
Основы репродукции клеток реферат
Обновлено: 02.07.2024
Возрстные изменения. Соединительные ткани с возрастом претерпевают изменения в строении, количестве и химическом составе. С возрастом увеличиваются общая масса соединительнотканных образований, рост костного скелета. Во многих разновидностях соединительнотканных структур изменяется соотношение типов коллагена, гликозаминогликанов; в частности, в них становится больше сульфатированных соединений.
№ 4 Репродукция клеток и ее биологическое значение. Способы репродукции. Митотический цикл: периоды, их структурно-функциональная характеристика, чувствительность клеток и воздействие эндогенных и экзогенных факторов. Особенности репродукции половых клеток.
Репродукция клеток.Один из постулатов клеточной теории гласит, что увеличение числа клеток, их размножение происходят путем деления исходной клетки. Делению клеток предшествует редупликация их хромосомного аппарата, синтез ДНК. Это правило является общим для прокариотических и эукариотических клеток. Времясуществования клетки как таковой, от деления до деления или от деления до смерти, обычно называют клеточным циклом.
Способы репродукции.Репродукция половых клеток осуществляется мейозом – спермато- и овогенезом.
Митоз (непрямое деление клетки) - это такое деление клеточного ядра, при котором образуется два дочерних ядра, каждое из которых содержит набор хромосом идентичный набору материнского ядра. Митоз входит в состав клеточного цикла, т.е. периода между появлением клетки и её делением. Клеточный цикл состоит из интерфазы, митотического (пролиферативного) цикла и цитокинеза. Во время интерфазы клетка подготавливается к делению: происходит деление митохондрий, сборка рибосом, редупликация ДНК, хроматин диспирализуется и становится виден в световой микроскоп.
Биологическое значение митотического цикла состоит в том, что он обеспечивает образование клеток, равноценных по объему и содержанию наследственной информации. В митотическом цикле выделяют репродуктивную (интерфаза) и разделительную (митоз) фазы.
В начальной отрезок интерфазы (постмитотический, пресинтетический или G1-период) восстанавливаются черты организации интерфазной клетки, завершается формирование ядрышка, начавшееся в телофазе. Из цитоплазмы в ядро поступает значительное (90%) количество белка. Масса клетки возрастает за счет интенсивного синтеза белка в цитоплазме.
В синтетическом (S-период) удваивается количество наследственного материала клетки. Наряду с ДНК, интенсивно образуются РНК и белок, а количество гистонов строго удваивается. Продолжительность синтетического периода в клетках человека составляет 7-12 ч.
Постсинтетический (предмитотический или G2-период) занимает отрезок времени от окончания синтетического периода до начала митоза. Он характеризуется интенсивным синтезом РНК и белка, завершается увеличение массы цитоплазмы.
Митоз делят на четыре фазы: профаза, метафаза, анафаза, телофаза. Обязательным для разделительной фазы митоза является построение митотического аппарата, который состоит из системы микротрубочек (ахроматиновое веретено, или веретено деления) и структур, поляризующих митоз, т.е. обозначающих два полюса в клетке, к которым разойдутся дочерние хромосомы. Митотический аппарат обеспечивает направленное перемещение дочерних хромосом в анафазе.
Особенности репродукции половых клеток. Мужские и женские клетки несутединичный (гаплоидный) набор хромосом и, следовательно, содержат ДНК в 2 раза меньше, чем все остальные клетки организма. Такие половые клетки (сперматозоиды и ооциты) с единичным набором хромосом называют гаплоидными. Плоидность обозначают буквой n. Соответственно количество ДНКна клетку (с) зависит от ее плоидности. При оплодотворении происходит слияние двух клеток, каждая из которых несет 1 n набор хромосом, поэтому образуется диплоидная (2 п, 2 с) клетка-зигота. В дальнейшем в результате деления диплоидной зиготы и последующего деления диплоидных клеток разовьется организм, клетки которого (кроме зрелых половых) будут диплоидны.
№ 11 Физиологическая и репаративная регенерация. Структурные основы регенераторных возможностей различных органов и тканей.
Физиологическая регенерация – восстановление организмом утраченных или поврежденных органов или тканей.
Репаративная регенерация –восстановление какой – либо ткани в патологических условиях.
Эпителиальная ткань:
Регенерация. Покровный эпителий постоянно испытывает влияние внешней среды, поэтому эпителиальные клетки сравнительно быстро изнашиваются и погибают. Источником их восстановления являются стволовые клетки эпителия. Они сохраняют способность к делению в течение всей жизни организма. Размножаясь, часть вновь образованных клеток вступает в дифференцировку и превращается в эпителиоциты, подобные утраченным. Высокая способность эпителия к физиологической регенерации служит основой для быстрого восстановления его в патологических условиях (репаративная регенерация).
С возрастом в покровном эпителии наблюдается ослабление процессов обновления.
Соединительная ткань:
Собственная соединительная ткань
Регенерация.Физиологическая регенерация хрящевой ткани осуществляется за счет малоспециализированных клеток надхрящницы и хряща путем размножения и дифференцировки прехондробластов и хондробластов. Однако этот процесс идет очень медленно. Посттравматическая регенерация хрящевой ткани внесуставной локализации осуществляется за счет надхрящницы. Репарация может происходить за счет клеток окружающей соединительной ткани, не потерявших способности к метаплазии.
Регенерация.Физиологическая регенерация костных тканей происходит медленно за счет остеогенных клеток надкостницы, эндоста и остеогенных клеток в канале остеона. Посттравматическая регенерация костной ткани протекает лучше в тех случаях, когда концы сломанной кости не смещены относительно друг друга. Процессу остеогенеза предшествует формирование соединительнотканной мозоли, в толще которой могут образовываться хрящевые отростки. Оссификация в этом случае идет по типу вторичного (непрямого) остеогенеза. Но прежде чем начнут строить кость остеобласты, остеокласты образуют небольшую щель между репонированными концами кости.
Мышечная ткань:
Регенерацияскелетной мышечной ткани:
Ядра миосимиластов делиться не могут, так как у них отсутствуют клеточные центры. Камбиальными элементами служат миосателлитоциты. Пока организм растет, они делятся, а дочерние клетки встраиваются в концы симпластов. По окончании роста размножение миосателлитоцитов затухает. После повреждения мышечного волокна на некотором протяжении от места травмы оно разрушается и егофрагменты фагоцитируются макрофагами. Восстановление тканей осуни ляется за счет двух механизмов: компенсаторной гипертрофии самого симпласта и пролиферации миосателлитоцитов. Миосателлитоциты образуют миотубы, которые входят в состав вновь образованных мышечных волокон или формируют новые волокна.
Возможности регенерации сердечной мышечной ткани: Стволовых клеток в сердечной мышце нет, поэтому погибающие кардиомиоциты не восстанавливаются.
Нервная ткань:
Регенерация зависит от места травмы. В ЦНС и в периферической нервной системе погибшие нейроны не восстанавливаются. Полноценной регенерации нервных волокон в составе ЦНС обычно не происходит, но нервные волокна в составе периферических нервов обычно хорошо регенерируют. При этом нейролеммоциты периферического отрезка и ближайшего к области травмы участка пролиферируют.
Поврежденные нервные волокна головного и спинного мозга не регенерируют.
№ 23 Рыхлая волокнистая соединительная ткань. Морфо-функциональная характеристика. Межклеточное вещество, строение и значение. Фибробласты и их роль в образовании межклеточного вещества.
Рыхлая волокнистая соединительная ткань обнаруживается во всех органах, так как она сопровождает крс-веносные и лимфатические сосуды и образует строму многих органов. Не смотря на наличие органных особенностей, строение рыхлой волокнистой соединительной ткани в различных органах имеет сходство. Она состоит из клеток и межклеточного вещества.
Межклеточное вещество, или матрикс, соединительной ткани состоит из коллагеновых и эластических волокон, а также из основного (аморфного) вещества. Межклеточное вещество как у зародышей, так и у взрослых образуется, с одной стороны, путем секреции, осуществляемой соединительнотканными клетками, а с другой — из плазмы крови, поступающей в межклеточные пространства.
У зародышей человека образование межклеточного вещества происходит начиная с 1—2-го месяца внутриутробного развития. В течение жизни межклеточное вещество постоянно обновляется — резорбируется и восстанавливается.
Коллагеновые структуры, входящие в состав соединительных тканей организмов человека и животных, являются наиболее представительными ее компонентами, образующими сложную организационную иерархию. Основу всей группы коллагеновых структур составляет волокнистый белок — коллаген, который определяет свойства коллагеновых структр.
Коллагеновые волокна в составе разных видов соединительной ткани определяют их прочность. В рыхлой неоформленной волокнистой соединительной ткани они располагаются в различных направлениях в виде волнообразно изогнутых, спиралевидно скрученных, округлых или уплощенных в сечении тяжей. Внутренняя структура коллагенового волокна определяется фибриллярным белком — коллагеном, который синтезируется на рибосомах гранулярной эндоплазматической сети фибробластов.
Различают 14 типов коллагена, отличающихся молекулярной организацией, органной и тканевой принадлежностью.
Эластические волокна.Наличие эластических волокон в соединительной ткани определяет ее эластичность и растяжимость. В рыхлой волокнистой соединительной ткани они широко анастомозируют друг с другом. В составе эластических волокон различают микрофибриллярный и аморфный компоненты.
Основой эластических волокон является глобулярный гликопротеин — эластин, синтезируемый фибробластами и гладкими мышечными клетками.
Фибробласты (фибробластоциты) — клетки, синтезирующие компоненты межклеточного вещества: белки (коллаген, эластин), протеогликаны, гликопротеины.
Среди мезенхимных клеток имеются стволовые клетки, дающие начало дифферону фибробластов: стволовые клетки, полустволовые клетки-предшественники, малоспециализированные, дифференцированные фибробласты (зрелые, активно функционирующие), фиброциты (дефинитивные формы клеток), а также миофибробласты и фиброкласты. С главной функцией фибробластов связаны образование основного вещества и волокон, заживление ран, развитие рубцовой ткани, образование соединительнотканной капсулы вокруг инородного тела и др. Морфологически в этом диффероне можно идентифицировать только клетки, начиная с малоспециализированного фибробласта.
В цитоплазме фибробластов, особенно в периферическом слое, располагаются микрофиламенты, содержащие белки типа актина и миозина, что обусловливает способность этих клеток к движению. Движение фибробластов становится возможным только после их связывания с опорными фибриллярными структурами с помощью фибронектина — гликопротеина, синтезированного фибробластами и другими клетками, обеспечивающего адгезию клеток и неклеточных структур.
№ 24 Рыхлая волокнистая соединительная ткань. Морфо-функциональная характеристика. Макрофаги, строение и источники развития. Понятие о макрофагической системе. Вклад русских ученых в гистофизиологию соединительных тканей.
Рыхлая волокнистая соединительная ткань обнаруживается во всех органах, так как она сопровождает крс-веносные и лимфатические сосуды и образует строму многих органов. Не смотря на наличие органных особенностей, строение рыхлой волокнистой соединительной ткани в различных органах имеет сходство. Она состоит из клеток и межклеточного вещества.
Макрофаги (макрофагоциты) — это гетерогенная специализированная клеточная популяция защитной системы организма. Различают две группы макрофагов — свободные и фиксированные. К свободным макрофагам относятся макрофаги рыхлой соединительной ткани, или гистиоциты; макрофаги серозных полостей; макрофаги воспалительных экссудатов; альвеолярные макрофаги легких. Макрофаги способны перемещаться в организме. Группу фиксированных (резидентных) макрофагов составляют макрофаги костного мозга и костной ткани (остеокласты), селезенки, лимфатических узлов (дендритные макрофаги), внутриэпидермальные макрофаги (клетки Лангерганса), макрофаги ворсин плаценты (клетки Хофбауэра), ЦНС (микроглия).
Обычно макрофаги, за исключением некоторых их видов (гигантские клетки инородных тел, хондро- и остеокласты), имеют одно ядро. В ядрах содержатся крупные глыбки хроматина.
Формы проявления защитной функции макрофагов: 1) поглощение и дальнейшее расщепление или изоляция чужеродного материала; 2) обезвреживание его при непосредственном контакте; 3) передача информации о чужеродном материале иммунокомпетентным клеткам, способным его нейтрализовать; 4) оказание стимулирующего воздействия на другую клеточную популяцию защитной системы организма.
Понятие о макрофагической системе.К этой системе относятся совокупность всех клеток, обладающих способностью захватывать из тканевой жидкости организма инородные частицы, погибающие клетки, неклеточные структуры, бактерии и др. К таким клеткам относятся макрофаги (гистиоциты) рыхлой волокнистой соединительной ткани, звездчатые клетки синусоидных сосудов печени, свободные и фиксированные макрофаги кроветворных органов (костного мозга, селезенки, лимфатических узлов), макрофаги легкого, воспалительных экссудатов (перитонеальные макрофаги), остеокласты, гигантские клетки инородных тел и глиальные макрофаги нервной ткани (микро-глия).
Макрофагическая система представляет собой мощный защитный аппарат, принимающий участие как в общих, так и в местных защитных реакциях организма. В целостном организме макрофагическая система регулируется как местными механизмами, так нервной и эндокринной системами.
Вклад русских учёных. Начатое еще А. А. Максимовым изучение соединительной ткани приобрело широкий размах в советский период. Изучение ведется в основном по двум направлениям. Первое направление в изучении соединительной ткани выражается в широких сравнительно-гистологических исследованиях соединительной ткани и крови (С. В. Мясоедов, А. А. Заварзин, Ф. М. Ла-заренко, Е. С. Данини, Г. В. Ясвоин, Г. К. Хрущев и др.).
Второе направление — изучение гистофизиологии соединительной ткани различных органов и систем, а также ее изменений под влиянием нервных и эндокринных факторов (В. Г. Елисеев, Т. А. Григорьева и др.)- С этими направлениями логически связано изучение гистогенеза соединительной ткани.
№ 25 Взаимодействие иммуннокомпетентных клеток в иммунных реакциях. Понятие о медиаторах иммунного ответа.
Основными клетками, осуществляющими иммунные реакции, являются Т- и В-лимфоциты (и их производные плазмоциты), макрофаги, а также ряд взаимодействующих с ними клеток (тучные клетки, эозинофилы).
Рассмотрение компонентов ядра: кариолеммы, кариоплазмы, хроматина и ядрышек. Этапы клеточного цикла: гетерокаталитическая интерфаза, митотический цикл (автокаталитическая интерфаза) и период относительного покоя. Метафаза, анафаза и телофаза мейоза.
| Рубрика | Биология и естествознание |
| Вид | презентация |
| Язык | русский |
| Дата добавления | 20.09.2014 |
| Размер файла | 4,1 M |
Студенты, аспиранты, молодые ученые, использующие базу знаний в своей учебе и работе, будут вам очень благодарны.
HTML-версии работы пока нет.
Cкачать архив работы можно перейдя по ссылке, которая находятся ниже.
Подобные документы
Изучение процесса митоза как непрямого деления клетки и распространенного способа репродукции эукариотических клеток, его биологическое значение. Мейоз как редукционное деление клетки. Интерфаза, профаза, метафаза, анафаза и телофаза мейоза и митоза.
презентация [7,6 M], добавлен 21.02.2013
Основные фазы клеточного цикла: интерфаза и митоз. Определение понятия "митоз" как непрямого деления клетки, наиболее распространенного способа репродукции эукариотических клеток. Характеристика и особенности процессов деления: амитоза и мейоза.
презентация [799,4 K], добавлен 25.10.2011
Период жизнедеятельности клетки, в котором происходят все обменные процессы и деление. Интерфаза, метафаза и анафаза, деление клетки. Биологический смысл митоза. Вирусы и бактериофаги как неклеточные формы жизни. Виды и формы размножения организмов.
реферат [20,3 K], добавлен 06.07.2010
Клеточный цикл как период существования клетки от момента ее образования путем деления материнской клетки до собственного деления или гибели. Принципы и методы его регуляции. Этапы и биологическое значение митоза, мейоза, обоснование данных процессов.
презентация [1,1 M], добавлен 07.12.2014
Единый план строения клеток организма. Строгая упорядоченность строения ядра и цитоплазмы. Клеточное ядро (вместилище всей генетической информации). Содержимое клеточного ядра (хроматин). Аппарат Гольджи, эндоплазматическая сеть, клеточные структуры.
реферат [21,6 K], добавлен 28.07.2009
Эволюционное значение клеточного ядра - компонента эукариотической клетки, содержащего генетическую информацию. Структура ядра: хроматин, ядрышко, кариоплазма и ядерная оболочка. Функции ядра: хранение, передача и реализация наследственной информации.
презентация [3,1 M], добавлен 21.02.2014
Теория кометного происхождения органических молекул. Этапы происхождения жизни по Опарину. Первые живые организмы на Земле. Обособление клеточного ядра. Эволюционная схема происхождения ядра профессора А.Н. Мосолова. Этапы ранней эволюции жизни на Земле.
Содержание
Введение
Содержание
Размножение клетки
Бесполое размножение клетки
Половое размножение клетки
Репродукция клетки
Жизненный цикл клетки
Способы репродукции
Апоптоз
Заключение
Список использованной литературы
Прикрепленные файлы: 1 файл
Реферат Цитологические основы размножения. Репродукция клеток..docx
По предмету Биология
«Цитогенетические основы размножения.
- Введение
- Содержание
- Размножение клетки
- Бесполое размножение клетки
- Половое размножение клетки
- Репродукция клетки
- Жизненный цикл клетки
- Способы репродукции
- Апоптоз
- Заключение
- Список использованной литературы
В данном реферате мы рассмотрим более подробно основы размножения и репродукцию клеток.
Размножение - способность производить себе подобных особей.
Функции размножение: 1) Продолжение жизни, 2) Преемственность поколений, 3) Сохранение видов.
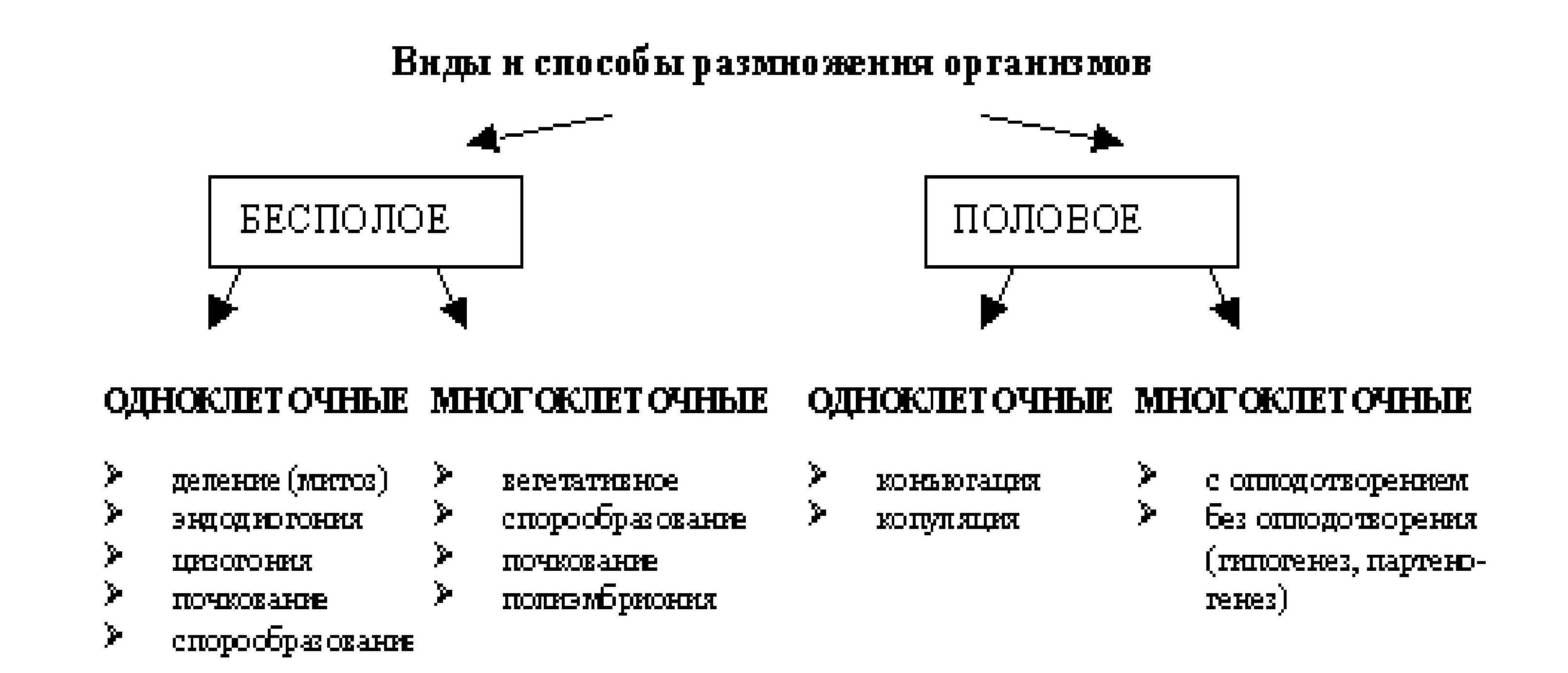
Целостный организм состоит из дискретных единиц - клеток. Существование вида поддерживается размножением. Известны две формы размножения: бесполое и половое. При бесполом размножении организм возникает из соматических клеток, и источником изменчивости могут быть случайные мутации. При половом размножении необходимо, как правило, наличие двух особей, и новый организм возникает из специализированных половых клеток или особей, выполняющих эти функции. В основе полового размножения лежит половой процесс - объединение в наследственном материале генетической информации из разных источников - родителей - для развития потомка. Организмам свойственна двойственная наследственность.
Преимущество полового размножения состоит в перекомбинации лучших наследственных признаков обоих родителей, что является источником изменчивости. Потомство более жизнеспособно и приспособлено к условиям существования. Быстрее происходит эволюция.
В бесполом размножении принимает участие одна родительская особь. В результате бесполого размножения можно получить большое количество особей, подобных материнской.
Бесполое размножение широко распространено среди растений, простейших и некоторых многоклеточных.
Основные формы бесполого размножения - деление на два, множественное деление - шизогония, почкование, фрагментация, спорообразование, вегетативное размножение у растений.
Деление на две сопровождается митозом (например, амеба, эвглена), в результате образуются две идентичные особи (рис. 1).
Рис. 1. Размножение зеленой эвглены: 1 - деление, 2, 3 - деление в цисте.
При множественном делении - шизогонии - ядро исходной клетки несколько раз делится митозом, а затем имеет место деление цитоплазмы. Шизогония встречается у малярийного плазмодия.
При почковании новая особь образуется в виде выроста (почки) на родительской особи, а затем отделяется от нее, превращаясь в самостоятельный организм, идентичный родительскому. Почкование характерно для дрожжей (рис.2), кишечнополостных.
Рис. 2. Почкование дрожжевых грибов.
Спорообразование характерно для водорослей, грибов, папоротников, мхов.
Спора - одна из стадий жизненного цикла, служащая для размножения, - состоит из клетки с ядром и цитоплазмой, покрытой оболочкой. Споры гаплоидны, образуются в результате мейоза в больших количествах. Они способны разноситься на далекие расстояния, имеют приспособления для полета и для защиты от неблагоприятных условий. У одноклеточных и нитчатых водорослей клетки, образующие споры, делятся на 2 и образуют зооспоры - споры, имеющие жгутики. Они способны плавать в воде.
У мхов и папоротников спорообразование чередуется с половым
размножением. Споры образуются в спорангиях. При вегетативном размножении от растения отделяется дифференцированная часть, способная развиться в самостоятельное растение. Вегетативное размножение может быть естественным и искусственным
Для естественного вегетативного размножения могут служить специальные образования, имеющие запас питательных веществ: клубни (картофель), луковицы (лук, тюльпан), клубнелуковицы (шафран), корневища (осот, ландыш), усы (земляника).
Для искусственного вегетативного размножения используют части самого растения: черенки (часть стебля), листья (бегония), отводки (ветви пригибают к земле и на них образуются придаточные корни (например, у малины), корневые отпрыски, производят деление кустов, прививки. Прививка состоит в пересадке одного растения (побега или почки) на нижнюю часть другого побега. Пересаженную часть растения - донора называют привоем, а реципиента - подвоем. Получившееся растение имеет корневую систему подвоя, а побег - привоя.
Вегетативное размножение применяют для получения большого числа однородных особей, для закрепления признаков хорошего сорта.
Бесполое размножение может происходить у млекопитающих и человека. Это полиэмбриония - бесполое размножение зародыша на ранних стадиях эмбриогенеза животных, размножающихся половым путем.
Это характерно для броненосцев, у которых на стадии бластулы происходит разделение на 4-8 зародышей.
В результате полиэмбрионии у человека рождаются идентичные близнецы.
Сущность бесполого размножения: 1) Участвует один родитель, 2) новый организм развивается из соматических клеток, 3) Клетки длятся митозом,
3) наследственный материал передается через соматические клетки, 4) дочерние клетки идентичны по генотипу и фенотипу.
При половом размножении в результате оплодотворения две клетки разного пола — гаметы — объединяются в одну клетку, зиготу, из которой затем развивается новая особь.
Поскольку при оплодотворении сливаются и ядра клеток, число хромосом удваивается. Поэтому на каком-то последующем этапе, но не позже образования новых гамет хромосомный набор должен снова уменьшиться наполовину путем мейоза. Таким образом, цикл полового размножения включает чередование диплофазы с двойным и гаплофазы с одиночным набором хромосом, т. е. связан со сменой ядерных фаз.
Половое размножение приводит к образованию новых комбинаций наследственных задатков, полученных от двух родителей, и тем самым к наследственной изменчивости потомства, которая является важным фактором эволюции.
У водорослей и многих грибов одиночные или множественные гаметы образуются в одноклеточных футлярах — гаметангиях. При оогамии в оогониях формируется небольшое число яиц (чаще всего одно яйцо), в спермангиях — множество сперматозоидов. У мхов и папоротников футляры с гаметами многоклеточны: в архегониях содержится по одной яйцеклетке, в антеридиях — по многу сперматозоидов. У семенных растений в цикле смены поколений гаметангии редуцированы
Гаметы у многоклеточных животных (за исключением губок) образуются в особых органах — гонадах (половых железах): яйца — в яичнике, спермин— в семеннике.
Диплоидные клетки, из которых развиваются гаметы, называют оогониями и сперматогониями. Их быстрая пролиферация путем митоза (фаза размножения) в случае человеческих оогониев заканчивается уже в начале третьего года жизни и дает свыше 400 ООО клеток (ооцитов), большинство из которых, впрочем, опять сливаются. Затем клетки растут (фаза роста), причем так называемые ооциты 1-го порядка достигают значительно больших размеров, чем сперматоциты 1-го порядка.
Затем одно за другим происходят два деления созревания, сначала мейотическое (редукционное), а потом митотическое (эквационное) или в обратном порядке; первый случай называют прередукцией, а второй — постредукцией.
В результате делений созревания образуются четыре гаплоидные клетки. Будущие мужские гаметы (сперматиды) одинаковы по размерам, а у особей женского пола продукты деления созревания неравноценны: ооцит 1-го порядка, отделяя маленькое направительное, или полярное, тельце, превращается в ооцит 2-го порядка, а тот в свою очередь отделяет еще одно полярное тельце и становится крупным, богатым цитоплазмой зрелым яйцом, причем одновременно первое направительное тельце тоже может разделиться (хотя чаще этого не происходит). Образовавшиеся два или три полярных тельца в дальнейшем развитии не участвуют.
После делений созревания яйцо готово к оплодотворению. Сперматиды, напротив, еще должны в процессе спермиогенеза морфологически преобразоваться в подвижные спермии (сперматозоиды).
Процесс оплодотворения состоит в слиянии цитоплазмы гамет (плазмогамия) и происходящем, как правило, сразу же после этого слиянии ядер гамет в диплоидный синкарион (кариогамия или амфимиксис). Особые вещества, выделяемые гаметами (гамоны), привлекают клетку-партнера.
Оплодотворение у растений при изо- и анизогамии происходит в воде (у многих водорослей и грибов), при оогамии это бывает редко (например, у бурой водоросли Fucus). Для оплодотворения сперматозоиды проникают в оогонии или архегонии. У семенных растений, за исключением немногих голосеменных (гинкго, саговники), вместо жгутиковых сперматозоидов формируются голые сперматические клетки, которые прорастают к яйцеклетке, образуя пыльцевую трубку
Для оплодотворения у многоклеточных животных свойственна оогамия. До оплодотворения яйцеклетка неактивна, и лишь внедрение спермия активирует ее (побуждает к развитию). Активация и амфимиксис не связаны жестко между собой: в некоторых случаях после "активации яйцеклетки мужское ядро растворяется (псевдогамия).
Яйцо выделяет гиногамон I, который привлекает и активирует' спермин. Это антагонист андрогамона I, который выделяется спермиями и тормозит движение спермиев своего вида и чужих (предохраняет их от преждевременной растраты энергии).
После внедрения головки и шейки сперматозоида (хвост чаще всего остается приклеенным к оболочке яйца) появляется оболочка оплодотворения, препятствующая внедрению других спермиев. Затем, как правило, оба ядра (мужское перед этим увеличивается в объеме) движутся друг к другу и сливаются. Центриоль, содержащаяся в уже отделившейся шейке, делится на две дочерние центриоли, которые участвуют в начинающемся вскоре делении яйца. Центросома яйцеклетки дегенерирует или остается неактивной.
Партеногенез (девственное размножение)— это развитие организма из неоплодотворенного яйца. При диплоидном партеногенезе (у тлей, дафний, коловраток, одуванчика (Taraxacum), ястребинок (Hieracium)) мейоза не происходит и развитие начинается с диплоидных ооцитов 1-го или 2-го порядка. При гаплоидном партеногенезе развитие начинается с гаплоидной яйцеклетки. Возникающие организмы при этом либо гаплоидны (самцы пчел — трутни), либо вследствие процесса, компенсирующего редукцию числа хромосом, диплоидны; для этого или яйцо сливается с одним из полярных телец (у рачка артемии), или хромосомы удваиваются без последующего разделения ядра и клетки, или же после первого деления оба ядра снова сливаются.
Сущность полового размножения: 1) участвуют два родителя, 2) в организме родителей образуются половые клетки – гаметы (яйцеклетки и сперматозоиды), 3) гаметы содержат разнородный материал, 4) новый организм образуется при слиянии яйцеклетки и сперматозоида, 5) дочерние организмы разнообразны по фенотипу и генотипу.
Цитогенетические основы размножения. Репродукция клеток
I. Способы размножения организмов, их сущность
Размножение организмов - это воспроизведение себе подобных. Размножение обеспечивает продолжение жизни, преемственность поколений и сохранение вида. Размножение может быть бесполым и половым.
При бесполом размножении происходит деление соматических клеток одной родительской особи. Наследственный материал передаётся через соматические клетки, и потомки абсолютно тождественны родителям.
При половом размножении новый организм развивается из особой клетки (зиготы), которая образуется при слиянии половых клеток родителей. При образовании половых клеток (гамет) происходит неоднократная перекомбинация наследственного материала (мейоз) и поэтому гаметы генетически неоднородны. При оплодотворении в зиготе комбинируется наследственный материал отца и матери, поэтому потомки не абсолютно тождественны родителям, они имеют индивидуальные особенности.
Особые виды полового размножения: партеногенез и гиногенез. При партеногенезе яйцеклетки способны развиваться в целый организм без оплодотворения (насекомые, плоские черви). При гиногенезе - спермии должны присутствовать в среде для активации яйцеклеток, причём иногда это могут быть спермии другого вида (некоторые рыбы, земноводные и др.).
В развитии одного вида могут чередоваться разные способы размножения. Метагенез - чередование полового и бесполого размножения (кишечнополостные, простейшие). Гетерогенез - чередование полового размножения с оплодотворением и партеногенеза (сосальщики).
Формы полового и бесполого размножения разнообразны у одноклеточных и многоклеточных организмов.
Особый вид бесполого размножения, который встречается и у человека - полиэмбриония. Обычно из зиготы развивается один организм, но иногда, после одного или нескольких (реже) митотических делений, образовавшиеся клетки (бластомеры) могут развиваться в самостоятельные организмы. Так развиваются однояйцевые близнецы - генетически идентичные организмы.
В основе любого размножения, роста и развития организма лежат процессы репродукции клеток (пролиферация клеток). Процессы репродукции, скорость пролиферации, частота делений клеток в разных органах находятся под регулирующим контролем генетического аппарата. Если этот контроль нарушается, то начинается нерегулируемое размножение клеток - развиваются новообразования (опухоли). В течение жизни организма клетки стареют и погибают, поэтому всегда происходят процессы самообновления, образование новых клеток (физиологическая регенерация). В молодом организме репродукция клеток преобладает, за счёт чего увеличивается количество клеток, увеличиваются размеры органов и организм растёт. У взрослых - репродукция клеток обеспечивает только замещение погибших, а не увеличение их количества.
II. Клеточные популяции
Жизненный цикл клетки (ЖЦ) или клеточный цикл - период с момента возникновения клетки в результате деления материнской до ее гибели или исчезновения в процессе собственного деления.
Митотический цикл (МЦ) - период подготовки клетки к делению и само деление.
При подготовке к делению клетка проходит особые периоды интерфазы: пресинтетический (G1), синтетический (S) и постсинтетический (G2).
Таким образом, митотический цикл (МЦ) = G1 + S + G2 + митоз. Часть интерфазы, не имеющая отношения к МЦ, характеризует активное функциональное состояние (состояние покоя у камбиальных клеток) и обозначается - G0.
Жизненный цикл клеток разных популяций неодинаков (рис 13).

По продолжительности жизни и по отношению к делению различают три популяции клеток: стабильная, растущая и обновляющаяся.
Стабильная популяция. Клетки этой популяции имеют наибольшую продолжительность жизни, высокодифференцированы и не способны делиться. К такой популяции относятся нервные клетки, клетки миокарда.
Растущая популяция. Клетки этой популяции высокодиф-ференцированные, имеют большую продолжительность жизни. Они составляют основную массу клеток внутренних органов (печень, поджелудочная железа, и т. д.). Эти клетки не утратили способности к делению, они могут перестраивать свой метаболизм, снижать уровень дифференцировки и делиться.
Обновляющаяся популяция. Среди клеток этой популяции есть два типа: высокодифференцированные и недифференцированные (стволовые или камбиальные клетки).Высокодифференцировнные клетки живут недолго (часы, сутки, месяцы), неспособны к делению,
постоянно отмирают. Например, клетки поверхностного слоя эпидермиса, клетки крови, клетки слизистой оболочки кишечника.
Недифференцированные (стволовые) клетки этой популяции постоянно делятся, дифференцируются и замещают погибшие. Так, стволовые клетки эпидермиса находятся в самом нижнем (мальпигиевом) слое; стволовые клетки слизистой оболочки кишечника - в глубоких отделах кишечных крипт, стволовые клетки крови - в красном костном мозге.
Таким образом, ЖЦ клеток стабильной популяции и дифференцированных клеток обновляющейся популяции равен G0, в их жизненном цикле нет митотического цикла.
ЖЦ стволовых клеток обновляющейся популяции состоит из подготовки к делению и деления, т.е. равен МЦ (периодом G0 в данном случае можно пренебречь, т.к. клетки функционально не активны, находятся в состоянии покоя). Такой ЖЦ также имеют клетки злокачественных опухолей, т. к. они не дифференцируются в нормальные клетки, а снова и снова вступают в деление.
ЖЦ клеток растущей популяции состоит из G0+(G1+S+G2+M)
III. Регуляция временной организации клетки
В разные периоды жизненного цикла строение клетки и ее функции различны.
В интерфазе в клетке идут разнообразные метаболические процессы, происходит реализация генетической информации, клетка выполняет соответствующие функции, то есть она функционально активна (G0). При переходе клетки в МЦ перестраивается характер метаболических процессов; синтезируются новые белки (структурные и ферменты), которые будут обеспечивать нормальный процесс деления. В этот период клетка утрачивает признаки специализации (дифференцировки) и те функции, которые свойственны клеткам данного типа.
Изучены некоторые факторы и механизмы, которые изменяют временную организацию клетки и либо способствуют делению, либо тормозят деление:
1.Ядерно-плазматическое отношение, сфера влияния ядра (отношение объема ядра к объему цитоплазмы). Если это отношение уменьшается, то клетка начинает подготовку к делению.
2.Повышение концентрации в клетке предшественников ДНК (в особенности тимидина) способствует переходу клетки к МЦ. 3.При подавлении синтеза белка и РНК процессы подготовки к делению тормозятся.
4.Наличие в клетке белков-кейлонов тормозит переход клетки из одного периода в другой на любой стадии.
5. Некоторые вещества провоцируют деление клеток, которые в нормальных условиях не делятся. Например, если лейкоциты периферической крови (высокодифференцированные клетки обновляющейся популяции) поместить в среду с фитогемагглютинином (ФГА), то они начинают делиться. Это используют для изучения кариотипа.
6. Выявлен ген клеточного деления (ген CDC), который кодирует белок протеинкиназу, способствующий вступлению клетки в МЦ.
IV. Понятие о кариотипе
Генетический аппарат эукариотической клетки представлен хромосомами. Кариотип - это совокупность хромосом соматической клетки,
признак постоянный и специфичный для вида. Кариотипы разных видов различаются количеством, величиной и строением хромосом.
Количество (набор) хромосом принято обозначать как " n ". Для разных видов организмов коэффициент " n " - различный.
Каждая соматическая клетка содержит двойной (парный) набор хромосом " 2n ". Такой набор называется диплоидным. Одинаковые хромосомы называются гомологичными (парными). Половые клетки содержат одинарный набор хромосом " n ". Такой набор называется гаплоидным. У человека соматические клетки содержат 46 хромосом (23х2), из них 44 хромосомы (22х2) - аутосомы, одинаковые для мужского и женского пола и 2 хромосомы половые (гетеросомы): у женщин - ХХ, у мужчин - ХУ. По химическому строению хромосомы представляют собой дезоксирибонуклеопротеид (ДНП). В состав одной хромосомы входит одна молекула ДНК. В различные периоды жизненного цикла клетки хромосомы имеют разную морфофункциональную организацию. Хромосомы в виде плотных структур выявляются во время деления клетки, а в интерфазе они деспирализованы и вся совокупность наследственного материала носит название - хроматин. Основная биологическая роль интерфазных хромосом - передача генетической информации.
Основная биологическая роль хромосом при делении - равномерное распределение их между дочерними клетками.
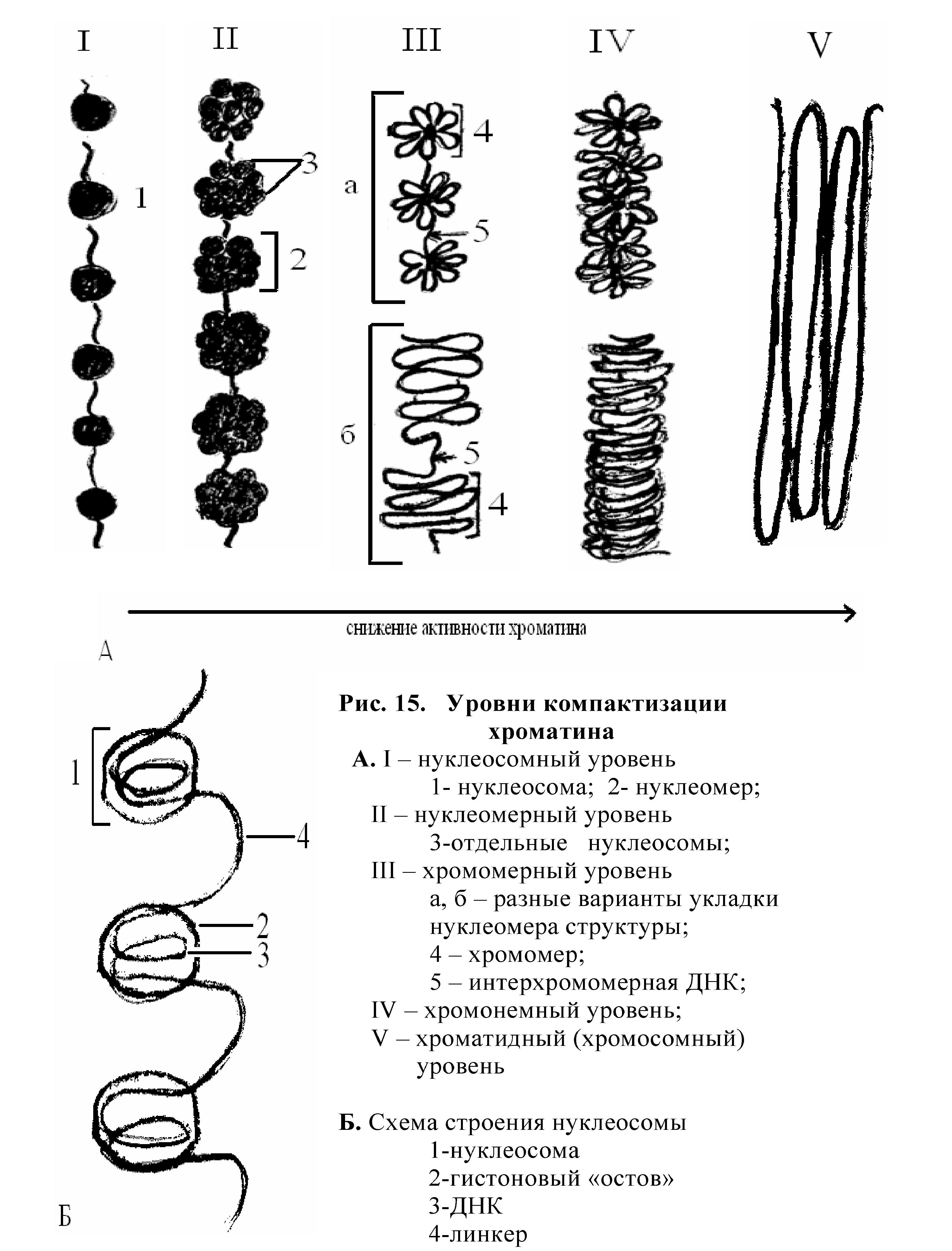
Большая часть интерфазного хроматина находится в деконденсированном состоянии, на разных его участках идут процессы транскрипции - это активный хроматин - эухроматин. Но есть участки конденсированного (плотного) хроматина - это неактивный гетерохроматин.
Различают структурный (конститутивный) гетерохроматин - участки постоянно конденсированные, неинформативные. Второй вид гетерохроматина - факультативный, это те участки, которые могут деконденсироваться и переходить в активное состояние. При подготовке к делению происходит постепенная общая конденсация хроматина и весь генетический материал представляет собой факультативный гетерохроматин; в световом микроскопе он обнаруживается в виде плотных структур - хромосом.
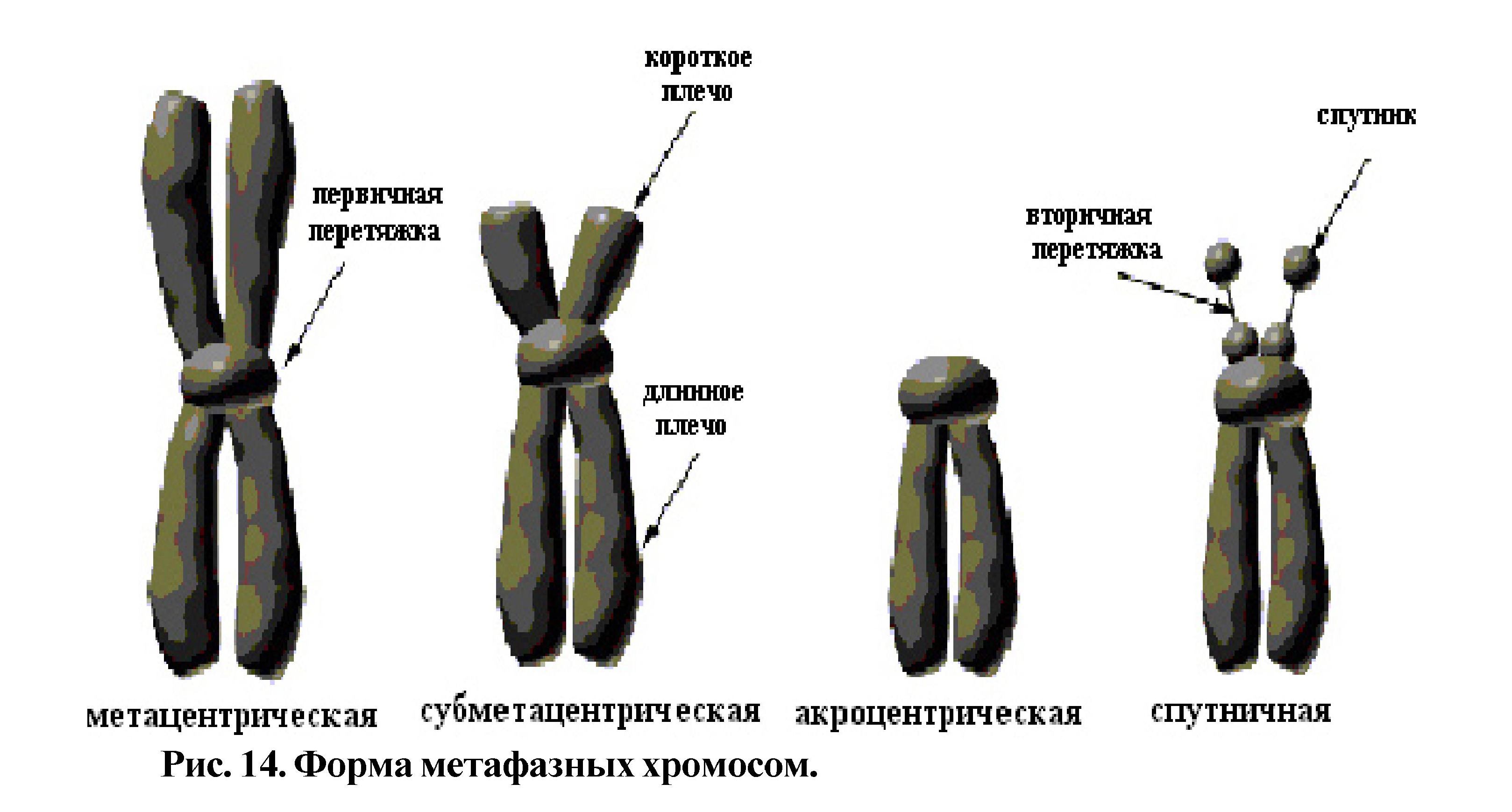
Процесс конденсации хроматина очень важен для регуляции активности генетического материала и для свободного распределения хромосом в цитоплазме клетки во время деления. По мере конденсации активность хроматина снижается. В результате усиления компактизации хроматина и процесса спирализации метафазные хромосомы уменьшаются по длине в несколько тысяч раз и свободно размещаются в цитоплазме клетки, а затем расходятся к полюсам. Форма метафазных хромосом зависит от расположения центромеры.
Различают метацентрические, субметацентрические, акроцентрические хромосомы и хромосомы с вторичной перетяжкой (спутничные) (рис. 14).
V. Способы репродукции клеток
Различают несколько способов деления клеток: митоз, амитоз, мейоз.
Митоз - универсальный способ деления клеток. Это непрямое сложное деление, характерное для соматических клеток. Биологическое значение митоза - увеличение количества генетически идентичных клеток.
Амитоз - это простое, прямое деление ядра на две или более частей. Не формируется аппарат деления, способствующий строго равномерному распределению генетического материала между дочерними ядрами. Дочерние ядра могут содержать разный объём генетического материала. Таким образом, амитоз нельзя считать полноценным делением. Деление цитоплазмы часто не происходит, и тогда образуются двуядерные (многоядерные) клетки. Такие клетки теряют способность в дальнейшем вступать в полноценное митотическое деление. Различают три вида амитоза: реактивный, дегенеративный и генеративный.
Мейоз - сложное деление, в результате которого образуются половые клетки (гаметы). Состоит из двух последовательных делений. Особенно сложным является первое деление мейоза (профаза I). При мейозе происходит перекомбинация генетического материала (кроссинговер, независимое расхождение целых хромосом в анафазе I и независимое расхождение хроматид в анафазе II). В результате мейоза образуются гаплоидные клетки ("nc") и возникает комбинативная изменчивость. Биологическое значение мейоза состоит в поддержании постоянства кариотипа и возникновении генетически неидентичных гамет, что определяет формирование организмов с индивидуальными особенностями. Мейоз происходит в процессе гаметогенеза (образование половых клеток) в половых железах (гонадах).
VI. Эндорепродукция
Эндорепродукция - явление, связанное не с увеличением количества клеток, а с увеличением (репродукцией) генетического материала в клетке. Различают два вида эндорепродукции: эндомитоз и политению.
Эндомитоз происходит при нарушении нормального хода митоза (сохранение ядерной оболочки в профазе, разрушение митотического аппарата в начале анафазы) и приводит к увеличению плоидности клетки, кратное " n ". Если в эндомитоз вступила клетка, содержавшая 2n, то образуется клетка - 4 n и т. д. Таким образом, результат эндомитоза - полиплоидия.
Политения - образование гигантских политенных (многонитчатых) хромосом. В периоде S одна репликация ДНК следует за другой десятки и сотни раз, поэтому образуются хромосомы, содержащие сотни молекул ДНК. Важным является то, что это - интерфазные хромосомы, на которых идут транскрипционные процессы (области пуффов), и это можно наблюдать в световой микроскоп. Сравнивая локализацию пуффов и синтез определённых белков, можно составить цитологические карты хромосом, то есть ориентировочно определить локализацию отдельных генов на хромосоме (рис.16). Биологическое значение политении - увеличение числа идентичных генов, и как следствие, резкая интенсификация синтеза определённых белков.
Читайте также: