
Нейронные механизмы построения движений реферат
Обновлено: 04.07.2024
Нам предстоит познакомиться с основными теор етическими сведениями об организации и контроле движений. В широком смысл е слова под нейромоторной системой понимают функционально-структурное единство различных уровней нервной системы и мускулатуры. Отношения центра и периферии (в нашем случае моторного аппарата) организованы по типу функциональных систем (П.К.Анохин, 1975). Суть этой организации заключается в том, что командный пункт - центр - имеет непрерывную информацию о выполнении задания. Это обеспечивается возвратной (обратной) афферентацией. Следует подчеркнуть важность афферентации в деятельности нейромоторной системы на всех уровнях. В деятельности центра с помощью афферентных систем строится афферентная модель будущего движения со всеми его временными и пространственными параметрами. При рассогласовании параметров идеального, афферентного образа движения и реального результата центр вносит соответствующую поправку в эфферентную систему. Следовательно, рассогласование на входе в функциональную систему является условием ее активации. Следует иметь в виду, что функциональная система не является постоянной структурой. Она может формироваться для достижения близкой цели в малые интервалы времени (часто бытовые явления), а также для решения задач долгосрочного, стратегического назначения, к примеру, творчество, образование и др. Естественно, что в этом контекст е моторика является малой частью системы. Как уже говорилось, в функциональной системе периферия и центр представляют собой единство. Сейчас нам предстоит рассмотреть основные морфо - функциональные характеристики нейромоторной системы периферической и центральной локализации.
Структурной единицей спинного мозга является сегмент, где сгруппированы нейрон ы различного назначения. Задние рога и центральная часть сегмента ориентированы на обработку сенсорных сигналов. В нейрофизиологической литературе их часто обозначают как 1-6 зоны Рекседа. Передние рога содержат тела эфферентных нейрон ов (7-9 зоны Рекседа). Отметим, что сенсорных нейрон ов в количественном отношении намного больше, чем эфферентных.
Мото нейрон ы представляют собой обособленную группу нейрон ов. Наиболее крупные из них a-мото нейрон ы с диаметром аксон а 12-22 мкм, скоростью проведения импульса 70-120 м/с, обеспечивают выполнение произвольных сокращений в поперечно-полосатой мускулатуре. Потенциал действия, возникающий в теле нейрон а в зоне аксон ного холмика проводится до конечного пункта - нервно-мышечного соединения (моторной бляшки). Здесь энерги я потенциал а действия (ПД) расходуется на высвобождение медиатора (в нервно-мышечном соединении - ацетилхолин) из пресинаптического окончания, переход его на постсинаптическую мембрану. Затем под влиянием медиатора происходит генерация нового ПД, но уже на базе мышечного субстрата. Возникающий ПД передается на соседние участки мышц (скорость проведения ПД по мышце 4-6 м/с.). Охватившая волна возбуждения вызывает пространственную перестройку сократительного субстрата мышц - образование актин-миозиновых комплексов, что сопровождается фазическим (быстрым) сокращением поперечно-полосатой мускулатуры. Кроме фазического сокращения, в мускулатуре поддерживается постоянный сократительный процесс, реализуемый особыми мышечными клетками - красными волокнами, в результате чего поддерживается постоянная готовность к быстрому сокращению. Это состояние известно как тонус мышцы. В обеспечении тонической активности мышц ведущее место принадлежит малым a-мото нейрон ам. Существует еще одна группа нейрон ов - g-эфференты, возбуждение которых не сопровождается видимым сокращением мышцы, а меняется состояние возбудимости проприо рецептор а в результате деформации участка мышцы внутри проприоцептора, т.е. интрафузального мышечного волокна.
Следует подчеркнуть, что деятельность мото нейрон ов контролируется в сложно организованной структуре двигательной системы, как в пределах сегмента, так и за счет супраспинальных влияний. Определенный Шеррингтоном "принцип общего конечного пути" заключается в конвергенции многих регулирующих влияний на мото нейрон е. Это значит, что количество афферентной информации должно испытывать значительную обработку во вставочных нейрон ах и на самом теле мото нейрон а, чтобы последний мог воспроизвести ПД необходимой интенсивности и точности.
Афферентный поток из мускулатуры организован несколькими каналами. В мышцах различают собственные проприо рецептор ы - нервно-мышечные веретена. Они соединены параллельно основной мышце. При удлинении мышцы они возбуждаются, при ее укорочении активация рецептор а снимается. Установлено, что такой вид рецепции обеспечивает центр информацией о степени растяжения мышцы и динамических нагрузках. Афференты этой группы (1а) заканчиваются моносинаптически на двигательных клетках передних рогов, практически не испытывая коррекции при прохождении через сегментарный аппарат. Это позволяет понять, что функциональная роль такого типа проприо рецептор ов заключается в осуществлении рефлекса на растяжение. Проприо рецептор ы мышц имеют разную морфологию и специализацию. Известны ядерно-сумчатые и ядерно-цепочные рецептор ы, а также так называемые вторичные окончания. Часть из них является сигнализатором статического усилия мышцы. Подчеркнем, что для нормальной функциональной активности систем построения движения важно соотношение проприоцепции динамического и статического характера.
Кроме мышечных рецептор ов важную роль в организации рефлекторной деятельности нейромоторного аппарата играют сухожильные аппараты Гольджи. Они включены последовательно (тандемно) с сухожилием, являются сигнализатором степени растягивающего усилия, развиваемой силы мышцы, дают начало афферентам 1б.
Импульсация, поступающая по афферентам Iа, вызывает в сегментах возбуждение (активацию) мото нейрон а своей мышцы и тормозит мото нейрон ы антагониста (т.е. осуществляется реципрокное торможение). Афференты группы 2, начинающиеся от вторичных окончаний веретен, путем полисинаптических влияний возбуждают мото нейрон ы сгибателей и тормозят мото нейрон ы разгибателей.
Афференты Iб вызывают торможение мото нейрон ов собственной мышцы (собственное, т.е. аутогенное торможение) и возбуждение мото нейрон ов антагонистов.
Мышечные веретена имеют и эфферентную иннервацию - к ним идут аксон ы g-мото нейрон ов, располагающихся в передних рогах сегмента. В состоянии покоя количество импульсов из веретен невелико. При растяжении мышцы частота импульсации из проприо рецептор ов повышается. У первичных окончаний частота импульсации зависит от скорости растяжения, у вторичных - от длины мышцы. Иначе, первичные окончания обеспечивают динамическую составляющую проприорецепции, вторичные - статическую.
Активация g-эфферентов приводит к повышению чувствительности нервно-мышечных веретен, т.е. рецептор ов. Установлено, что афферентация из веретен может быть осуществлена не только растяжением, но и активацией g-эфферентов. Это означает, что рефлекторная возбудимость a-мото нейрон ов зависит от состояния возбудимости g-мото нейрон а через изменение возбудимости нервно-мышечного веретена. Показано, что возбуждение a-мото нейрон ов тоже сопровождается активацией g-эфферентов опять же через изменение проприоцепции: это так называемая a-g коактивация. Состояние баланса возбудимости рефлекторного кольца обеспечивается супраспинальным контролем через кортикоспинальный и ретикулоспинальный пути. Реализуются эти влияния путем изменения возбудимости как a-, так и g- нейрон ов.
Таким образом, мышечные-веретена реагируют на два взаимодействия: периферическое и центральное. Поэтому в естественных условиях проприоцепция из мышц, сухожилий и суставов испытывает сложные взаимоотношения. Следует учесть, что проприоцепторы суставов, связок оказывают на мото нейрон ы не менее сложное влияние через афференты различного назначения и систему вставочных нейрон ов. Взаимодействие нейрон ов реализуется двумя путями - возбуждением и торможением.
Возбуждение (активация) происходит в результате деполяризации мембраны клетки, изменения ионной проницаемости с последующим формированием потенциал а действия (ПД). Торможение в своей сути тоже представляет собой активный процесс с генерацией тормозного потенциал а, отличающегося от потенциал а действия тем, что он локален, т.е. не передается по нервному волокну и имеет возможность пространственной и временной суммации. С точки зрения приложения тормозных влияний на нейрон различают пресинаптическое и постсинаптическое торможение.
Пресинаптическое торможение осуществляется на входе в нейрон ; в его основе лежит деполяризация мембраны с уменьшением величины ПД, поступающего к этому нейрон у из другого, в результате чего генерация постсинаптического ПД резко угнетается.
Постсинаптическое торможение вызывает уменьшение величины ПД нейрон а на выходе, т.е. генерация ПД постсинаптической мембраной под влиянием тормозных потенциал ов затрудняется или становится невозможной. В основе постсинаптического торможения лежат механизмы гиперполяризации мембраны синапс а.
Классическим примером пресинаптического торможения является уменьшение сенсорного потока в заднем роге спинного мозга, благодаря чему осуществляется так называемый воротный контроль. Постсинаптическое торможение, к примеру, представлено в возвратном торможении a-мото нейрон а, обеспечиваемого вставочным нейрон ом Реншоу. В свою очередь, вставочный нейрон активизируется деятельностью основного: чем интенсивнее работа a-мото нейрон а, тем более активен тормозный и тем большее тормозящее влияние испытывает мото нейрон . В сегментарном аппарате известен и другой тип торможения - двойное. В результате торможения тормозного нейрон а может быть активизирована деятельностью основного.
Таким образом, основная функциональная единица сегментарного аппарата - рефлекс. Как бы сложно не была организована рефлекторная реакция, она строго ограничена временными и пространственными параметрами, т.е. жестко детерминирована местом возбуждения, количеством активированных нейрон ных групп и способом реализации. Поскольку рефлекторные связи организованы по типу колец, включающих эфферентную иннервацию афферентных нейрон ов, этот тип организации движения называют кольцевым или коррекционным. Тем самым подчеркивается малая пластичность и функциональная жесткость описанных рефлекторных комплексов.
Супраспинальный контроль над деятельностью сегментарного аппарата осуществляется системой нисходящих и восходящих путей. Восходящая афферентация осуществляется системой проприоцептивных путей (сознательных и бессознательных), достигающих всех релейных станций обработки сенсорных сигналов. Тем самым обеспечивается разносторонняя характеристика параметров движения - сила, объем, точность, плавность, целесообразность, законченность, согласованность с вегетативными, гуморальными, поведенческими компонентами движения и др. Многоуровневая обработка сенсорного потока составляет основу акцептора действия по П.К.Анохину, параметры которого сравниваются с афферентными параметрами реального результата движения. Следует учесть, что движению дается эмоционально-личностная, т.е. сознательная оценка в пределах этой же функциональной системы.
Результат обработки сенсорного потока является базой организации нисходящего контроля из вышерасположенных структур за рефлекторной деятельностью и деятельностью афферентных систем сегментарного аппарата, обеспечивающего информацию с периферии. Объектами супраспинальных влияний являются активность мото нейрон ов, вставочных нейрон ов, нейрон ов восходящих систем и первичных афферентных волокон.
Быстропроводящие пути (кортико-, ретикуло-, вестибулоспинальные) способны возбуждать мото нейрон ы спинного мозга моносинаптически. Тем самым обеспечивается высокая эффективность и специфичность нисходящих команд. Установлено, что моносинаптические кортико-спинальные влияния наиболее активны в мото нейрон ах, иннервирующих дистальные мышцы конечностей. Ретикуло- и вестибулоспинальные пути оказывают преимущественное влияние на мото нейрон ы мышц туловища и проксимальных мышц конечностей.
Нисходящие тракты оказывают влияние и на различные типы вставочных нейрон ов спинного мозга. Наиболее важное значение, как уже говорилось, имеют связи с g- нейрон ом, результат активации которого супраспинальной командой имеет важное значение в установочной активности мускулатуры в будущем движении. Реализуется же этот тип активности изменением возбудимости проприоцептора, предварительно активированного g- нейрон ом. Иначе, в афферентной системе мышцы кодируются параметры ожидаемого движения.
Нисходящие пути могут вызвать торможение как первичных, так и вторичных афферентов восходящего направления. Это вызывает контроль интенсивности и качества сенсорного потока, доставляемого в супраспинальные структуры.
Таким образом, деятельность супрасегментарного комплекса организации движений интегрируется на многих уровнях центральной нервной системы. Основными из них являются ствол головного мозга (мозжечок, вестибулярный анализатор, ретикулярная формация, четверохолмие), подкорковые серые ядра (полосатое тело, хвостатое ядро, таламус) и сенсомоторные зоны коры больших полушарий, включающие передние и задние центральные извилины, префронтальную кору. Функциональные отношения между различными уровнями организации движения изучены Н.А.Бернштейном (1948).
Выделены уровни А, В, С, D, Е. Сущность надсегментарной организации движения заключается в формировании программ движения, алгоритм а двигательных комплексов ("мелодии движения" - по Н.А.Бернштейну). Как правило, программа движения приобретается большей частью индивидуальным опытом (игра на музыкальных инструментах, профессиональные двигательные навыки и пр.). Конечно же, в программе движения отражен и опыт видовой, т.е. генетический - содружественные движения, двигательные комплексы взаимодействия анализаторов, характерные жесты, мимика и пр.
Программа движения не предусматривает жесткую сцепленность этапов выполнения, допускается изменение параметров движения в известных пределах, т.е. пластичность организации движения. В отличие от этого, спинально-сегментарный уровень организации движения характеризуется жесткостью исполнения и текущего контроля за реализацией каждого этапа сокращения мышцы. Иначе, точность активации и торможение мото нейрон ов сегментарного аппарата точно контролируется состоянием проприоцепции в большей мере, чем супраспинальным контролем. В принципе, командные супраспинальные импульсы могут активировать большую группу мото нейрон ов или даже мото нейрон ы антагониста. Этим объясняются ошибки при заучивании новых движений, дискоординация их при многих патологических состояниях. В сегментарном же аппарате деятельность отдельного мото нейрон а может быть охарактеризована возбуждением или торможением. В наиболее общем виде следует говорить о супрасегментарном уровне организации как о качестве моторики. Сегментарный уровень обеспечивает ее количественную характеристику - силу, тонус, объем.
Постоянное взаимодействие механизмов программного, т.е. вероятностного, характера организации движения с механизмами конкретных исполнителей создает неповторимую индивидуальную моторику как в совершенном, так и искаженном виде.
Клиническая практика располагает многочисленными фактами, свидетельствующими о возможности быстрого изменения функционального состояния спинально-сегментарного аппарата, стойкости и пластичности механизмов супраспинального контроля. В качестве примера можно привести спастическую кривошею.
Известно, что спустя определенный срок от начала заболевания патологическая активность центральных механизмов гиперкинеза головы исчезает. Патологическая поза головы может быть обусловлена изменениями суставно-мышечного аппарата позвоночника, которые лечебными приемами мануальной терапии могут быть устранены достаточно быстро. Однако, у большинства больных сохраняется патологическая синкинезия - подъем плеча на стороне обращенного к нему лица. Перестройка этой сложившейся патологической программы движения - патологического двигательного стереотипа представляет собой достаточно сложную задачу.
Движения человека развиваются в онтогенезе на протяжении довольно длительного времени и проходят ряд последовательных этапов, начиная от рефлекторных движений новорожденного до координированной произвольной двигательной активности, которая может совершенствоваться, не ограничиваясь какими-либо пределами. Мастерство танцора, меткость стрелка, скорость бегуна, прыгучесть спортсмена, виртуозная игра пианиста — примеры огромных потенциальных резервов развития двигательных функций.
Содержание
1. Механизмы и закономерности их организации
2. Физиология движений: уровни организации двигательной функции.
3. Общие принципы управления движения
4. Теория построения движения Н.А. Бернштейна
5. Список литературы
Прикрепленные файлы: 1 файл
нейрофизиология.doc
1. Механизмы и закономерности их организации
2. Физиология движений: уровни организации двигательной функции.
3. Общие принципы управления движения
4. Теория построения движения Н.А. Бернштейна
5. Список литературы
Механизмы и закономерности их организации
Движения человека развиваются в онтогенезе на протяжении довольно длительного времени и проходят ряд последовательных этапов, начиная от рефлекторных движений новорожденного до координированной произвольной двигательной активности, которая может совершенствоваться, не ограничиваясь какими-либо пределами. Мастерство танцора, меткость стрелка, скорость бегуна, прыгучесть спортсмена, виртуозная игра пианиста — примеры огромных потенциальных резервов развития двигательных функций.
Основой для произвольных (сознательно регулируемых) движений служит кинестетическая афферентация и целенаправленное управление. Движение, как специальный процесс, осуществляющийся во времени, состоит из цепи закономерно сменяющихся психофизиологических событий, промежуточным и окончательным итогом которых является внешне наблюдаемое поведение человека, детерминируемое как внутренними потребностями, так и социальными факторами-условиями.
Основные разработки в области физиологии произвольных актов были проведены Бернштейном и Анохиным, проанализировавшими как многоуровневость их регуляции, так и роль в поведении афферентных механизмов.
В самом общем виде механизм движения описывался схемой и принципом рефлекторной дуги: стимул — его центральная переработка — двигательная реакция. Основным теоретическим положением, выдвинутым Бернштейном в отношении человеческого поведения, явился тезис, согласно которому сколько-нибудь сложное движение на основе указанного принципа осуществляться не может (но относительно примитивные двигательные акты типа коленного рефлекса или отдергивания руки от огня ему подчиняются). Обусловлено это тем, что сложные движения зависят не только от управляющих сигналов, но и от целого ряда дополнительных факторов, не поддающихся предварительному учету и вносящих в запланированный ход движений множество отклонений (реактивные, инерционные воздействия, внешние влияния, исходное состояние мышц). В результате окончательная цель движений может быть достигнута, только если в него будут вноситься соответствующие поправки или коррекции. Для этого ЦНС должна учитывать реальные параметры текущего движения, то есть в нее должны непрерывно поступать афферентные сигналы об актуальном положении органа, его отклонении от цели и перерабатываться в сигналы коррекции. Этот механизм регуляции выполнения сложных движений был назван принципом сенсорных коррекций. Из него логически вытекал другой принцип, сформулированный Бернштейном в 1934 г. — это принцип рефлекторного кольца — относительной замкнутости и непрерывности циркуляции сенсорной кинестетической информации, черпаемой из движения и реализующейся в двигательных актах. Обычный рефлекс в соответствии с этим принципом является лишь частным случаем движения, не нуждающегося в коррекции. Обратив внимание на качество афферентных сигналов, поступающих при движении, Бернштейн пришел к выводу, что существует несколько уровней их построения, включая различные морфофункциональные слои ЦНС — спинной и продолговатый мозг, подкорковые центры и кору.
В рамках общей системы регуляции движений традиционно выделяют три основных компонента: пирамидную, экстрапирамидную и мозжечковую системы. Это деление, основывающееся как на анатомо-физиологических, так и на клинических аргументах, в определенной степени сохраняет свое значение, и в клинике мы можем четко дифференцировать пирамидные, экстрапирамидные и мозжечковые расстройства. Однако новые экспериментальные и клинические данные показывают, что все три системы тесно взаимодействуют между собой на разных уровнях, и поражение одной из них влечет за собой вторичные изменения функционального состояния других.
По Лурия, реальным анатомическим и функциональным образованием, включенным в реализацию двигательного акта, помимо собственно моторных зон, является почти вся кора больших полушарий. Передние отделы мозга связаны с построением разворачивающихся во времени кинетических программ двигательного акта, а задние отделы — с их кинестетическим и пространственно-обусловленным обеспечением. Если же конкретизировать эфферентные механизмы произвольных движений, то традиционно к ним относят две взаимосвязанные, но относительно автономные системы — экстрапирамидную и пирамидную, корковые отделы которых составляют единую сенсомоторную зону коры. Обе системы реально представляют единый эфферентный механизм, различные уровни которого отражают этапы эволюции становления двигательных функций.
Экстрапирамидная система — обеспечивает сравнительно простые автоматизированные движения. Она управляет в основном непроизвольным компонентом движений, к которому относятся поддержание позы, регуляция физиологического тремора, физиологические синергии, общая согласованность двигательных актов, их интеграция и пластичность. (Объем произвольных движений по сравнению с тоническими составляет около 10%). Структурный состав экстрапирамидной системы среди исследователей окончательно не согласован.
Пирамидная система (кортико-спинальный путь) — начинается от крупных пирамидных клеток Беца, находящихся в основном в 5-м слое моторной коры 4-го поля передней центральной извилины. Это первичное поле, различные участки которого связаны с иннервацией соответствующих групп мышц.на растяжение. Выпадение функций пирамидного пути проявляется в невозможности произвольных движений.
Физиология движения: уровни организации двигательной функции.
Двигательная функция широко представлена в мозге человека. Движения человека очень многообразны, и многие заболевания головного мозга человека также связаны с повреждением двигательной функции. Двигательная функция в мозге человека устроена иерархично. Каждый новый морфологический этаж мозга, каждый очередной функциональный уровень приносит с собой новые полноценные движения. Наши конечности и тело – это кинематические цепи. Кинематическая цепь называется управляемой, если можно назначить желаемую для нас траекторию ее движения. Для этого необходимо связывать избыточные степени свободы. Это собственно и является основной задачей ЦНС в координации движений. Координация движений есть преодоление избыточных степеней свободы движущегося органа. Одно из решений, к которому прибегает организм, – организация синергий, т.е. содружественных движений отдельных групп мышц. Длина мышцы, необходимая для данного движения (положение мышцы), полностью задается ее жесткостью: отношением длины мышцы к растягивающей ее силе. Этот параметр в мышце регулируется специальным механизмом, который локализуется на уровне спинного мозга. Название этого механизма – рефлекс
Общие принципы управления движениями.
Управление движениями осуществляется ЦНС. Двигательная функция человека достигла наивысшего развития в связи с прямохождением и трудовой деятельностью. Поэтому в управление этой функции включены также высшие центры, включая кору больших полушарий. Это прежде всего относится к произвольным движениям – спортивным и рабочим движениям, речи и т.д.
Очень важным биомеханическим свойством опорно-двигательного аппарата организма животных и человека является наличие в скелете большого числа степеней свободы. Это происходит вследствие многозвенности скелета, а также двух- и трехосности многих суставов. С одной стороны, многозвенность скелета обеспечивает большую свободу движений, но с другой – сильно затрудняет управление такой сложной системой. В каждом конкретном случае используются только некоторые движения, но ЦНС постоянно контролирует (ограничивает) остальные, что сообщает движению устойчивость. Нарушение этой функции ЦНС приводит к характерным патологическим изменениям движений (мозжечковые нарушения и пр.).
Существуют два типа коррекции движений: при помощи обратных связей и на основе программы. Первый тип коррекций, как правило, выполняется при медленных движениях, тогда как второй при быстрых. Примерами быстрых движений могут служить печатание на машинке, кидание мяча, движения глаз в орбитах и др. В коррекции движений активно участвуют не только мышечные, суставные, сухожильные и другие рецепторы такого типа, но также зрение, слух, вестибулярный аппарат. Главным элементом нейрофизиологического механизма являются центральные генераторы движений.
Теория построения движения Н.А. Бернштейна
Основными структурными элементами системы регуляции движений являются лобная кора, базальные ганглии, мозжечок, таламус, стволовые ядра и спинной мозг. Система регуляции движении имеет иерархический многоуровневый характер, что проявляется не только в норме, но и при патологии. Выдающийся отечественный нейрофизиолог Н.А.Бернштейн (1947) разработал теорию построения движения. Согласно этой теории выделяются пять уровней построения движений:
Этот процесс создает многоуров невую структуру двигательного навыка и обозначается как автоматизация движений.
При поражении одного из уровней возникает сложное сочетание негативных (гиподинамических) симптомов, вызванных выпадением функций, и позитивных (гипердинамических) симптомов, вызванных растормаживанием активности нижележащих уровней. Характер изменения того или иного двигательного акта зависит от того, был ли пораженный уровень для него фоновым или ведущим. При вовлечении фоновых уровней смысловая структура движения сохраняется, но происходит его дезавтоматизация. При поражении ведущего для данного движения уровня страдает не только его смысловая структура, но и фоновый состав движений.
Хотя некоторые анатомо-физиологические представления Н. А. Бернштейна не соответствуют современному уровню знаний, многие положения его теории сохраняют эвристическую ценность.
Эти круги регулируют функциональную активность моторных зон коры. Многие структуры, участвующие в регуляции движений, имеют соматотопическое строение, и вертикальные связи часто объединяют группы нейронов, контролирующие движения в одной и той же части тела. Кроме того, вертикальные кольцевые связи объединяют и те группы нейронов, которые, располагаясь на различных уровнях нервной системы, специализируются на регуляции одних и тех же параметров.
Программирование движений. Каждому целенаправленному движению предшествует формирование программы, которая позволяет прогнозировать изменения внешней среды и придать будущему движению адаптивный характер. Результат сличения двигательной программы с информацией о движении, передающейся по системе обратной связи, является основным фактором перестройки программы. Последнее зависит от мотивированности движения, его временных параметров, сложности и автоматизированности.
Мотивации определяют общую стратегию движения. Каждый конкретный двигательный акт нередко представляет собой шаг к удовлетворению той или иной потребности. Биологические мотивации приводят к запуску либо жестких, в значительной степени генетически обусловленных моторных программ, либо формируют новые сложные программы. Однако мотивация определяет не только цель движения и его программу, она же обуславливает зависимость движения от внешних стимулов. В качестве обратной связи здесь выступает удовлетворение потребности.
Двигательная команда определяет, как будет осуществляться запрограммированное движение, т.е. каково распределение во времени тех эфферентных залпов, направляемых к мотонейронам спинного мозга, которые вызовут активацию различных мышечных групп. В отличие от программ команды движения должны точно соответствовать функциональному состоянию самого скелетно-двигательного аппарата как непосредственного исполнителя этих команд. Непосредственное управление движением обуславливается активностью моторной зоны коры, полосатого тела и мозжечка. Полосатое тело участвует в преобразовании "намерения действовать" в соответствующие "командные сигналы" для инициации и контроля движений.
Особую роль в программировании движения играют ассоциативные системы мозга, и в первую очередь таламопариетальная ассоциативная система. Во-первых, именно она участвует в формировании интегральной схемы тела. При этом все части тела соотносятся не только друг с другом, но и с вестибулярными и зрительными сигналами. Во-вторых, она регулирует направление внимания к стимулам, поступающим из окружающей среды так, чтобы учитывалась ориентация всего тела относительно этих стимулов. Эта система "привязана" к настоящему моменту времени и к анализу пространственных взаимоотношений разномодальных признаков.
Таламофронтальная ассоциативная система отвечает за переработку информации о мотивационом состоянии и происходящих в организме вегетативных изменениях. Фронтальная ассоциативная область коры опосредует мотивационные влияния на организацию поведения в целом благодаря связям с другими ассоциативными областями и подкорковыми структурами. Таким образом, фронтальные отделы коры больших полушарий, контролируя состояние внутренней среды организма, сенсорные и моторные механизмы мозга, обеспечивают гибкую адаптацию организма к меняющимся условиям среды.
Функциональная структура произвольного движения. Из вышеизложенного следует, что в обеспечении любого движения принимают участие разные компоненты, поэтому один из главных вопросов состоит в том, каким образом обеспечивается единовременность команды, поступающей к исполнительным аппаратам. Независимо от стратегии и тактики конкретного движения, основная задача системы, обеспечивающей программу, заключается в координации всех компонетов команды.
ЦНС располагает некоторым числом генетически закрепленных программ (например, локомоторная программа шагания, базирующаяся на активности спинального генератора). Такие простые программы объединяются в более сложные системы типа поддержания вертикальной позы. Подобное объединение происходит в результате обучения, которое обеспечивается благодаря участию передних отделов коры больших полушарий.
Самой сложной и филогенетически самой молодой является способность формировать последовательность движений и предвидеть ее реализацию. Решение этой задачи связано с фронтальной ассоциативной системой, которая запоминает и хранит в памяти такие последовательности движений. Высшим отражением этого кодирования у человека является вербализация, или словесное сопровождение, основных понятий движения.
Всеобщей закономерностью работы системы управления движениями является использование обратной связи. Сюда входит не только проприоцептивная обратная связь от начавшегося движения, но и активация систем поощрения или наказания. Кроме того, включается и внутренняя обратная связь, т.е. информация об активности нижележащих уровней двигательной системы, или эфферентная копия самой двигательной команды. Этот вид обратной связи необходим для выработки новых двигательных координаций. Для движений различной сложности и скорости обратная связь может замыкаться на разных уровнях. Поэтому оба типа управления — программирование и слежение — могут сосуществовать в системе управления одним и тем же движением.
В заключение целесообразно привести высказывание выдающегося физиолога Н.А. Бернштейна о том, что движения ". ведет не пространственный, а смысловой образ и двигательные компоненты цепей уровня действий диктуются и подбираются по смысловой сущности предмета и того, что должно быть проделано с ним".

Координационные способности – это совокупность психологических, морфологических, физиологических компонентов организма человека, единство которых в границах функциональной системы обеспечивает продуктивную двигательную деятельность, то есть умение целесообразно строить движение, управлять им и в случае необходимости быстро перестраивать его.
Они зависят от уровня двигательной подготовки и врожденных качеств спортсмена. Н.А. Бернштейн (1966) указывает, что координация – это преодоление избыточных степеней свободы органов движения, превращение их в управляемые системы. На основании информации от органов чувств, интегрирующейся в ЦНС, в движения вносятся непрерывные поправки, то есть происходит их сенсорная коррекция. Таким образом, автор представляет ее как кольцо афферентно-эфферентных импульсов, которые приводят к формированию определенных двигательных актов, исходя из изменяющихся условий. При этом внешняя структура движений не меняется, благодаря использованию различных мышечных волокон, обеспечивающих поддержание позы, равновесия и т.д.
Координационная сложность выполняемых двигательных актов зависит от способности нервной системы и ее высших отделов к согласованной и оперативной переработке информации.
Структуры, обеспечивающие двигательную активность человека, расположены в различных отделах центральной нервной системы – от спинного мозга до коры больших полушарий. Повышение спортивной результативности во многом обусловлено совершенствованием координационной деятельности между нервными центрами.
Координированность работы нервных центров, регулирующих определенные двигательные акты, обусловлена постоянным импульсным воздействием на них проприоцептивных анализаторов. При выработке устойчивых навыков движения формируется функциональная система управления движением и взаимосвязанные центры коры больших полушарий образуют единую структуру. С ростом квалификации спортсмена отдельные функциональные зоны объединяются в плеяду, которые, как правило, локализуются в левом полушарии и функционально изолируются от других корковых зон. На электроэнцефалограмме это выглядит как классический механизм формирования доминантных очагов. Такая синхронизация структур коры по электроактивности происходит по частоте, которая соответствует темпу выполняемого (или воображаемого) движения.
Таким образом, систематические занятия приводят к специфическим приспособительным реакциям на уровне центральной нервной системы, способствующим проявлению высокой координированности движений и совершенствованию функций нервно-мышечного аппарата.
Необходимый для адекватного ответа центральной нервной системы поток информации о длине мышц, степени их растяжения, расположения в пространстве звеньев тела и углах в суставах поступает от рецепторов сухожилий мышц, связок (проприорецепторов); об особенностях взаимодействия организма с внешней средой – от рецепторов кожи, вестибулярного аппарата и сенсорных систем.
Достижение высоких результатов в спорте возможно лишь при условии развития способностей точно оценивать и регулировать динамические, пространственные и временные параметры движения. Поэтому степень развития координационных способностей спортсмена зависит от его способности к переработке информации от различных сенсорных систем и обеспечения адекватного двигательного действия, то есть определяется уровнем его сенсомоторной координации.
Сенсомоторная координация (от лат. sensus – чувство, ощущение, motor – двигатель, со – вместе, ordinatio – расположение в порядке) – согласование во времени и пространстве двигательных действий. Еще И.П. Павлов, исследуя физиологические механизмы произвольных движений, обратил внимание на ассоциирование нейронов двигательного анализатора с многочисленными нейронами коры больших полушарий, при передаче импульсов от внешних и внутренних рецепторов.
Американские ученые также полагают, что сенсомоторная координация – это развитие, управление и коррекция движений с помощью сенсорных систем (органов чувств): зрительной, двигательной, вестибулярной, слуховой и др. При этом коррекция осуществляется корковыми структурами на двигательные единицы (Дж.Х. Уилмор, Д.Л. Костилл, 2001).
Физиологическим механизмом сенсомоторной координации является система обратной афферентации и она представляет собой сложный навык. На начальных этапах обучения выполняемые действия в составе сенсомоторной координации, могут состоять из отдельных реакций, затем они объединяются в пластичную систему. Уровень сложности проявлений сенсомоторной координации является отличительным признаком новичка от мастера.
С точки зрения физиологии, Дж.Х. Уилмор, Д.Л. Костилл (2001) описывают явление сенсомоторной координации следующей последовательностью событий:
– чувствительные рецепторы принимают сенсорные стимулы;
– импульс передается в центральную нервную систему (ЦНС). Участок, где заканчивается передача импульсов называется интеграционный центр;
– интеграционный центр ЦНС перерабатывает поступившую информацию и определяет наиболее подходящую ответную реакцию;
– принятое решение передается двигательным нейронам;
– импульс от двигательных нейронов пересылается мышцам и осуществляется ответная реакция.
Система управления движениями достаточно сложная, она включает в себя структуры спинного мозга (интеграционный центр простого двигательного рефлекса); продолговатого мозга (вызывают подсознательные двигательные реакции, например, постуральный контроль); среднего мозга (проводящие пути); промежуточного мозга (формирование новых навыков), интеграционные центры таламуса начинают различать ощущения и двигательные действия достигают сознания; лимбической системы (обеспечивает эмоциональность двигательных действий), мозжечка (сенсомоторная координация позы и двигательного акта). Основная же координация деятельности различных звеньев тела и мышечных групп в пространстве и времени осуществляется корой больших полушарий (теменная доля, зона постцентральной извилины). Таким образом, координация двигательной деятельности осуществляется всеми структурами центральной нервной системы.
С позиции психофизиологии, деятельность в спорте, заключается, прежде всего, в пространственно-временной организации психомоторных реакций двигательных актов, которые являются специфичными для каждого вида спортивной деятельности. Важнейшими элементами такой организации являются:
– сенсорный процесс обнаружения и восприятия внешнего стимула, целью действий на который является двигательная реакция;
– центральные процессы переработки воспринятого стимула с его различением и оценкой;
– моторный процесс, определяющий начало движения.
По сложности протекания центрального процесса различают реакции простые и сложные. Простая сенсомоторная реакция – это ответ на внезапно появляющийся, но при этом заранее известный сигнал простым одиночным движением, также заранее известным. Скорость реакции оценивается либо по времени реакции с момента подачи сигнала до ответного действия, либо по общему времени реагирования. Продолжительность латентного периода простой реакции зависит, в основном, от скорости передачи нервных импульсов по всей рефлекторной дуге от рецепторов до двигательных единиц. Скорость двигательной реакции определяется интенсивностью предъявляемого стимула, интервала времени между стимулами, способностью к предвидению (антиципацией), видом сигнала (звуковой зрительный, и т.д.), текущим функциональным состоянием спортсмена.
Сложная сенсомоторная реакция зависит от центральных процессов переработки воспринятого стимула и может быть:
– реакцией выбора, если необходимо выбрать двигательный ответ из ряда возможных;
– реакцией различения, если вид выполняемого движения зависит от типа сигнала, при этом на другие сигналы не надо делать никакого движения;
– реакцией переключения, если при изменении смысловой связи подаваемых стимулов необходимо выбирать возможные двигательные акты;
– реакция задержки, которая заключается в том, что двигательное действие должно быть не столько быстрым, сколько своевременным (когда ответная реакция должна быть отделена от стимула определенным интервалом времени.
Сенсомоторная координация является проявлением индивидуальных свойств личности и субъективными условиями успешного осуществления двигательной активности, обнаруживается в быстроте и прочности овладения приемами двигательных действий. То есть, является основным условием проявления координационных способностей.
В ходе анализа научно-исследовательской литературы не было выявлено единого мнения относительно сенситивных периодов развития координационных способностей. Это можно связать с многообразием проявления координационных способностей, каждые из которых имеют свои физиологические механизмы, а так же тем, что в онтогенезе можно выявить несколько сенситивных периодов развития каждого из видов координационных способностей. Так В.К. Бальсевич (1985) и Г.В. Доля (1973) утверждают, что координационные способности интенсивнее всего развиваются в дошкольном возрасте параллельно с освоением основных локомоций. Н.А. Фомин, Ю.А. Вавилов (1991) выделяют возраст от 8 до 12 лет, поскольку для этого возрастного периода характерна
пластичность ЦНС, интенсивное совершенствование проприоцептивного анализатора, выражающееся в частности, в улучшении пространственно-временных характеристиках движения. Л.В. Волков (1981) и В.А. Ярмалюк (1989) высказывают мнение, что наиболее интенсивное развитие координационных способностей наблюдается в 11–12 лет и последующие годы идет уже их стабилизация. D.-D. Blume (1978) в своих работах указывает, что сенситивными периодами в дифференциации мышечных усилий – 6–10 лет, ритмичность движений 9–11 лет, ориентировка в пространстве 11–15 лет, равновесие 10–12 лет.
Управление двигательными актами строится на двух основных принципах — принципе сенсорных коррекций текущего движения по цепи обратной связи и принципе прямого программного управления. Последний особенно важен для тех случаев, когда имеются быстрые изменения в системе, ограничивающие возможность сенсорных коррекций.
Накоплено множество экспериментальных фактов, подтверждающих реальное существование двух механизмов управления движением; посредством центральных моторных программ и с помощью обратной афферентации, которая используется для непрерывного контроля и коррекции выполняемого движения. Для многих
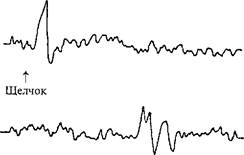
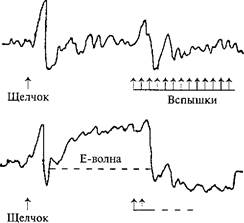
Рис. 52.Волна ожидания при отведенииЭЭГ от вертекса.
1 и 2 — реакции на изолированное действие щелчка и вспышек света; 3 —накомбинированное действие обоихстимулов, следующих один задругим; 4 — Е-волна (ожидания), когда испытуемыйдолжен был нажиматьна кнопку припоявлении вспышек света и тем самымих прерывать.Интервал междущелчком (предупреждающим стимулом) и вспышками (императивным стимулом)фиксирован. Видна медленная негативность (отклонение вверх), предшествующаяпоявлению императивного стимула (по Г. Уолтеру, 1966).
видов движения управление может осуществляться одновременно двумя механизмами при разном их соотношении для движений, различающихся сложностью и уровнем организации
Существуют два типа командных нейронов.Одни из них лишь запускают ту или другую двигательную программу, ноне участвуют в ее дальнейшем осуществлении.Это нейроны-тригеры. Приме-
ром такого нейрона является маутнеровская клетка рыбы. Командные нейроны другого типа получили название воротных нейронов. Они поддерживают или видоизменяют двигательные программы, лишь будучи постоянно возбужденными. Такие нейроны обычно управляют позными или ритмическими движениями. В качестве примера можно привести нейрон генератора локомоторного ритма у таракана.
На принципиальную роль афферентации в регуляции движения и поведения в целом указывали НА. Бернштейн (1966) и П.К. Анохин (1968). Сенсорные раздражения не только могут запускать движение, но и выполнять корректирующую функцию. Обратная аффе-рентация, сигнализирующая о результатах действия, сопоставляется с программой движений и служит уточнению координат цели и траектории движения.
До недавнего времени мало что было известно о процессах формирования и построения новых моторных программ. Однако благодаря изучению нейронной активности корыбольших полушарий и других структур мозга у высших животных, а также клиническим данным, полученным при наблюдении за человеком, сложилось мнение, что ведущая роль в построении новых моторных программ принадлежит лобным отделам коры больших полушарий (префронтальной коре). Поражение в префронтальной коре может привести вообще к распаду произвольной регуляции движений. Этот дефект особенно выражен, когда требуется менять программы движений. Передние отделы коры для построения новых моторных программ используют весь видовой и накопленный в течение жизни индивидуальный опыт. При этом, по-видимому, происходит выбор отдельных фрагментов из хранящихся в памяти программ (как для врожденных форм поведения, так и приобретенных двигательных автоматизмов) дляих последующей интеграции в новую моторную программу.
Первая группа клеток реагировала только на условный сигнал и при этом по-разному на зажигание лампы слева и справа, т.е. эти нейроны реагировали на пространственные свойства условного раздражителя. Они были отнесены автором к сенсорным нейронам с пространственной селективностью. Вторая группа нейронов активировалась только в период отсрочки, обнаруживая селективность к условному сигналу слева и справа. Третья группа нейронов, обладавших также свойством пространственной селективности, отвечала лишь после открытия экрана, т.е. в пусковой период программы.
Исследователи предполагают, что сенсорные нейроны (первая группа) передают информацию второй группе нейронов, принадлежащей кратковременной памяти, которые возбуждаются в течение всего периода отсрочки, сохраняя тем самым полученную информацию. Третья группа нейронов — нейроны моторных программ, вероятно, получают информацию от нейронов памяти и запускают хорошо отработанную двигательную реакцию нажатия на правый или левый рычаг. Таким образом, нейроны передних отделов коры мозга принимают непосредственное участие в построении сложного двигательного акта, отвечающего требованиям пространственно-временных характеристик среды.А.С. Батуев говорит о последовательном эстафетном вовлечении различных популяций клеток в процесс программирования нового двигательного акта (Батуев А.С., 1991).
Программирующая функция префронтальной коры исследовалась также П. Гольдман-Ракич и ее коллегами на модели выработки отставленного инструментального глазодвигательного рефлекса у обезьяны. В этих опытах, так же как и в опытах С.А. Батуева, процедура отставленного ответа позволяла оценивать способность животного реагировать на те или иные ситуации, опираясь на представления о внешней среде, сохраняющиеся в рабочей памяти.
П. Гольдман-Ракич и ее коллеги рассматривают префронталь-ную кору как посредника между памятью и действием. В разных ее участках в режиме рабочей памяти кодируется информация о местоположении объектов, их признаках (цвете, размере, форме), а у людей — семантические и математические знания. Исследователи полагают, что префронтальная кора извлекает информацию из
Существует и другое объяснение взаимосвязи обеих структур. Гиппокамп, так же как и префронтальная кора, причастен, к извлечению информации из долговременной памяти, которая ему необходима для сопоставления с текущими стимулами, чтобы вынести вердикт о новизне или его тождестве с прогнозом.
Сейчас уже многое известно о функциях моторной коры. Ее рассматривают как центральную структуру, управляющую самыми тонкими и точными произвольными движениями, посылающую свои сигналы к мотонейронам спинного мозга (так называемая пирамидная система). Именно в моторной коре строится конечный и конкретный вариант моторного управления движением. Моторная кора использует оба принципа управления; контроль через петли обратной сенсорной связи и через механизм программирования. Это достигается тем, что к ней сходятся сигналы от мышечной активности, от сенсомоторной, зрительной и других отделов коры, которые и используются для моторного контроля и коррекции движения.
Моторная кора включает первичную и дополнительную моторные области (ДМО), характеризующиеся соматотопической организацией с элементами множественного представительства периферии в этих отделах моторной коры- Первичная моторная кора расположена вдоль центральной борозды преимущественно в пре центральной извилине. В 5-м слое первичной моторной коры находятся гигантские клетки Беца, аксоны которых входят в состав пирамидного тракта, образующего эфферентные пути от моторной коры к мотонейронам спинного мозга. ДМО расположена в глубине межполушарной щели, примыкая к первичной моторной коре.
Клетки первичной моторной коры образуют колонки, которые возбуждают и тормозят группу функционально близких мотонейронов. Двигательная колонка представляет собой объединение нейронов, регулирующих работу нескольких мышц, действующих на сустав. При раздражении через микроэлектрод различных колонок возникают разнообразные движения в определенном суставе. Таким образом, в разных колонках представлены не отдельные мышцы, а разные движения. Это объясняет результаты А. Георго-пулоса (Сеог§ори1о5 А.) из университета Дж. Гопкинса, который нашел в моторной коре (в прецентральной извилине) обезьяны нейроны, кодирующие движения руки.Он вычислял векторы дви-
жений на основе суммирования частоты импульсации многих нейронов, возбуждающихся перед движением. По его данным, вектор движения определяется результатом активности не более чем 100 нейронов. Он формируется за несколько миллисекунд до сокращения соответствующих мышц, приводящих руку в движение.
Премоторная кора (дорзальная и вентральная области) содержит представительство каждой ноги и руки, образуя прямые связи с мотонейронами спинного мозга. Дорзальная Премоторная кора, кроме того, имеет раздельное представительство дистальной и про-ксимальной части руки. Разряды нейронов дорзолатеральной пре-фронтальной и дорзальной премоторной коры тесно коррелируют со зрительно управляемыми движениями, характеризуясь селективностью в отношении направления движений конечностей.
Возникает вопрос: что именно отражает разряд нейронов в премоторной и префронтальной коре? Чтобы ответить на него, Г. Пеллегрино и С. Вайс (РеНееппоО., \У15е 5.) провели две серии опытов. В одной серии обезьяна двигала рычаг в направлении к цели, в другой — появление стимула в разных точках пространства служило сигналом к движению всегда в одном направлении. Стимул мог появляться в одной из 8 позиций при строгой фиксации взора на центр экрана. Оказалось, что премоторные нейроны разряжались избирательно по отношению к целевому движению, а не к самому движениюкак таковому. В отличие отних нейроны префронтальной коры отвечали избирательно на цель независимо от типа движения.
Важную функцию в управлении движением выполняет мозжечок.Он обеспечивает сохранение равновесия, поддержание позы, регуляцию и перераспределение мышечного тонуса, тонкую координацию движений. Нейроны моторной коры находятся под контролирующим влиянием мозжечка. В опытах с регистрацией нейронной активности у обезьян показано, что при выполнении ими заученного движения активность нейронов зубчатого ядра мозжечка на 10 мс опережает изменение активности нейрона в моторной коре, которое предшествует появлению мышечного движения. Влияние сигналов, поступающих из мозжечка, на активность нейронов моторной коры у обезьян также установлено в опытах с временным понижением температуры мозжечка. Во время его охлаждения импульсация нейронов моторной коры и соответствующее выученное движение запаздывали или вообще были невозможны.
Многие авторы отождествляют мозжечок с мощным процессором, в котором перерабатывается огромная информация. Полагают, что он обеспечивает временную избирательную настройку при выполнении любого двигательного акта, точное выполнение дви-
жения во времени. Поражение мозжечка ведет кдисметрии — плохому выполнению точных движений.
Кроме сигналов от мозжечка, в моторную кору поступают сигналы от базальных ганглиев — структуры, которая ответственна за хранение как двигательных программ врожденного поведения (пищевого, питьевого и др.), так и приобретенных навыков. Показано, что клетки базальных ганглиев, так же как и мозжечка, разряжаются задолго до движений, совершаемых животными в ответ на сигнал. По данным Н.Ф. Суворова (1980), разряды нейронов хвостатого ядра кошки на 50—150 мс опережают ЭМГ-ком-понент хватательного движения лапы, направленного на пищу, т.е. возникающего как натуральный пищевой рефлекс. В хвостатом ядре были найдены также нейроны, у которых возбуждение опережало условную двигательную пищевую и оборонительную реакции. Эти реакции у нейронов появлялись с выработкой условного рефлекса и исчезали с его угасанием. Нейроны с реакциями опережающими двигательные оборонительные и пищевые условные рефлексы, локализованы в различных частях хвостатого ядра.
Сходный механизм действует и в системе черная субстанция — ХЯ. Черная субстанция относительно ХЯ выполняет функцию, аналогичную ВТО для прилегающего ядра. Однако стимулами, запускающими эту цепь реакций, являются сигналы не из лимби-
Рис. 53. Схема связей хвостатого ядра и прилегающего ядра — двух ключевых структур базальных ганглиев с корой и со структурами, распространяющими ДА-ергические влияния. Ассоциативная кора больших полушарий илимбическая система инициируют программы моторных актов через базальные ганглии. ВТО — вентрально-тегментальная область.
ческой системы, а из ассоциативной коры, и прежде всего из пре-фронтальной, которая имеет прямую проекцию на ХЯ. Через бледный шар ХЯ и прилегающее ядро запускают исполнение поведенческого акта, управляя моторными системами нижнего ствола мозга. Параллельно сигналы через таламус посылаются в моторную кору, которая включает контролирующий механизм, влияющий на выполнение действия.
Таким образом, моторная кора управляет движением, используя информацию, поступающую как по сенсорным путям от других отделов коры, так и от генерируемых в ЦНС моторных программ, которые актуализируются в базальных ганглиях и мозжечке и доходят до моторной коры через таламус и префронтальную кору.
Согласно гипотезе, высказанной П. Робертсом, актуализация моторных программ происходит вследствие активации командных нейронов. Сами командные нейроны могут контролироваться и затормаживаться сверху. Снятие торможения с командных нейронов повышает их возбудимость и тем самым высвобождает
По-видимому, существует общий биохимический механизм актуализации моторных программ в результате роста активности ДА- и НА-ергических систем в мозге. Эти системы обладают свойством тормозить тормозные интернейроны и тем самым растормаживать нейронные сети.
Имеются данные о наличии в стриатуме ГАМК-ергических тормозных и нтер нейронов, которые тонически тормозят запрограммированные нейронные цепи, управляющие позой и движениями, а сами тормозятся ДА-ергическими нейронами из черной субстанции. Показано, что НА-волокна проецируются в мозжечок, их активация также ведет к торможению тормозных интернейронов.
Сейчас многие исследователи разделяют точку зрения, согласно которой ДА- и НА-ергические пути участвуют в подавлении активности тормозных интернейронов в структурах, вносящих вклад в инициацию и двигательный контроль поведения- Таким образом, в базальных ганглиях, мозжечке уже заложен механизм, который может актуализировать хранящиеся в них двигательные программы. Однако для приведения в действие всего механизма, необходимо, чтобы в эти структуры поступил сигнал, который послужил бы начальным толчком процесса.
Читайте также: