочных спинальных нейронах, облегчая протекание нужных в данный момент спинномозговых рефлексов;
· осуществляет контроль за потоками афферентных сигналов в нервные центры, выключая постороннюю информацию и обеспечивая обратные связи от работающих мышц.
Рис. 1. Схема основных нисходящих путей регуляции двигательной деятельности
1 -- быстрая подсистема и 2 -- медленная подсистема корково-спиномозго-вого пути (пирамидного тракта); 3 -- корково-красноядерно-спиномозговой путь. Латеральная система -- I, 2, 3. Медиальная система -- 4, 5. М -- мотонейрон спинного мозга, получающий фазные (Фазн.) и тонические (Тонич.) возбуждающие (+) и тормозящие (-) влияния.
управление движение мозг центральный нервный система
1. Возрастная физиология / Ю.А. Ермалаев. - М.: Высш. шк. 1985. 384 с., ил. 2. Общая психология / Под редак. А.В. Петровского - М., 1986 г., стр. 480
3. Физиология ВНД / Л.Г. Воронин. - М.: Высш. шк. 1979
4. Физиология человека / под. ред. Г.И. Косицкого. - Ф50 3-е изд., перераб. и доп., - М.: Медицина, 1985. 544с., ил.
5. Физиология человека / под. ред. Н. В. Зимкина. - 3-е изд. - М.: Физкультура и спорт. 1964.
Подобные документы
Разновидности движений человека и рецепторы, участвующие в регуляции моторики: проприорецепторы, вестибулярные, зрительные. Рефлексы спинного мозга: сгибательный, перекрестный разгибательный, чесательный и шагательный. Моторные рефлексы ствола мозга.
презентация [1,4 M], добавлен 29.08.2013
Особенности строения ствола головного мозга, физиологическая роль ретикулярной формации мозга. Функции мозжечка и его влияние на состояние рецепторного аппарата. Строение вегетативной нервной системы человека. Методы изучения коры головного мозга.
реферат [1,7 M], добавлен 23.06.2010
Двусторонние связи клеток коры мозжечка. Участие мозжечка в выполнении осознанных (произвольных) движений. Двойной тип влияний клеток Пуркинье. Дифференцировка влияний отдельных структур мозжечка. Зоны коры, участвующие в осуществлении сложных движений.
презентация [652,3 K], добавлен 29.08.2013
Роль нервной системы в регуляция мозгового кровотока. Роль парасимпатической системы в регуляции мозгового кровообращения. Роль ствола мозга в обеспечении адекватного кровотока. Регуляторные контуры: нейрогенный, гуморальный, метаболический и миогенный.
реферат [16,7 K], добавлен 25.04.2009
Структурная и функциональная характеристика мозжечка - отдела головного мозга, отвечающего за координацию движений, регуляцию равновесия и мышечного тонуса. Сравнительная анатомия и эволюция. Полушарии мозжечка, серое и белое вещество, проводящие пути.
презентация [1,3 M], добавлен 27.04.2015
Наследственные и приобретенные нарушения обмена веществ. Метаболические энцефалопатии как расстройства различных отделов ЦНС. Нарушения мозгового кровотока, миелина, нервных механизмов управления движениями и нарушение движений при повреждении мозжечка.
реферат [19,6 K], добавлен 13.04.2009
Общая структура головного мозга человека. Функции его отделов: лобной, теменной, затылочной, височной доли, островка. Развитие мозжечка как структуры в ходе эволюции организмов. Строение больших полушарий головного мозга. Нарушение функций мозжечка.
Произвольные, они же сознательные движения, - это те, которые человек способен контролировать при помощи коры больших полушарий головного мозга. В осуществлении двигательного акта участвует множество уровней периферической и центральной нервной системы. Эти уровни не работают изолированно, они находятся в постоянной взаимосвязи, передавая друг другу нервные импульсы. Что же обеспечивает произвольные движения человека? Об этом подробно в статье.

Значение афферентных сигналов
Вам будет интересно: Пополняем словарный запас: тирада — это.
Главная роль в осуществлении произвольных движений человека ложится на афферентные сигналы. Это импульсы, которые поступают к телу человека извне. Перед тем, как осуществить какое-либо движение, нервный сигнал улавливается рецепторами и через сенсорные нервные пути поступает к структурам центральной нервной системы. Через эти пути головной мозг узнает, что скелетные мышцы готовы к выполнению движения.
Афферентные импульсы выполняют такие функции:
- сообщают коре головного мозга о том, что есть необходимость выполнить движение;
- "говорят", правильно ли оно выполняется;
- повышают или, наоборот, снижают силу сокращения мышечных волокон;
- корригируют последовательность сокращения мышечной ткани;
- информируют кору о том, стоит ли прекратить двигательную активность или есть необходимость в ее продолжении.
Две зоны коры - моторная и чувствительная - составляют единое целое сенсомоторного отдела. Он контролирует работу нижележащих структур головного и спинного мозга при обеспечении произвольных движений человека.
Вам будет интересно: Что такое ливрея? Значение слова
Двигательные центры
Центры системы движений человека в коре головного мозга расположены в прецентральной извилине. Она размещена перед центральной бороздой в лобном отделе коры. Этот отдел вместе с парацентральной долькой и небольшим участком лобной доли называют первичным двигательным проекционным полем.
Вторичное поле размещено в премоторном участке коры. Именно за счет первых двух полей происходит реализация запланированного двигательного акта.
Произвольные движения человека интегрируются в третичном поле, которое расположено в передних частях лобной доли. Благодаря работе этого участка коры двигательный акт точно соответствует поступившей сенсорной информации.
Все процессы, которые происходят в человеческом организме, интегрируются двумя отделами нервной системы: автономным и соматическим. Именно автономная нервная система человека управляет произвольными движениями.
Пирамидные клетки
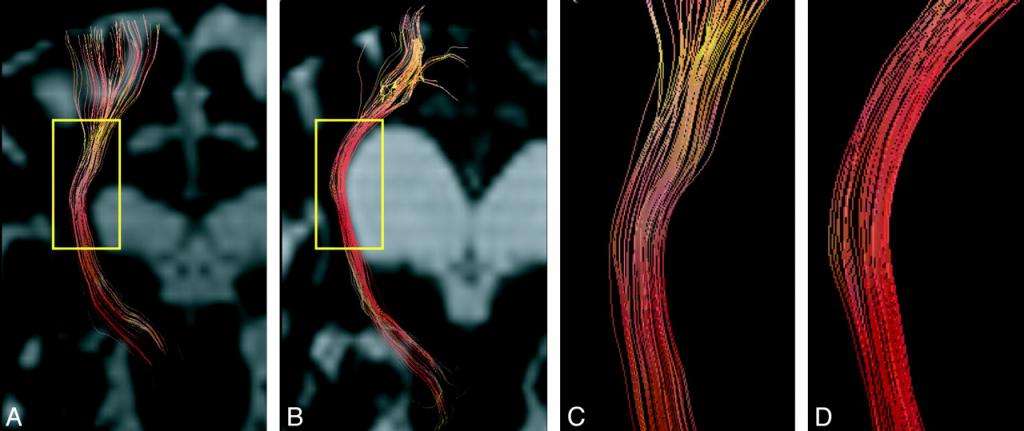
В области первичного и вторичного двигательных полей в пятом слое серого вещества головного мозга расположены гигантские пирамидные клетки. Эти образования открыл ученый В. А. Бец, поэтому их также называют в его честь - клетки Беца. С этих клеток начинается длинный пирамидный путь. Он, взаимодействуя с нервными волокнами периферической нервной системы и поперечно-полосатой мышечной тканью, дает нам возможность двигаться по собственной воле.
Элементы корково-мышечного пути
Произвольные движения человека обеспечивает в первую очередь корково-мышечный, или пирамидный путь. В состав этого образования входят два нейрона. Один из них получил название центрального, второй - периферического.
Центральный нейрон представляет собой тело пирамидной клетки Беца, от которой отходит длинный отросток (аксон). Этот аксон спускается к передним рогам спинного мозга, где передает нервный импульс на второй нейрон. От тела второй нервной клетки также отходит длинный отросток, который идет на периферию и передает информацию к скелетным мышцам, заставляя их двигаться. Именно так осуществляется движение туловища и конечностей.
Но что же о мышцах лица? Чтобы их произвольные сокращения были возможными, часть аксонов центральных нервных клеток идет не к спинному мозгу, а к ядрам черепных нервов. Эти образования находятся в продолговатом спинном мозге. Именно они являются вторыми двигательными нейронами для мышц лица.
Таким образом, пирамидный путь состоит из двух частей:
- корково-спинномозговой путь, который передает импульсы нейронам спинного мозга;
- корково-ядерный путь, идущий к продолговатому мозгу.
Осуществление движений туловища
Отростки центральных нейронов сначала размещены под корой. Здесь они радиально расходятся в виде лучистого венца. Далее они подходят ближе друг к другу и располагаются на колене и задней ножке внутренней капсулы. Это структура в полушариях головного мозга, которая расположена между таламусом и базальными ганглиями.
Потом волокна подходят через ножки мозга к продолговатому мозгу. На передней поверхности этой структуры пирамидные пути образуют две выпуклости - пирамиды. В месте, где продолговатый мозг переходит в спинной, происходит перекрест части нервных волокон.
Перекрещенная часть далее идет в составе бокового канатика, неперекрещенная входит в состав переднего канатика спинного мозга. Так образуются боковой и передний корково-спинномозговые пути соответственно. Волокна этих путей постепенно становятся тоньше и в итоге заканчиваются на ядрах передних рогов спинного мозга. Они передают импульсы на альфа-мотонейроны, находящиеся в этом участке.
При этом волокна переднего пути делают перекрест в спинном мозге на его передней спайке. То есть весь корково-спинномозговой пути заканчивается с противоположной стороны.
Длинные отростки альфа-мотонейронов выходят из спинного мозга, находясь в составе корешков. После они включаются в нервные сплетения и периферические нервы, неся импульс к скелетным мышцам. Таким образом, мышцы обеспечивают произвольные движения человека за счет импульса, полученного от пирамидных клеток коры головного мозга.
Осуществление движений лица
Часть отростков первых нейронов пирамидного пути не спускается на спинной мозг, а заканчивается на уровне продолговатого мозга. Так образуется корково-ядерный путь. За счет него нервный импульс передается от пирамидных клеток к ядрам черепных нервов.
Эти волокна также частично перекрещиваются на уровне продолговатого мозга. Но есть и отростки, которые осуществляют полный перекрест. Они идут к нижнему отделу ядра лицевого нерва, а также к ядру подъязычного нерва. Такой неполный перекрест означает то, что мышечная ткань, обеспечивающая произвольные движения человека на уровне лица, получает иннервацию сразу с двух сторон коры.
Благодаря такой особенности поражение коры головного мозга с одной стороны вызывает обездвиживание только нижней части лица, а моторная активность верхней полностью сохраняется.
Симптомы повреждения двигательного пути
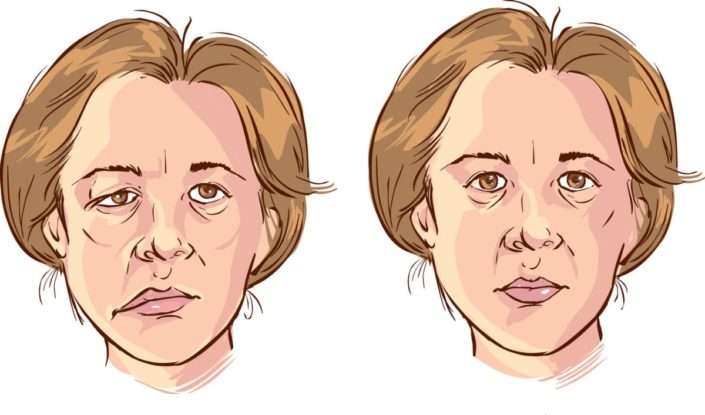
Произвольные движения человека обеспечивают, в первую очередь, кора и пирамидный путь. Поэтому повреждение этих участков при ухудшении кровообращения головного мозга (инсульте), травме или опухоли приводит к нарушению двигательной активности человека.
На каком бы уровне не произошло поражение, мышцы перестают получать импульс от коры, что приводит к полной неспособности осуществить действие. Такой симптом получил название паралича. Если повреждение частично, наблюдается слабость мышц и трудность в осуществлении движений - парез.
Виды параличей
Выделяют два основных вида обездвиживания человека:
- центральный паралич;
- периферический паралич.
Они получили свое название от вида пораженных нейронов. При центральном параличе происходит повреждение первого нейрона. При периферическом обездвиживании поражается, соответственно, периферическая нервная клетка.
Определить вид повреждения можно уже при первом обследовании больного, без дополнительных инструментальных методов. Для центрального паралича характерны такие особенности:
- повышение мышечного тонуса, или гипертония;
- увеличение амплитуды сухожильных рефлексов, или гиперрефлексия;
- уменьшение активности брюшных рефлексов;
- появление патологических рефлексов.
Симптомы периферического паралича являются полной противоположность проявлениям центрального:
- Для учеников 1-11 классов и дошкольников
- Бесплатные сертификаты учителям и участникам
Нарушения произвольных движений и действий относятся к сложным двигательным расстройствам, которые в первую очередь связаны с поражением коркового уровня двигательных функциональных систем.
Этот тип нарушений двигательных функций получил в неврологии и нейропсихологии название апракс и й. Под апраксиями понимаются такие нарушения произвольных движений и действий, которые не сопровождаются четкими элементарными двигательными расстройствами - параличами и парезами, явными нарушениями мышечного тонуса и тремора, хотя возможны сочетания сложных и элементарных двигательных расстройств. Апраксиями обозначают прежде всего нарушения произвольных движений и действий, совершаемых с предметами.
При усилении зрительного контроля движения можно в определенной степени скомпенсировать. При поражении левого полушария кинестетическая апраксия обычно носит двухсторонний характер, при поражении правого полушария - чаще проявляется только в одной левой руке.
Эта форма апраксии изучалась рядом авторов - К. Кляйстом, О. Ферстером и др. Особенно подробно она исследовалась А. Р. Лурия, установившим при данной форме апраксии общность нарушений двигательных функций руки и речевого аппарата в виде первичных трудностей автоматизации движений, выработки двигательных навыков. Кинетическая апраксия проявляется в нарушении самых различных двигательных актов: предметных действий, рисования, письма, - в трудности выполнения графических проб, особенно при серийной организации движений (динамическая апраксия). При поражении нижне-премоторных отделов коры левого полушария (у правшей) кинетическая апраксия наблюдается, как правило, в обеих руках.
Четвертая форма апраксии - регуляторная или префронтальная апраксия - возникает при поражении конвекситальной префронтальной коры кпереди от премоторных отделов; протекает на фоне почти полной сохранности тонуса и мышечной силы. Она проявляется в виде нарушений программирования движений, отключения сознательного контроля за их выполнением, замены нужных движений моторными шаблонами и стереотипами. При грубом распаде произвольной регуляции движений у больных наблюдаются симптомы эхопраксии в виде бесконтрольных подражательных повторений движений экспериментатора. При массивных поражениях левой лобной доли (у правшей) наряду с эхопраксией возникает эхолалия - подражательные повторения услышанных слов или фраз.
Для регуляторной апраксии характерны системные персеверации, т. е. персеверации всей двигательной программы в целом, а не ее отдельных элементов. Такие больные после письма под диктовку на предложение нарисовать треугольник обводят контур треугольника движениями, характерными для письма, и т. п. Наибольшие трудности у этих больных вызывает смена программ движений и действий. В основе этого дефекта лежит нарушение произвольного контроля за осуществлением движения, нарушение речевой регуляции двигательных актов. Данная форма апраксии наиболее демонстративно проявляется при поражении левой префронтальной области мозга у правшей.
Классификация апраксии, созданная А. Р. Лурия, основана главным образом на анализе нарушений двигательных функций у больных с поражением левого полушария головного мозга. В меньшей степени исследованы формы нарушения произвольных движений и действий при поражении различных корковых зон правого полушария; это - одна из актуальных задач современной нейропсихологии.
6.1. Основные принципы организации движений
Выполнение двигательных актов осуществляется обширным комплексом нейронов, расположенных в различных отделах ЦНС. Такая функциональная система управления движениями является многоэтажной и многоуровневой.
6.1.1. Общая схема управления движениями
Решающим фактором поведения является полезный результат. Для его достижения в нервной системе формируется группа взаимосвязанных нейронов — функциональная система (П. К.Анохин, 1975). Деятельность ее включает следующие процессы: 1) обработка всех сигналов, поступающих из внешней и внутренней среды организма — так называемый афферентный синтез, 2) принятие решения о цели и задачах действия; 3) создание представления об ожидаемом результате и формирование конкретной программы движений; 4) анализ полученного результата и внесение в программу поправок — сенсорных коррекций.
6.1.2. Рефлекторное кольцевое регулирование и программное управление движениями
В двигательной деятельности человека различают произвольные движения —сознательно управляемые целенаправленные действия и непроизвольные движения, происходящие без участия сознания и представляющие собой либо безусловные реакции, либо автоматизированные двигательные навыки. В основе управления произвольными движениями человека лежат два различных физиологических механизма: 1) рефлекторное кольцевое регулирование и 2) программное управление по механизму центральных команд.
Замкнутая система рефлекторного кольцевого регулирования характерна для осуществления различных форм двигательных действий и позных реакций, не требующих быстрого двигательного акта. Это позволяет нервным центрам получать информацию о состоянии мышц и результатах их действий по различным афферентным путям и вносить поправки в моторные команды по ходу действия.
Программное управление по механизму центральных команд — это механизм регуляции движений, независимый от афферентных проприоцептивных влияний. Такое управление используется в случае выполнения кратковременных движений (прыжков, бросков, ударов, метаний), когда организм не успевает использовать информацию от проприорецепторов мышц и других рецепторов. Вся программа должна быть готова еще до начала двигательного акта. При этом отсутствует замкнутое кольцо регуляции. Управление производится по так называемой открытой петле, а активность во многих произвольно сокращающихся мышцах возникает раньше, чем регистрируется обратная афферентная импульсация. Например, при выполнении прыжковых движений электрическая активность в мышцах, направленная на амортизацию удара, возникает раньше, чем происходит соприкосновение с опорой, т. е. она носит предупредительный характер.
Такие центральные программы создаются согласно сформированному в мозге (главным образом — в ассоциативной передне-лобной области коры) образу двигательного действия и цели движения. В дальнейшей конкретной разработке моторной программы принимают участие мозжечок (латеральная область его коры) и базальные ядра (полосатое тело и бледное ядро). Информация от них поступает через таламус в моторную и премоторную области коры и далее — к исполнительным центрам спинного мозга и скелетным мышцам.
Механизм кольцевого регулирования является более древним филогенетически и возникает раньше в процессе индивидуального развития. Примерно к трем годам достаточное развитие получают зрительные обратные связи, осуществляющие текущий зрительно-моторный контроль, ас 5-6 лет происходит переход к текущему контролю движений с участием проприоцептивных обратных связей. Этот механизм достигает значительного совершенства к 7-9 летнему возрасту, после чего начинается переход к формированию механизма центральных команд. К 10-11 годам повышение скорости произвольных движений обеспечивается достаточным развитием процессов предварительного программирования их пространственных и временных параметров. С этого возраста представлены оба механизма управления произвольными движениями, дальнейшее совершенствование которых продолжается вплоть до 17-19 лет.
Читайте также: