
Генетический аппарат клетки реферат
Обновлено: 05.07.2024
Вы можете изучить и скачать доклад-презентацию на тему Генетический аппарат клетки. Презентация на заданную тему содержит 38 слайдов. Для просмотра воспользуйтесь проигрывателем, если материал оказался полезным для Вас - поделитесь им с друзьями с помощью социальных кнопок и добавьте наш сайт презентаций в закладки!

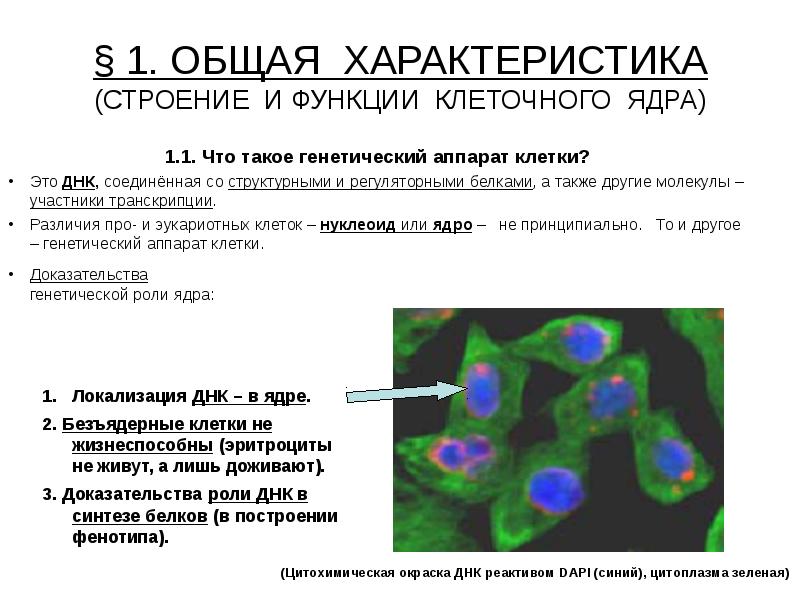

























§ 1. ОБЩАЯ ХАРАКТЕРИСТИКА (СТРОЕНИЕ И ФУНКЦИИ КЛЕТОЧНОГО ЯДРА) 1.1. Что такое генетический аппарат клетки? Это ДНК, соединённая со структурными и регуляторными белками, а также другие молекулы – участники транскрипции. Различия про- и эукариотных клеток – нуклеоид или ядро – не принципиально. То и другое – генетический аппарат клетки. Доказательства генетической роли ядра:
1.2. Общая морфология ядра эукариотной клетки 1.2. Общая морфология ядра эукариотной клетки Размер варьирует: объем ядра относительно объема цитоплазмы (Я/Ц отношение) = 1/2 - 1/10 - 1/100. Форма чаще округлая, бывает веретеновидная, лопастная. Локализация обычно в центре клетки или полярно, или под мембраной. Число ядер – обычно одно, бывает 2 и более, до нескольких сотен. Ядерный гигантизм – следствие полиплоидии.
Структурные компоненты ядра: Структурные компоненты ядра: 1. Ядерная оболочка – разобщение транскрипции и трансляции. 2. Ядерный матрикс (скелет) – пространственная организация. 3. Хроматин ↔ хромосомы – генетический материал. Концепция структурной целостности хромосом в клеточном цикле: хромосомы присутствуют в клетке всегда, но в разных структурно-функциональных состояниях. 4. Ядрышко – локус хроматина, производящий рРНК (рибосомы). 5. Кариолимфа (ядерный сок) – жидкая фракция ядра; содержит предшественники ядерных синтезов (нуклеотиды), белки-мигранты из цитоплазмы, продукты ядерных синтезов (мРНК, тРНК).
1.3. Геном 1.3. Геном Геном – гаплоидный набор генов биологического вида (человека, мыши…) Геном эукариот включает: информационную (генную) часть (хромосомную и внехромосомную): 1. Структурные гены – кодируют мРНК разнообразных структурных (рабочих) белков клетки. 2. Регуляторные гены – кодируют мРНК регуляторных белков. 3. Вспомогательные (РНКовые) гены – кодируют рРНК, тРНК и т.п.; неинформативные (сателлитные) области (функция – структурная? буферная? конъюгативная. ); транспозоны (подвижные генетические элементы) (функция – участие в репарации повреждений ДНК. ).
Размер генома – гаплоидная масса ДНК (с) – видоспецифичен: Размер генома – гаплоидная масса ДНК (с) – видоспецифичен: - Число молекул ДНК (число хромосом n) – от 1 до десятков и сотен. - Размер (масса) генома в целом увеличивается в ходе эволюции:
§ 2. СТРУКТУРНАЯ ОРГАНИЗАЦИЯ ХРОМАТИНА Длина 1 молекулы ДНК очень велика: - у бактерий кольцевая молекула ДНК = 1 – 2 мм, - у человека в разных хромосомах молекула ДНК = от 10 до 80 мм (всего 2с человека = около 1,8 м ДНК; 2с саламандры = 100 м!). Т.о., необходима упаковка (компактизация) ДНК – для работы и деления.
2.2. Структура хроматина (хромосом) эукариот Учитывать отличия организации генетического аппарата эукариот! В ядре содержится диплоидный (2n) набор хромосом, которые развернуты в рыхлую форму – хроматин. Состав хроматина = 40% ДНК и 60% белков = дезоксирибо-нуклео-протеид (ДНП). Белки хроматина: 1) Гистоны – оснóвные белки (богатые аргинином и лизином) – обеспечивают первичную упаковку и репрессию ДНК; 2) матриксины – нейтрально-кислые белки – обеспечивают высшие уровни упаковки ДНК; 3) регуляторные белки – изменяют связи ДНК и структурных белков. NB: эти связи регулируемые, изменяют активность генов.
Хроматин в световом микроскопе выглядит как хроматиновая сеть. Хроматин в световом микроскопе выглядит как хроматиновая сеть.
В электронном микроскопе эухроматин и гетерохроматин хорошо различимы как рыхлые и плотные участки ядра. В электронном микроскопе эухроматин и гетерохроматин хорошо различимы как рыхлые и плотные участки ядра.
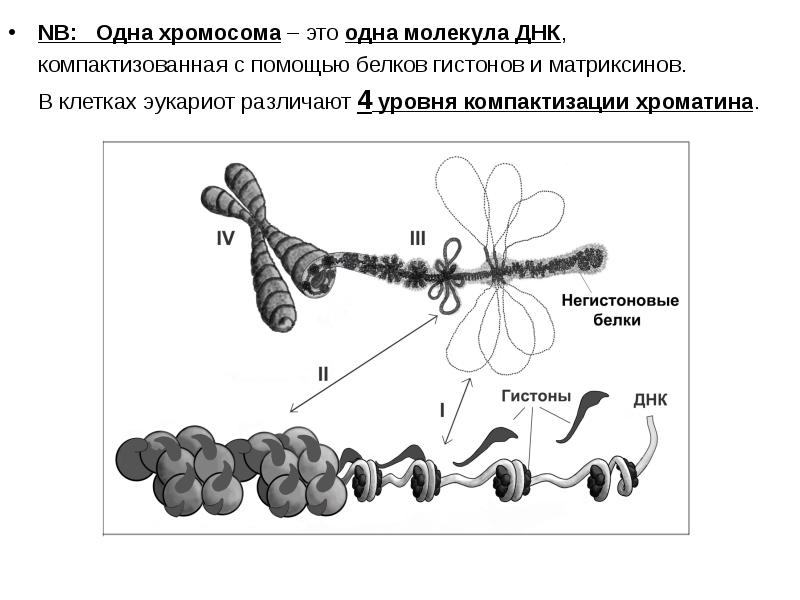
NB: Одна хромосома – это одна молекула ДНК, NB: Одна хромосома – это одна молекула ДНК, компактизованная с помощью белков гистонов и матриксинов. В клетках эукариот различают 4 уровня компактизации хроматина.
Третий уровень компактизации – хромонема – Третий уровень компактизации – хромонема – петельно-хромомерная фибрилла Ø 100-200 нм.
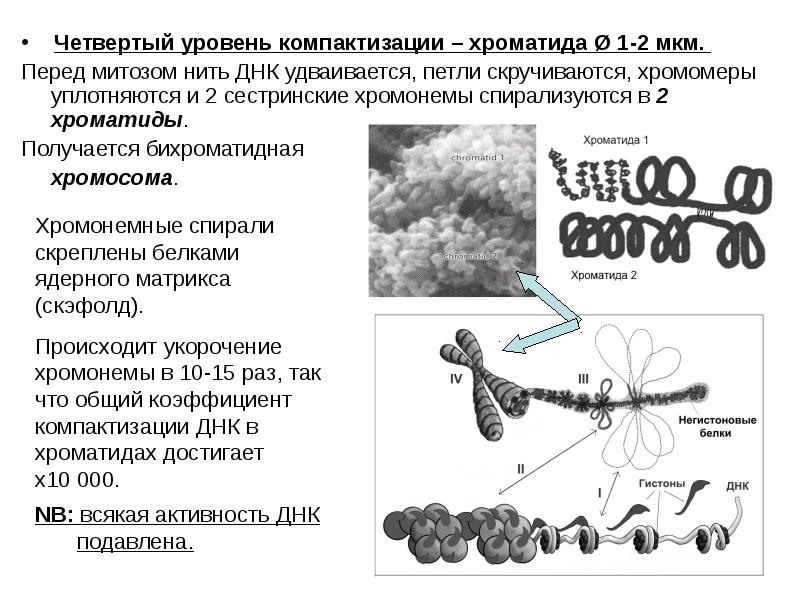
Четвертый уровень компактизации – хроматида Ø 1-2 мкм. Перед митозом нить ДНК удваивается, петли скручиваются, хромомеры уплотняются и 2 сестринские хромонемы спирализуются в 2 хроматиды. Получается бихроматидная хромосома.
§3. ХРОМОСОМНЫЙ НАБОР Каждый биологический вид имеет свой (видоспецифичный) хромосомный набор. Морфологическая характеристика хромосомного набора – кариотип. Графическое изображение кариотипа – идиограмма. Диплоидные (2n) хромосомные наборы: 1 – 22 хромосомы жабы Bufo arenarum; 2 – 140 хромосом ящерицыTupinambis teguixin; 3 – 12 хромосом чернушки Nigella orientalis (сем. лютиковые); 4 – 208 хромосом камчатского краба Paralithodes camtschatica; 5 – 46 хромосом человека Homo sapiens; 6 – 23 хромосомы прямокрылого насекомого Chromacris miles; 7 – 8 хромосом сложноцветного растения Hypochoeris tweedie. NB: Все рисунки – при одном увеличении.
При описании кариотипа используют следующие признаки: 1. Число хромосом. 2n у разных видов варьирует от 2 (аскарида) до 1000-1600. Наиболее типичные значения 2n = 10-50 (n = 5-25) . У человека 2n = 46. 2. Размер хромосом. Длина хромосом варьирует от 0,2 мкм (дрожжи) до 50 мкм (лилейные). У человека длина хромосом от 10 мкм (№ 1) до 2-3 мкм (№ 22).
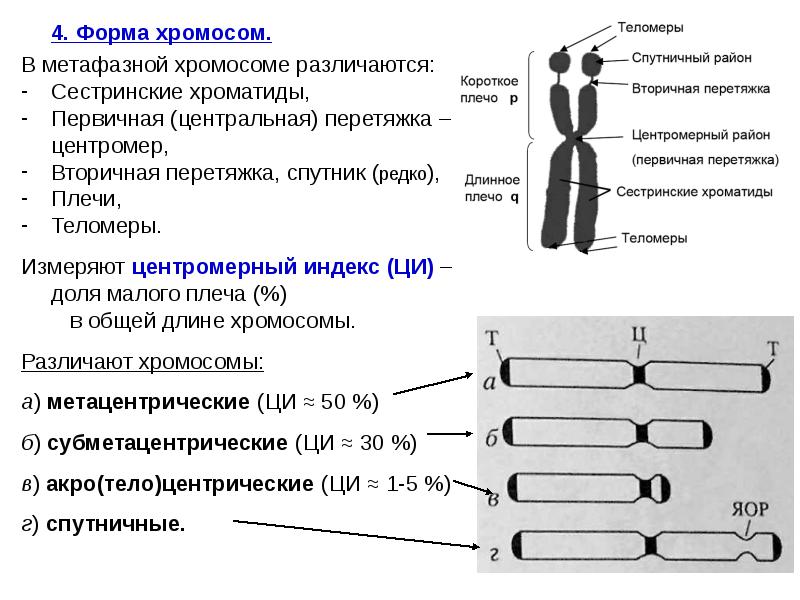
4. Форма хромосом. 4. Форма хромосом. В метафазной хромосоме различаются: Сестринские хроматиды, Первичная (центральная) перетяжка – центромер, Вторичная перетяжка, спутник (редко), Плечи, Теломеры. Измеряют центромерный индекс (ЦИ) – доля малого плеча (%) в общей длине хромосомы. Различают хромосомы: а) метацентрические (ЦИ ≈ 50 %) б) субметацентрические (ЦИ ≈ 30 %) в) акро(тело)центрические (ЦИ ≈ 1-5 %) г) спутничные.
5. Дифференциальная окраска хромосом. 5. Дифференциальная окраска хромосом. При специальных окрасках выявляются поперечные полосы, которые соответствуют разным фракциям ДНК: сателлитные (АТ-богатые), центромерные, ядрышковые и пр.
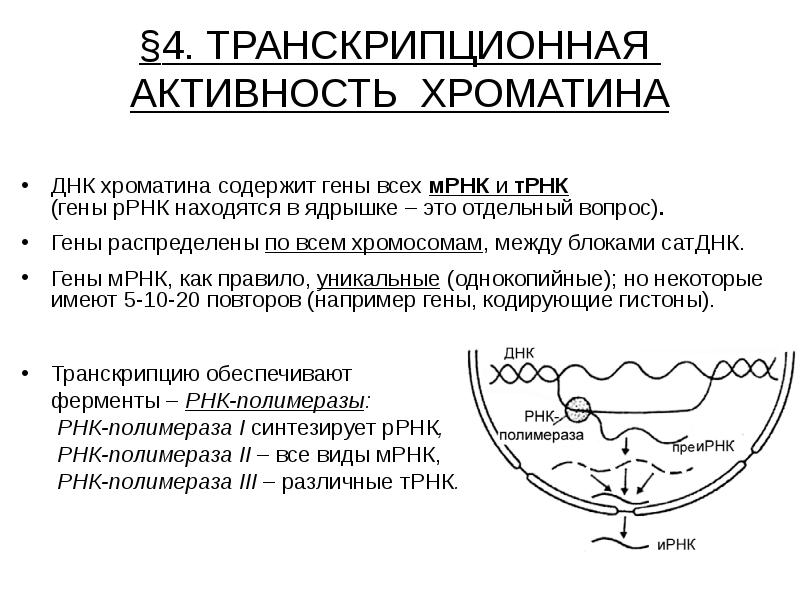
§4. ТРАНСКРИПЦИОННАЯ АКТИВНОСТЬ ХРОМАТИНА ДНК хроматина содержит гены всех мРНК и тРНК (гены рРНК находятся в ядрышке – это отдельный вопрос). Гены распределены по всем хромосомам, между блоками сатДНК. Гены мРНК, как правило, уникальные (однокопийные); но некоторые имеют 5-10-20 повторов (например гены, кодирующие гистоны). Транскрипцию обеспечивают ферменты – РНК-полимеразы: РНК-полимераза I синтезирует рРНК, РНК-полимераза II – все виды мРНК, РНК-полимераза III – различные тРНК.
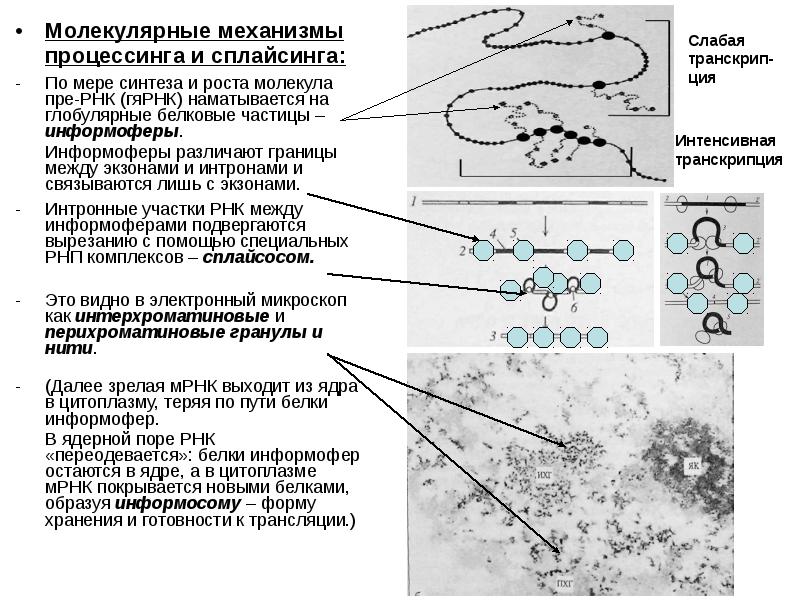
Активный ген транскрибируется одновременно несколькими молекулами РНК-полимеразы, что резко ускоряет синтез данного вида РНК в ядре. Активный ген транскрибируется одновременно несколькими молекулами РНК-полимеразы, что резко ускоряет синтез данного вида РНК в ядре. По ходу транскрипции (слева направо) видны растущие цепи РНК-транскриптов (пре-РНК). Транскрипция происходит в одном направлении. Ген (единица транскрипции) ограничен специальными участками ДНК (последовательностями Н-тидов) – сигналом инициации и сигналом терминации, которые распознаются РНК-полимеразой.
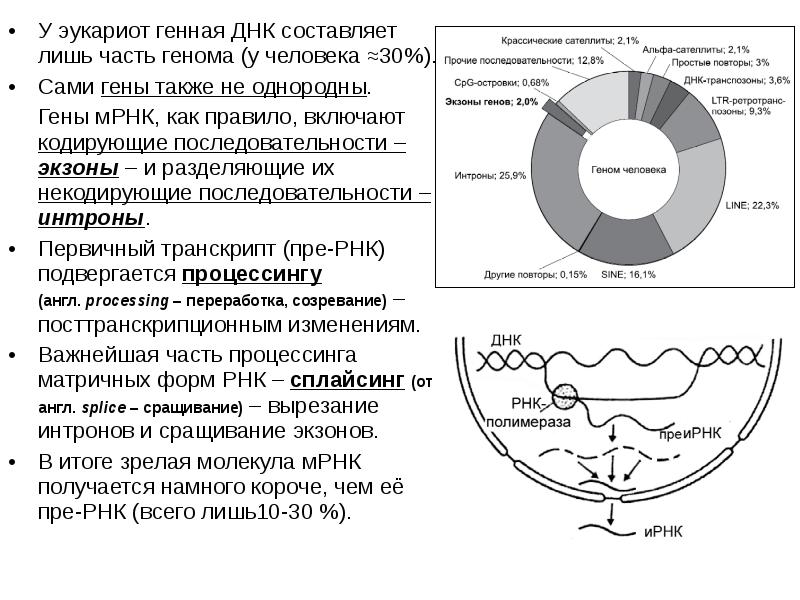
У эукариот генная ДНК составляет лишь часть генома (у человека ≈30%). У эукариот генная ДНК составляет лишь часть генома (у человека ≈30%). Сами гены также не однородны. Гены мРНК, как правило, включают кодирующие последовательности – экзоны – и разделяющие их некодирующие последовательности – интроны. Первичный транскрипт (пре-РНК) подвергается процессингу (англ. processing – переработка, созревание) –посттранскрипционным изменениям. Важнейшая часть процессинга матричных форм РНК – сплайсинг (от англ. splice – сращивание) – вырезание интронов и сращивание экзонов. В итоге зрелая молекула мРНК получается намного короче, чем её пре-РНК (всего лишь10-30 %).
Размер некоторых генов человека, производимых ими мРНК (в тысячах нуклеотидов) и число интронов в этих генах
В итоге с одного гена образуются частично различающиеся белки, например мембранная и внеклеточная формы фибронектина. В итоге с одного гена образуются частично различающиеся белки, например мембранная и внеклеточная формы фибронектина. (Лимфоциты продуцируют миллионы разновидностей антител (иммуноглобулинов) - белков, специфичных к чужеродным антигенам. Такое разнообразие специфичных белков образуется в результате альтернативного сплайсинга из немногих исходных генов.)
Модели изучения транскрипции – Модели изучения транскрипции – политенные хромосомы диптер хромосомы-ламповые щетки в мейотических ооцитах
§ 5. СТРУКТУРА И ФУНКЦИИ ЯДРЫШКА Рибосома состоит из 2 субчастиц, в которых рРНК выполняет роль структурной основы, организуя правильное расположение рабочих рибосомных белков. Косвенным показателем массы рибосом и макромолекул являются единицы Сведберга (S), которыми измеряется скорость их осаждения в центрифуге.
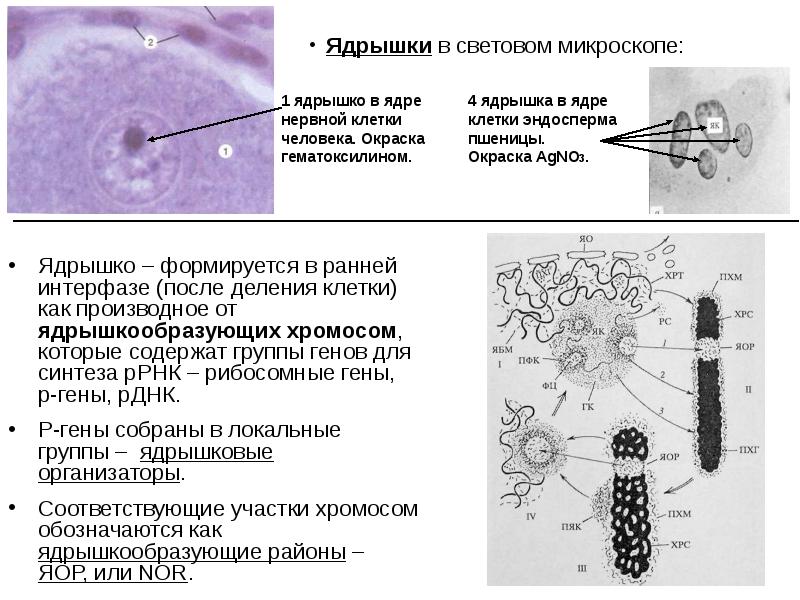
Ядрышко – формируется в ранней интерфазе (после деления клетки) как производное от ядрышкообразующих хромосом, которые содержат группы генов для синтеза рРНК – рибосомные гены, р-гены, рДНК. Ядрышко – формируется в ранней интерфазе (после деления клетки) как производное от ядрышкообразующих хромосом, которые содержат группы генов для синтеза рРНК – рибосомные гены, р-гены, рДНК. Р-гены собраны в локальные группы – ядрышковые организаторы. Соответствующие участки хромосом обозначаются как ядрышкообразующие районы – ЯОР, или NOR.
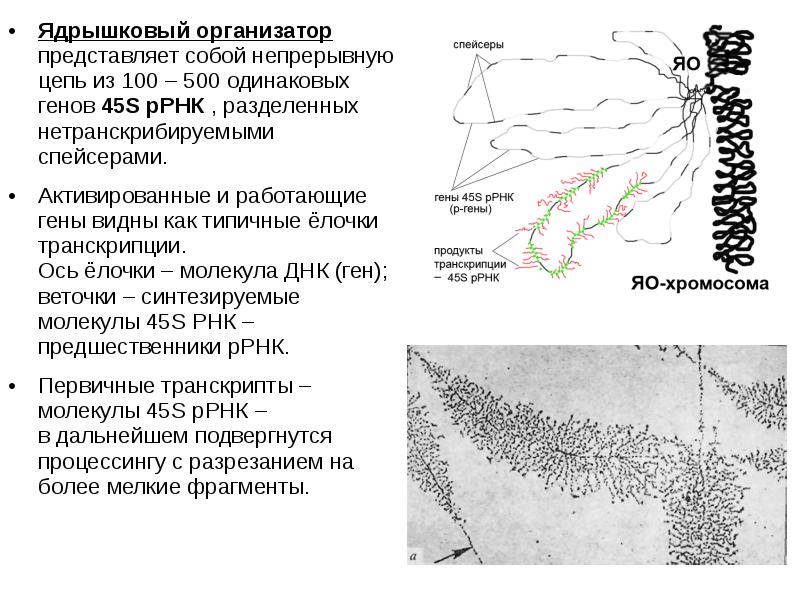
Ядрышковый организатор представляет собой непрерывную цепь из 100 – 500 одинаковых генов 45S рРНК , разделенных нетранскрибируемыми спейсерами. Ядрышковый организатор представляет собой непрерывную цепь из 100 – 500 одинаковых генов 45S рРНК , разделенных нетранскрибируемыми спейсерами. Активированные и работающие гены видны как типичные ёлочки транскрипции. Ось ёлочки – молекула ДНК (ген); веточки – синтезируемые молекулы 45S РНК – предшественники рРНК. Первичные транскрипты – молекулы 45S рРНК – в дальнейшем подвергнутся процессингу с разрезанием на более мелкие фрагменты.
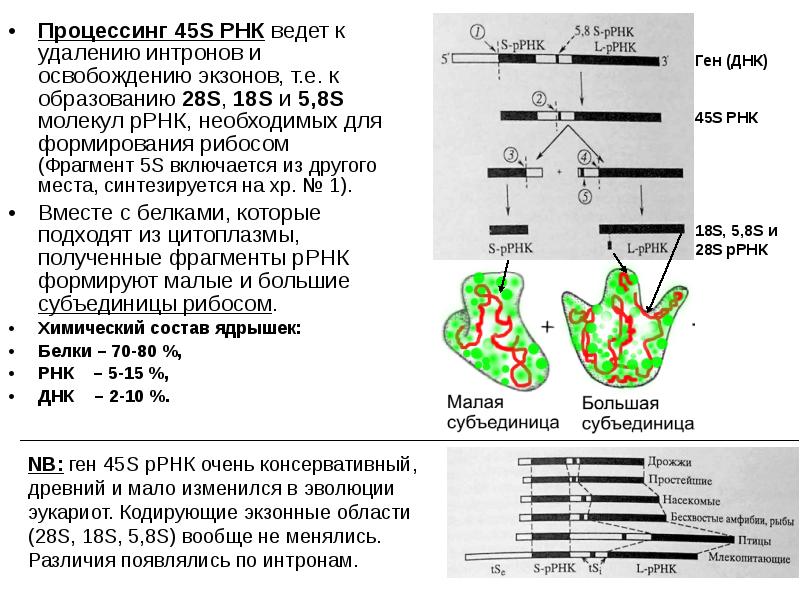
Процессинг 45S РНК ведет к удалению интронов и освобождению экзонов, т.е. к образованию 28S, 18S и 5,8S молекул рРНК, необходимых для формирования рибосом (Фрагмент 5S включается из другого места, синтезируется на хр. № 1). Процессинг 45S РНК ведет к удалению интронов и освобождению экзонов, т.е. к образованию 28S, 18S и 5,8S молекул рРНК, необходимых для формирования рибосом (Фрагмент 5S включается из другого места, синтезируется на хр. № 1). Вместе с белками, которые подходят из цитоплазмы, полученные фрагменты рРНК формируют малые и большие субъединицы рибосом. Химический состав ядрышек: Белки – 70-80 %, РНК – 5-15 %, ДНК – 2-10 %.
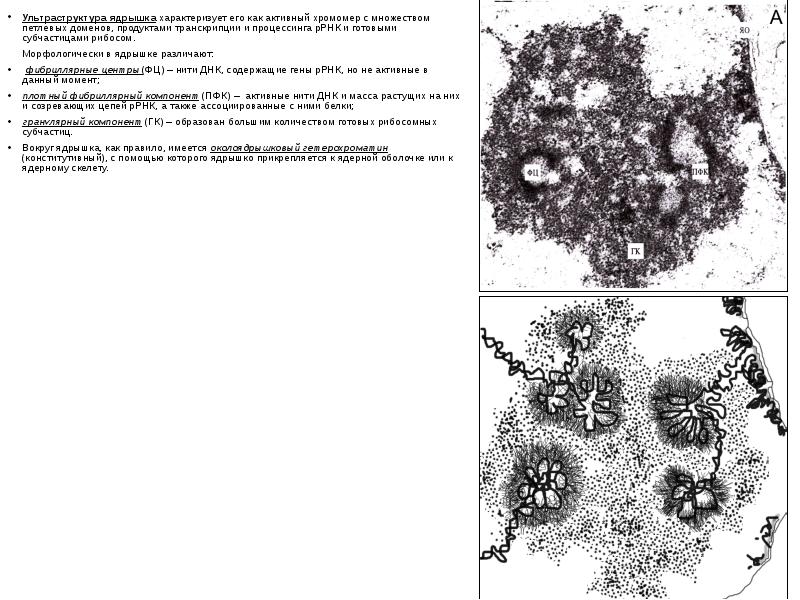
Ультраструктура ядрышка характеризует его как активный хромомер с множеством петлевых доменов, продуктами транскрипции и процессинга рРНК и готовыми субчастицами рибосом. Ультраструктура ядрышка характеризует его как активный хромомер с множеством петлевых доменов, продуктами транскрипции и процессинга рРНК и готовыми субчастицами рибосом. Морфологически в ядрышке различают: фибриллярные центры (ФЦ) – нити ДНК, содержащие гены рРНК, но не активные в данный момент; плотный фибриллярный компонент (ПФК) – активные нити ДНК и масса растущих на них и созревающих цепей рРНК, а также ассоциированные с ними белки; гранулярный компонент (ГК) – образован большим количеством готовых рибосомных субчастиц. Вокруг ядрышка, как правило, имеется околоядрышковый гетерохроматин (конститутивный), с помощью которого ядрышко прикрепляется к ядерной оболочке или к ядерному скелету.
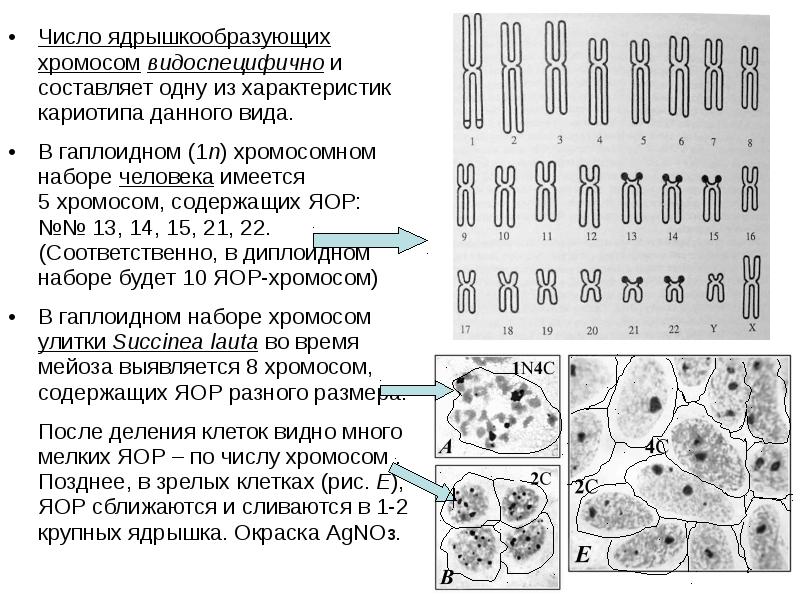
Число ядрышкообразующих хромосом видоспецифично и составляет одну из характеристик кариотипа данного вида. Число ядрышкообразующих хромосом видоспецифично и составляет одну из характеристик кариотипа данного вида. В гаплоидном (1n) хромосомном наборе человека имеется 5 хромосом, содержащих ЯОР: №№ 13, 14, 15, 21, 22. (Соответственно, в диплоидном наборе будет 10 ЯОР-хромосом) В гаплоидном наборе хромосом улитки Succinea lauta во время мейоза выявляется 8 хромосом, содержащих ЯОР разного размера. После деления клеток видно много мелких ЯОР – по числу хромосом . Позднее, в зрелых клетках (рис. Е), ЯОР сближаются и сливаются в 1-2 крупных ядрышка. Окраска AgNO3.
§ 6. ЯДЕРНАЯ ОБОЛОЧКА, МАТРИКС И ЯДЕРНО-ЦИТОПЛАЗМАТИЧЕСКИЕ ОТНОШЕНИЯ Ядерная оболочка обрамляет ядро эукариотной клетки. Нуклеоид прокариот оболочки не имеет. Оболочка ядра является центральной частью эндоплазматической сети, или ретикулума (ЭПС, ЭПР, ЭР), поэтому она образована двумя мембранами – внутренней и наружной, разделенными перинуклеарным пространством. Наружная мембрана переходит непосредственно в мембрану ЭПР, а перинуклеарное пространство сообщается с полостями каналов и цистерн ретикулума.
Ядерная оболочка имеет многочисленные поровые комплексы для транспорта веществ между ядром и цитоплазмой (мРНК, белки и пр.). Ядерная оболочка имеет многочисленные поровые комплексы для транспорта веществ между ядром и цитоплазмой (мРНК, белки и пр.). Ядерная пора – комплекс из >1000 белков нуклеопоринов. - центральная белковая глобула играет роль канала (транспортёр), - 8 гранул на наружной мембране, - два кольца и спицы фиксируют центральную глобулу. Все макромолекулы, проходящие через ядерные поры, имеют на переднем конце особую полипеп-тидную последовательность – сигнал ядерного импорта, которая распознается порой. После преодоления поры эта сигнальная последовательность отсоединяется от макромолекулы.
Читайте также: