
Характеристика биосинтеза днк реферат
Обновлено: 05.07.2024
Репликация- самовоспроизведение молекулы ДНК с целью передачи генетической информации. В репликации ДНК участвует сложный репликационный комплекс, который включает в себя:
- нуклеотиды в виде трифосфатов (АТФ, ГТФ, ТТФ, ЦТФ). Они являются одновременно и структурным материалом и источником энергии;
- ДНК-матрица – программа, по которой будет синтезироваться ДНК;
- праймер (затравка) - короткая молекула РНК, к которой присоединяется первый нуклеотид ДНК;
- ферменты:
- ДНК-полимеразы соединяют нуклеотиды в полинуклеотидную цепь,
- ДНК-рестриктазы разрывают полинуклеатидную цепь ДНК.
- ДНК-лигазы соединяют фрагменты ДНК,
- хеликазы раскручивают молекулу ДНК,
- топоизомеразы осуществляют кратковременные разрывы ДНК в процессе синтеза и контролируют конформацию вновь синтезированной ДНК,
- праймазы – осуществляют синтез затравки;
- белковые факторы;
- ионы металлов.
Репликация ДНК происходит в S-фазу клеточного цикла. Важную роль в чередовании циклов выполняют белки - циклины.
Сущность репликации сводится к соединению нуклеотидов в комплементарную антипараллельную дочернюю цепь ДНК. В репликации выделяют 3 стадии: инициация (начало синтеза), элонгация (удлинение цепи), терминация (окончание синтеза).
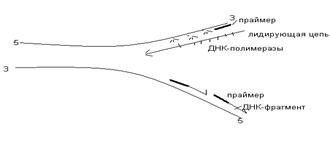
Матрицей для синтеза ДНК являются обе полинуклеотидные цепи материнской ДНК. Репликация начинается в нескольких участках материнской ДНК (точки репликации, точки Ориджины). В них происходит частичный разрыв водородных связей с формированием репликационных утолщений (узелков). В последующем, при участии ферментов ДНК-рестриктаз происходит разрезание молекулы ДНК. Под действием ферментов хеликаз возникает расплетание (раскручивание) двух цепей, и при участии топоизомераз формируется репликационная вилка, в которой одна цепь имеет направление 5→3, а вторая 3→5. Затем на каждой из ПНЦ синтезируются дочерние нити ДНК по принципу комплементарности. Поскольку ДНК- полимераза замыкает связь только в направлении 5’→3’, то на одной цепи ДНК происходит синтез непрерывной (лидирующей) нити ДНК в направлении 5→3. При синтезе лидирующей цепи праймазы синтезируют праймер, затем ДНК- полимераза присоединяет к праймеру первый ДНК-овый нуклеотид и по принципу комплементарности происходит удлинение молекулы ДНК. На второй материнской нити ДНК синхронно синтезируется отстающая нить ДНК, которая синтезируется в виде небольших фрагментов в направлении 5→3. В последующем эти фрагменты (фрагменты Оказаки) соединяются между собой ДНК-лигазами. Праймеры при этом расщепляются.
Таким образом, биосинтез ДНК проходит по полуконсервативному типу, при котором в новой ДНК одна цепь материнская, а другая - дочерняя.
Репарация ДНК происходит при появлении в её молекуле повреждений, искажений. При репарации вначале распознаётся место повреждения, затем ферменты рестриктазы вырезают дефектный участок, ДНК-полимеразы по принципу комплементарности синтезируют отсутствующий участок, а ДНК-лигазы прикрепляют его к сохранившимся участкам неповреждённой ДНК.
Биосинтез РНК (транскрипция)
Транскрипция – синтез молекулы РНК по матрице ДНК. Биологическая роль: перенос генетической информации с ДНК на РНК. Для транскрипции необходимы:
- матрица (программа) – кодирующая нить ДНК,
- субстраты – АТФ, ГТФ, УТФ, ЦТФ,
- ферменты – РНК-полимеразы,
- белковые факторы,
- ионы магния, марганца.
Выделяют 3 стадии транскрипции: инициация, элонгация, терминация
На молекуле ДНК имеется особый участок промотор, с которым связывается РНК-полимераза. Промотор иногда называют ТАТА участком (преобладает тимин, аденин, между которыми 2 Н-связи). Рядом с промотором расположены сигнальные участки, определяющие скорость транскрипции. Далее в молекуле ДНК располагаются кодирующие (экзоны) и некодирующие (интроны) участки гена. Участок (сайт) терминации определяет окончание синтеза РНК.
Инициация заключается во взаимодействии иницирующих белков с промотором и расхождении нитей ДНК, их раскручивании и формировании транскрипционной вилки. РНК-полимераза связывается с промоторным участком и по принципу комплементарности соединяет нуклеотиды цепи РНК в направлении от 5 конца к 3. РНК-полимераза - это олигомерный фермент, состоящий из нескольких субъединиц, не требующий затравки. При достижении РНК – полимеразой участка терминации происходит его связывание с белками терминации, что сопровождается отсоединением РНК-полимеразы от ДНК, диссоциацией её и окончанием транскрипции.
Процессинг РНК
Синтезированная РНК переписывает с кодирующей нити ДНК, как кодирующие участки, так и некодирующие участки гена и является про-РНК (незрелой РНК). Про-РНК в последующем подвергается созреванию (процессингу). Существует несколько механизмов процессинга:
- сплайсинг – вырезание копий интронов и соединение копий экзонов;
- присоединение к про-РНК добавочных нуклеотидов;
- модификация азотистых оснований в составе про-РНК
Особенности процессинга для рРНК, тРНК, иРНК.
Процессинг иРНК заключается в добавлении КЭП - участка и полиаденилового хвоста в сочетании со сплайсингом.
Процессинтг тРНК происходит путём метилирования азотистых оснований и добавления акцепторного участка ЦЦА в сочетании со сплайсингом.
Процессинтг рРНК заключается в вырезании из большого предшественника фрагментов всех видов РНК: 18S; 5S; 5,8S; 28S;.
Возможен альтернативный сплайсинг, который состоит в том, что для разных белков интроны могут играть роль экзонов.
Возможен и безматричный синтез РНК; он происходит из нуклеозиддифосфатов при участии фермента полинуклеотидфосфорилазы. В этом варианте синтезируются стандартные, небольшие молекулы РНК, необходимые для синтеза стандартных белков.
Таким образом, передача генетической информации происходит в следующем направлении: ДНК→ РНК→ белок. Однако, в некоторых фагах, эмбриональных тканях возможен синтез ДНК по матрице РНК (РНК→ДНК). Этот вариант синтеза катализирует фермент РНК-зависимая ДНК-полимераза (обратная транскриптаза, ревертаза). В вирусах возможен также вариант синтеза РНК→РНК при участии РНК-репликазы.

Прежде чем изложить современные представления о механизме биосинтеза ДНК, следует представить сведения о синтезе этого соединения в бесклеточной системе, которыми располагает биохимия. Известно, что для любого синтеза полимерной органической молекулы, осуществляемого in vitro или in vivo, требуется энергия. Источником энергии в реакциях полимеризации мононуклеотидов является энергия, освобождаемая всеми четырьмя типами дезоксирибонуклеозидтрифосфатов, участвующих в синтезе ДНК. Образующийся пирофосфат под действием пирофосфатазы также расщепляется на две молекулы ортофосфата, давая дополнительную энергию для биосинтеза ДНК.
Помимо энергии, биогенез ДНК требует наличия специфических ферментов, катализирующих отдельные этапы синтеза, и множества белковых факторов, абсолютно необходимых для регулирования процесса репликации и проявления каталитической активности ферментов.
Ферментные системы синтеза ДНК у про- и эукариот до конца не выяснены. По имеющимся данным, в репликации ДНК, включающей узнавание точки начала процесса, расплетение родительских цепей ДНК в репликационной вилке, инициацию биосинтеза дочерних цепей и дальнейшую их элонгацию и, наконец, окончание (терминация) процесса, участвует более 40 ферментов и белковых факторов, объединенных в единую ДНК-репликазную систему, называемую реплисомой.
1.Процесс биосинтеза ДНК.
Важную функцию соединения двух цепей ДНК или замыкания двух концов одной цепи ДНК в процессе репликации либо репарации ДНК выполняет особый фермент ДНК-лигаза, катализирующая за счет энергии АТФ образование фосфодиэфирной связи между 3'-ОН-группой дезоксирибозы одной цепи и 5'-фосфатной группой другой цепи ДНК.
Из клеток животных выделено несколько ДНК-полимераз, и в разных лабораториях они получили различные наименования.
К настоящему времени у эукариот, как и у бактерий (см. ранее), открыто несколько ДНК-полимераз. В репликации ДНК эукариот участвуют два главных типа полимераз – α и δ. Показано, что ДНК-полимераза α состоит из 4 субъединиц и является идентичной по структуре и свойствам во всех клетках млекопитающих, причем одна из субъединиц оказалась наделенной праймазной активностью. Самая крупная субъединица ДНК-полимеразы а (мол. масса 180000) катализирует реакцию полимеризации, преимущественно синтез отстающей цепи ДНК, являясь составной частью праймасомы. ДНК-полимераза δ состоит из 2 субъединиц и преимущественно катализирует синтез ведущей цепи ДНК (см. далее). Открыта также ДНК-полимераза ε, которая в ряде случаев заменяет δ-фермент, в частности при репарации ДНК (исправление нарушений ДНК, вызванных ошибками репликации или повреждающими агентами). Следует отметить, что в эукариотических клетках открыты два белковых фактора репликации, обозначаемых RFA и RFC. Фактор репликации А выполняет функцию белка – связывание одноцепочечной ДНК (наподобие белковых факторов связывания разъединенных цепей ДНК при репликации у Е. coli), фактор С – функцию стабилизатора всего реплика-ционного комплекса.
В генетической инженерии с целью получения белков в достаточных количествах и с заданными свойствами (например, для генотерапии наследственных и соматических болезней) широкое применение получили эндо-нуклеазы рестриктазы, катализирующие расщепление молекулы двух-цепочечной ДНК по специфическим нуклеотидным последовательностям внутри цепи. Рестриктазы узнают определенные 4–7-членные последовательности, вызывая, таким образом, разрывы в определенных сайтах цепи ДНК. При этом образуются не случайные последовательности, а фрагменты ДНК строго определенной структуры с липкими концами (ре-комбинантные ДНК), используемые далее для конструирования гибридных молекул и получения генно-инженерной, биотехнологической продукции (например, инсулина, гормона роста, интерферона, вакцин против вируса гепатита В, СПИДа и др.).
2. Общий механизм синтеза ДНК.

Химический смысл полимеризации состоит в том, что свободная 3'-гидроксильная группа матрицы атакует α-фосфатную группу соответствующего присоединяемого нуклеозидтрифосфата (определяется природой азотистого основания затравки), при этом происходят отщепление остатка пирофосфата и образование фосфодиэфирной связи. Далее свободный 3'-гидроксил вновь присоединенного нуклеотида атакует α-фосфатную группу следующего нуклеозидтрифосфата, и таким путем продолжается процесс полимеризации, идущий в направлении 5'–3', антипараллельно матрице, оканчивающейся 5'-фосфатом:

Реакция требует присутствия одноцепочечной ДНК или в крайнем случае небольшого полидезоксирибонуклеотида. В деталях выяснено значение предобразованной ДНК в механизмах действия ДНК-полимераз: ДНК служит не только затравкой, но и матрицей, на которой фермент комплементарно и антипараллельно синтезирует дочернюю цепь ДНК. Это можно представить в виде схемы:

Сложность процесса репликации ДНК объясняется тем, что обе цепи реплицируются одновременно, хотя имеют разное направление (5'–3' и 3'–5'); кроме того, рост дочерних цепей также должен происходить в противоположных направлениях. Элонгация каждой дочерней цепи может осуществляться только в направлении 5'–3'. Р. Оказаки высказал предположение, подтвержденное экспериментальными данными, что синтез одной из дочерних цепей осуществляется непрерывно в одном направлении, в то время как синтез другой дочерней цепи происходит прерывисто, путем соединения коротких фрагментов (в честь автора названы фрагментами Оказаки), в свою очередь синтезирующихся в противоположном направлении (рис. 13.4).
Как видно, синтез ведущей цепи ДНК идет всегда в направлении 5'–3', соответствующем направлению движения репликационной вилки. Сохраняя правило синтеза дочерних молекул ДНК 5'–3', синтез на второй цепи родительской ДНК идет в направлении, противоположном движению репликационной вилки. В зависимости от типа клетки фрагменты Оказаки имеют разные размеры – от нескольких сот до нескольких тысяч нуклеотидов (150–200 у эукариот и 1000–2000 у бактерий).

Рис. 13.1. Роль ДНК-полимеразы и ДНК-лигазы в синтезе кольцевой одноцепочеч-ной ДНК фага φХ174.
3. Особенности репликации ДНК у эукариот.

Рис. 13.3. Основные этапы репликации ДНК (схема).
Как было указано, инициация биосинтеза дочерних цепей ДНК требует предварительного синтеза на матрице ДНК необычного затравочного олигорибонуклеотида, названного праймером, со свободной гидроксильной группой у С-3' рибозы. Этот короткий олигорибонуклеотид синтезируется комплементарно на матрице ДНК при участии особого фермента – прай-мазы, наделенной РНК-полимеразной активностью.
Синтез начинается с реакции между 3'-ОН-группой концевого рибонуклеотида праймера и α-фосфатной группой первого дезоксирибонуклеотидтрифосфата в строгом соответствии с комплементарностью родительской цепи ДНК, при этом освобождается пирофосфат. В дальнейшем этот фрагмент РНК, комплементарно присоединенный к новообразованной цепи ДНК, разрушается под действием ДНК-полимеразы I, и возникшая брешь застраивается олигодезоксирибо-нуклеотидом при помощи той же ДНК-полимеразы I. Вполне допустимо предположение, что синтез праймера из олигорибонуклеотида имеет глубокий биологический смысл, поскольку в этом случае могут устраняться ошибки, неизбежно возникающие при инициации репликации ДНК.
4. Этапы биосинтеза ДНК.
Предложен ряд моделей механизма биосинтеза ДНК с участием указанных ранее ферментов и белковых факторов, однако детали некоторых этапов этого синтеза еще не выяснены. Основываясь главным образом на данных, полученных в опытах in vitro, предполагают, что условно механизм синтеза ДНК у Е. coli может быть подразделен на три этапа; инициацию, т.е. начало, элонгацию, т.е. продолжение, и терми-нацию, т.е. завершение (прекращение) синтеза. Каждый из этих этапов требует участия специфических ферментов и белковых факторов.
Этап I – инициация биосинтеза ДНК – является началом синтеза дочерних нуклеотидных цепей; в инициации участвует минимум восемь хорошо изученных и разных ферментов и белков. Первая фаза – это, как указано ранее, ферментативный биосинтез на матрице ДНК необычного затравочного олигорибонуклеотида (праймера) со свободной гидроксиль-ной группой у С-3' рибозы. При инициации к цепям ДНК последовательно присоединяются ДНК-раскручивающие и ДНК-связывающие белки, а затем комплексы ДНК-полимераз и праймаз (см. рис. 13.3). Инициация представляется единственной стадией репликации ДНК, которая весьма тонко и точно регулируется, однако детальные механизмы ее до сих пор не раскрыты и в настоящее время интенсивно исследуются.
Этап II – элонгация синтеза ДНК – включает два кажущихся одинаковыми, но резко различающихся по механизму синтеза лидирующей и отстающей цепей на обеих материнских цепях ДНК. Синтез лидирующей цепи начинается с синтеза праймера (при участии праймазы) у точки начала репликации, затем к праймеру присоединяются дезоксирибонуклеотиды под действием ДНК-полимеразы III; далее синтез протекает непрерывно, следуя шагу репликационной вилки. Синтез отстающей цепи, напротив, протекает в направлении, обратном движению репликационной вилки и начинается фрагментарно. Фрагменты всякий раз синтезируются раздельно, начиная с синтеза праймера, который может переноситься с готового фрагмента при помощи одного из белковых факторов репликации в точку старта биосинтеза последующего фрагмента противоположно направлению синтеза фрагментов. Элонгация завершается отделением олигорибонуклеотидных праймеров, объединением отдельных фрагментов ДНК при помощи ДНК-лигаз и формированием дочерней цепи ДНК. Нельзя исключить, однако, возможности сопряженного и согласованного механизма синтеза лидирующей и отстающей цепей ДНК при участии полимераз и всего комплекса праймасом.
Этап III – терминация синтеза ДНК – наступает, скорее всего, когда исчерпана ДНК-матрица и трансферазные реакции прекращаются. Точность репликации ДНК чрезвычайно высока, возможна одна ошибка на 1010 трансферазных реакций, однако подобная ошибка обычно легко исправляется за счет процессов репарации.
нуклеиновых кислот может меняться от сотен тысяч до десятков миллиардов.
Они были открыты и выделены из клеточных ядер ещё в 19 в., но их
биологическая роль выяснена только во второй половине 20 в.
Строение нуклеиновых кислот можно установить, анализируя продукты их
гидролиза. При полном гидролизе нуклеиновых кислот образуется смесь
пиримидиновых и пуриновых оснований, моносахарид (β - рибоза или β -
дезоксирибоза) и фосфорная кислота. Это означает, что нуклеиновые
При частичном гидролизе нуклеиновых кислот образуется смесь
нуклеотидов, молекулы которых построены из остатков фосфорной кислоты,
моносахарида (рибозы или дезоксирибозы) и азотистого основания
(пуринового или пиримидинового). Остаток фосфорной кислоты связан с 3-м
или 5-м атомом углерода моносахарида, а остаток основания – с первым
атомом углерода моносахарида. В зависимости от типа азотистого основания
Нуклеотид – основная структурная единица нуклеиновых кислот, их
мономерное звено. Нуклеиновые кислоты, состоящие из рибонуклеидов,
называют рибонуклеиновыми кислотами (РНК). Нуклеиновые кислоты,
состоящие из дезоксирибонуклеотидов, называют дезоксирибонуклеиновыми
кислотами (ДНК). В состав молекул РНК входят нуклеотиды, содержащие
основания аденин, гуанин, цитозин и урацил. В состав молекул ДНК входят
нуклеотиды, содержащие аденин, гуанин, цитозин и тимин. Для обозначения
оснований используют сокращения: аденин – А, гуанин – G, тимин – Т,
Свойства ДНК и РНК определяются последовательностью оснований в
полинуклеотидной цепи и пространственным строением цепи.
Последовательность оснований содержит генетическую информацию, а
остатки моносахаридов и фосфорной кислоты играют структурную роль
В молекулах ДНК и РНК отдельные нуклеотиды связаны в единую
полимерную цепь за счёт образования сложноэфирных связей между
остатками фосфорной кислоты и гидроксильными группами при 3-м и 5-м
Пространственная структура полинуклеотидных цепей ДНК и РНК была
определена методом рентгеноструктурного анализа. Одним из самых
крупных открытий биохимии 20 века оказалась модель двуспиральной
структуры ДНК, которую предложили в 1953 г. Дж. Уотсон и Ф. Крик.
Согласно этой модели, молекула ДНК представляет собой двойную спираль
и состоит из двух полинуклеотидных цепей, закрученных в
противоположные стороны вокруг общей оси. Пуриновые и пиримидиновые
основания расположены внутри спирали, а остатки фосфата и дезоксирибозы
– снаружи. Две спирали удерживаются вместе водородными связями между
парами оснований. Важнейшее свойство ДНК – избирательность в
образовании связей (комплементарность). Размеры оснований и двойной
спирали подобраны в природе таким образом, что тимин образует
водородные связи только с аденином, а цитозин – только с гуанином.
Таким образом, две спирали в молекуле ДНК комплементарны друг другу.
Последовательность нуклеотидов в одной из спиралей однозначно
определяет последовательность нуклеотидов в другой спирали.
В каждой паре оснований, связанных водородными связями, одно из
оснований – пуриновое, другое – пиримидиновое. Отсюда следует, что общее
число остатков пуриновых остатков в молекуле ДНК равно числу остатков
В отличие от ДНК молекулы РНК состоят из одной полинуклеотидной
цепи. Число нуклеотидов в цепи колеблется от 75 до нескольких тысяч, а
молекулярная масса РНК может изменяться в пределах от 2500 до
нескольких миллионов. Полинуклеотидная цепь РНК не имеет строго
ДНК – главная молекула в живом организме. Она хранит генетическую
информацию, которую передаёт от одного поколения к другому. В молекулах
ДНК в закодированном виде записан состав всех белков организма. Каждой
аминокислоте, входящей в состав белков, соответствует свой код в ДНК, т. е.
ДНК содержит всю генетическую информацию, но непосредственно в
синтезе белков не участвует. Роль посредника между ДНК и местом синтеза
белка выполняет РНК. Процесс синтеза белка на основе генетической
информации схематично можно разбить на две основные стадии: считывание
Клетки содержат три типа РНК, которые выполняют различные функции.
1) Информационная или матричная РНК (м-РНК) считывает и переносит
генетическую информацию от ДНК, содержащейся в хромосомах, к
рибосомам, где происходит синтез белка со строго определённой
2) Транспортная РНК (т-РНК) переносит аминокислоты к рибосомам, где
3) Рибосомная РНК (р-РНК) непосредственно участвует в синтезе белков в
рибосомах. Рибосомы – сложные надмолекулярные структуры, которые
состоят из четырёх р-РНК и нескольких десятков белков. Фактически
Последовательность оснований в м-РНК – это генетический код,
управляющий последовательностью аминокислот в белках. Замечательная
особенность генетического кода состоит в том, что он универсален для всех
Репликация- самовоспроизведение молекулы ДНК с целью передачи генетической информации. В репликации ДНК участвует сложный репликационный комплекс, который включает в себя:
- нуклеотиды в виде трифосфатов (АТФ, ГТФ, ТТФ, ЦТФ). Они являются одновременно и структурным материалом и источником энергии;
- ДНК-матрица – программа, по которой будет синтезироваться ДНК;
- праймер (затравка) - короткая молекула РНК, к которой присоединяется первый нуклеотид ДНК;
- ферменты:
- ДНК-полимеразы соединяют нуклеотиды в полинуклеотидную цепь,
- ДНК-рестриктазы разрывают полинуклеатидную цепь ДНК.
- ДНК-лигазы соединяют фрагменты ДНК,
- хеликазы раскручивают молекулу ДНК,
- топоизомеразы осуществляют кратковременные разрывы ДНК в процессе синтеза и контролируют конформацию вновь синтезированной ДНК,
- праймазы – осуществляют синтез затравки;
- белковые факторы;
- ионы металлов.
Репликация ДНК происходит в S-фазу клеточного цикла. Важную роль в чередовании циклов выполняют белки - циклины.
Сущность репликации сводится к соединению нуклеотидов в комплементарную антипараллельную дочернюю цепь ДНК. В репликации выделяют 3 стадии: инициация (начало синтеза), элонгация (удлинение цепи), терминация (окончание синтеза).
Матрицей для синтеза ДНК являются обе полинуклеотидные цепи материнской ДНК. Репликация начинается в нескольких участках материнской ДНК (точки репликации, точки Ориджины). В них происходит частичный разрыв водородных связей с формированием репликационных утолщений (узелков). В последующем, при участии ферментов ДНК-рестриктаз происходит разрезание молекулы ДНК. Под действием ферментов хеликаз возникает расплетание (раскручивание) двух цепей, и при участии топоизомераз формируется репликационная вилка, в которой одна цепь имеет направление 5→3, а вторая 3→5. Затем на каждой из ПНЦ синтезируются дочерние нити ДНК по принципу комплементарности. Поскольку ДНК- полимераза замыкает связь только в направлении 5’→3’, то на одной цепи ДНК происходит синтез непрерывной (лидирующей) нити ДНК в направлении 5→3. При синтезе лидирующей цепи праймазы синтезируют праймер, затем ДНК- полимераза присоединяет к праймеру первый ДНК-овый нуклеотид и по принципу комплементарности происходит удлинение молекулы ДНК. На второй материнской нити ДНК синхронно синтезируется отстающая нить ДНК, которая синтезируется в виде небольших фрагментов в направлении 5→3. В последующем эти фрагменты (фрагменты Оказаки) соединяются между собой ДНК-лигазами. Праймеры при этом расщепляются.
Таким образом, биосинтез ДНК проходит по полуконсервативному типу, при котором в новой ДНК одна цепь материнская, а другая - дочерняя.
Репарация ДНК происходит при появлении в её молекуле повреждений, искажений. При репарации вначале распознаётся место повреждения, затем ферменты рестриктазы вырезают дефектный участок, ДНК-полимеразы по принципу комплементарности синтезируют отсутствующий участок, а ДНК-лигазы прикрепляют его к сохранившимся участкам неповреждённой ДНК.
Биосинтез РНК (транскрипция)
Транскрипция – синтез молекулы РНК по матрице ДНК. Биологическая роль: перенос генетической информации с ДНК на РНК. Для транскрипции необходимы:
- матрица (программа) – кодирующая нить ДНК,
- субстраты – АТФ, ГТФ, УТФ, ЦТФ,
- ферменты – РНК-полимеразы,
- белковые факторы,
- ионы магния, марганца.
Выделяют 3 стадии транскрипции: инициация, элонгация, терминация
На молекуле ДНК имеется особый участок промотор, с которым связывается РНК-полимераза. Промотор иногда называют ТАТА участком (преобладает тимин, аденин, между которыми 2 Н-связи). Рядом с промотором расположены сигнальные участки, определяющие скорость транскрипции. Далее в молекуле ДНК располагаются кодирующие (экзоны) и некодирующие (интроны) участки гена. Участок (сайт) терминации определяет окончание синтеза РНК.
Инициация заключается во взаимодействии иницирующих белков с промотором и расхождении нитей ДНК, их раскручивании и формировании транскрипционной вилки. РНК-полимераза связывается с промоторным участком и по принципу комплементарности соединяет нуклеотиды цепи РНК в направлении от 5 конца к 3. РНК-полимераза - это олигомерный фермент, состоящий из нескольких субъединиц, не требующий затравки. При достижении РНК – полимеразой участка терминации происходит его связывание с белками терминации, что сопровождается отсоединением РНК-полимеразы от ДНК, диссоциацией её и окончанием транскрипции.
Процессинг РНК
Синтезированная РНК переписывает с кодирующей нити ДНК, как кодирующие участки, так и некодирующие участки гена и является про-РНК (незрелой РНК). Про-РНК в последующем подвергается созреванию (процессингу). Существует несколько механизмов процессинга:
- сплайсинг – вырезание копий интронов и соединение копий экзонов;
- присоединение к про-РНК добавочных нуклеотидов;
- модификация азотистых оснований в составе про-РНК
Особенности процессинга для рРНК, тРНК, иРНК.
Процессинг иРНК заключается в добавлении КЭП - участка и полиаденилового хвоста в сочетании со сплайсингом.
Процессинтг тРНК происходит путём метилирования азотистых оснований и добавления акцепторного участка ЦЦА в сочетании со сплайсингом.
Процессинтг рРНК заключается в вырезании из большого предшественника фрагментов всех видов РНК: 18S; 5S; 5,8S; 28S;.
Возможен альтернативный сплайсинг, который состоит в том, что для разных белков интроны могут играть роль экзонов.
Возможен и безматричный синтез РНК; он происходит из нуклеозиддифосфатов при участии фермента полинуклеотидфосфорилазы. В этом варианте синтезируются стандартные, небольшие молекулы РНК, необходимые для синтеза стандартных белков.
Таким образом, передача генетической информации происходит в следующем направлении: ДНК→ РНК→ белок. Однако, в некоторых фагах, эмбриональных тканях возможен синтез ДНК по матрице РНК (РНК→ДНК). Этот вариант синтеза катализирует фермент РНК-зависимая ДНК-полимераза (обратная транскриптаза, ревертаза). В вирусах возможен также вариант синтеза РНК→РНК при участии РНК-репликазы.
Читайте также: