
Эволюция мозга приматов реферат
Обновлено: 07.07.2024
Прикрепленные файлы: 1 файл
Этапы эволюции.rtf
На тему: Этапы эволюции приматов и человека.
Работу выполнил
студент группы Б-173:
Фёдоров Александр
1. Понятие об антропогенезе
В XVIII в. Зарождается и научная приматология. Так в 1766 году появилась научная работа Ж. Бюффона об орангутанге. Голландский анатом П. Кампер показал глубокое сходство в строении основных органов человека и животных.
В XVIII - первой половине XIX в. археологи, палеонтологи, этнографы накопили большой эмпирический материал, который лёг в основу учения об антропогенезе. Большую роль сыграли исследования французского археолога Ж. Буше де Перта.
Ж.Б. Ламарк первым писал, что человек произошёл от обезьяноподобных предков, перешедших от лазанья по деревьям к хождению по земле. Новый способ передвижения привёл к выпрямлению тела, освобождению рук и изменению стопы. Стадный образ жизни способствовал развитию речи. Однако даже Ламарк не решался довести до логического завершения идею эволюции животных и человека и отрицать роль Бога в происхождении человека.
Антропогенез не следует представлять в виде линейного процесса.
2. Человек как биологический вид
В эмбриональном развитии человека есть черты, характерные для всех представителей типа хордовых: хорда, нервная трубка, жаберные щели в глотке. Развитие позвоночного столба, развитие двух пар конечностей, расположение сердца на брюшной стороне тела определяют принадлежность человека к подтипу позвоночных. Четырёхкамерное сердце, сильно развитая кора головного мозга, теплокровность, молочные железы и другое свидетельствует о принадлежности человека к классу млекопитающих. Развитие плода в теле матери и питание его через плаценту характерно для класса плацентарных. Конечности хватательного типа (первый палец противопоставлен остальным), ногти на пальцах, одна пара сосков молочных желез, хорошо развитые ключицы и т.д. определяют положение человека в отряде приматов.
Таким образом, согласно зоологической систематике человека относят к царству животных, типу позвоночных, классу млекопитающих, отряду приматов, подотряду высших приматов, семейству гоминид (человекообразные), роду (homo), виду человек разумный (homo sapiens).
Чтобы понять что у нас общего с приматами, и что нас отличает от них, целесообразно проанализировать общие признаки приматов, среди которых можно найти и предпосылки для развития особенностей строения, физиологии и поведения человека.
2.1 Общая характеристика приматов
Приматов можно кратко охарактеризовать как группу лесных теплолюбивых плацентарных млекопитающих, приспособившихся к лазающему, древесному образу жизни, сохранив достаточно примитивную основу организации. Архаические черты в строении приматов проявляются в сохранении ими пятипалой конечности, опирающийся при ходьбе на всю ступню, сохранении ключиц, отсутствие специализации питания. Сохранение некоторых примитивных признаков и отсутствие специализации различных органов к какой - то одной функции обеспечивают высокую эволюционную пластичность приматов, проявивших в ходе филогенеза способность адаптироваться к самым различным условиям существования и способам использования природных ресурсов.
Важнейшие особенности приматов связаны с развитием приспособлений к древесной, лазающей жизни. Прежде сего это связано с совершенствованием хватательной способности конечностей сопровождающуюся приобретением возможности вращения кисти и предплечья вокруг их продольной оси, а также противопоставление большого пальца всем остальным, что значительно облегчает хватание.
Лазание по веткам требует хорошо развитого осязания (для исследования поверхности опоры), в связи с чем на концевых фалангах пальцев усиленно развиваются мясистые пальцевые подушечки и когти начинают преобразовываться в ногти.
Но самую важную роль среди органов чувств при лазании приобретают глаза, поскольку прыжки с ветки на ветку требуют возможности объёмного (стереоскопического) зрения, с точной оценкой расстояния и надёжности опоры. Развитие стереоскопического зрения достигается перемещением глаз на переднюю (лицевую) поверхность головы, их сближением и всё большим перекрыванием полей зрения
Вместе с тем при жизни на деревьях падает роль обоняния.
У приматов начинает совершенствоваться центральная нервная система - прогрессивное развитие головного мозга, особенно коры больших полушарий. Размер головного мозга увеличился, что сказалось на пропорциях черепа, в котором относительно увеличилась мозговая коробка и уменьшался челюстной отдел.
Кочевая жизнь на деревьях, которую ведёт большинство обезьян, не позволяет самкам иметь одновременно много детёнышей. Поэтому появляется тенденция к уменьшению числа детёнышей до 1-2 за одну беременность. Малочисленное потомство развило сложные формы родительского поведения, обеспечивающие длительный уход за детёнышами и защиту их родителями. Результатом уменьшения числа детёнышей была и редукция числа млечных желез до единственной (грудной) пары.
2.2 Сходство человека с приматами
Известно сходство человека и обезьяны в строении зубов, органов зрения, системы размножения, в частности наличия уникальных половых циклов. Изумительно похожи узоры пальцев, ладони, стопы (дерматоглифика); даже облысение у обезьян совместимо с облысением обычного типа у мужчин.
Гориллу и шимпанзе сближают с человеком группы крови системы АВО, антиген резус - фактор найден как у человека, так и у низшей обезьяны. У человека такой же физиологический механизм биологического старения.
Наглядны результаты исследования ДНК: у человека и шимпанзе 92% общих, сходных по первичной структуре генов, у человека и гиббона - 76%.
На общность мозга человека и животных указывает их структура: ископаемые останки самых ранних из известных позвоночных свидетельствуют о том, что основное деление мозга на задний, средний и передний существовало уже несколько сот миллионов лет назад. И мозг человека в сравнении с мозгом обезьяны не содержит ни одного типа клеток, тканей или органов, к тому же отдельные его части имеют аналогичные пропорции.
Подсчитано, что у человека и шимпанзе 396 общих анатомических признаков, у человека и гориллы - 385, у человека и орангутанга - 359.
Самым человекообразным из всех приматов считают карликового бонобо.
Итак, если всех млекопитающих расположить в один ряд от более простых к более сложным, то в конце этого ряда самым совершенным по строению, самым универсальным по своим возможностям окажется человек, а наиболее яду будут человекообразные обезьяны.
2.3 Различие человека и приматов
Человек имеет свои особенности, которые качественно отличают его от животных, в том числе и наших ближайших родственников - человекообразных обезьян.
3. Наши предки: кто они?
Антропологи спорят о том, кто из обезьян - шимпанзе, бонобо или гориллы - ближе к человеку, и о том, что послужило толчком к уникальным морфологическим и поведенческим преобразованиям: развитию двуногости, эволюцию кисти, увеличению головного мозга, становлению орудийной деятельности, речи, сознания. Нет окончательной ясности в понимании пути социальной эволюции человека.
Однако наука хотя и медленно, но последовательно заглядывает всё дальше в глубь времён.
Как выглядит цепочка предшественников современного человека?
Расхождение филогенетических стволов, ведущих к двум семействам антропоморфов, т.е. человекообразным и людям (гоминидам), произошло по разным оценкам, от 15 до 25 млн.лет назад. Ископаемые останки древесных обезьян дриопитеков по основным особенностям строения и размерам тела напоминали современных шимпанзе. Многие антропологи считают дриопитеков возможными непосредственными предками всех высших антропоморфов.
Высшего рассвета антропоморфные обезьяны достигли в неогене. Вероятно, древнейшие человекообразные обезьяны обитали в разреженных лесах на границах лесных массивов и лесостепей. В этих пограничных местообитаниях и могла произойти дивергенция понгид и гоминид: первые остались лесными обитателями, предки же вторых перешли к освоению всё более открытых ландшафтов. Предпосылки для этого заключались, во - первых, в переходе к наземности; во - вторых, во все большем использовании различных орудий для защиты и нападения и, соответственно, в освобождении рук от участия в передвижении; в - третьих, в необходимом совершенствовании двуногой локомоции.
Мозг приматов отличается от мозга других млекопитающих наибольшим развитием и дифференцированностью неокортекса (новой коры), составляющего у них 96% всей коры. Рисунок борозд характерный для приматов: вертикальная направленность центральных и тенденция к горизонтальному ходу других борозд.
Изучение строения мозга приматов в сравнительноанатомическом ряду позволило выявить два основные направления в его развитии. Первое отражает общую тенденцию развития от низших форм приматов к высшим. Оно выявляет закономерности усложнения мозга в соответствии с повышением уровня их общей организации и совершенствованием регуляторных механизмов.
Общее направление развития мозга включает шесть этапов.
Первый этап — мозг самых примитивных приматов (тупайя, галаго, лемуры) (рис. IV. 1). Характерный признак — ведущее развитие аллокортекса, т. е. древней старой и промежуточной коры). Слабо дифференцированный неокортекс расположен в дорсальной части и отделен от алло-кортекса обонятельной бороздой. Имеются четыре слабо дифференцированные и малообособленные цито-архитектонические области, соответствующие четырем долям мозга и корковым зонам (представительствам) сенсорных систем: зоны слуха, зрения, кожной чувствительности и движения. Значительное развитие аллокортекса сближает этих приматов с низшими представителями других млекопитающих. У более высокоорганизованных приматов аллокортекс уменьшается и вытесняется неокортексом с боковой поверхности мозга на базальную и медиальную.
Наиболее примитивен мозг тупайи: максимальное развитие аллокортекса и минимальное — неокортекса. Некоторые галаго имеют несколько прогрессивных черт, в частности, у них появляется сильвиева борозда. Среди лемуров встречается небольшое разрастание височной доли и зачаток сильвиевой борозды. У некоторых долгопят заметна еще одна прогрессивная особенность: височный полюс опускается ниже горизонтали остальной части полушария, затылочная область надвигается на мозжечок, образуется шпорная борозда. Второй этап — мозг некоторых представителей полуобезьян — лемуровых, а также игрунковых — представителей обезьян Нового Света. Для них характерен интенсивный рост височной доли, однако борозды в ней отсутствуют и дифференцировка на цитоархитектонические поля появляется лишь у отдельных представителей. Эта доля спереди четко отграничена сильвиевой бороздой от нижнелобной области, а сзади — теменно-затылочной зарубкой от затылочной. Вся височная доля выпячивается, расширяется и становится крупнее других. Основные поля располагаются по приматному типу. Появляются орименты филогенетически более молодых нижнетеменных полей. На медиальной поверхности полушарий мозга выражены теменно-затылочная и шпорная борозды. Затылочная доля сильно нависает и над мозжечком. У некоторых представителей появляются зачатки верхней височной, интрапариетальной, принципальной и фронтоорбитальной борозд. Появляется также дифференцировка полей в височной доле. Сильвиева борозда поднимается высоко и сливается с интрапариетальной. У некоторых представителей появляются, с одной стороны, отдельные черты более прогрессивной организации (образование лунной борозды в результате развития затылочной доли), с другой — выявляются признаки сходства с мозгом низшего ранга млекопитающих — хищных (фрагменты дугообразного расположения борозд вокруг сильвиевой борозды).
В целом, второй этап характеризуется разрастанием неокортекса по приматному типу (форма полушария, доли, области и поля), более четким делениям на области, разделяемые бороздами, и началом деления на цитоархитектонические поля. Тип организации второго этапа, вероятно, обусловлен адаптацией животных к ночному и сумеречному образу жизни, обитанию в тенистых лесах и др. В этих условиях увеличивается значение слухового и обонятельного анализаторов, которые начинают играть роль интегративного аппарата в организации целостных поведенческих реакций. Намечается также переход ведущей роли в нервной деятельности от височной к центральной сенсомоторной и затылочной областям.
Третий этап общего направления развития мозга приматов отражает мозг обезьян Нового Света — собственно лемуровых, ревунов, капуциновых, низших обезьян Старого Света (рис. IV. 1, 2). Для этого этапа характерно формирование питекоидного типа мозга с прогрессивным развитием центральных соматотопических сенсомоторных зон и затылочной области. Наблюдается отграничение центральной бороздой прецентральной (агранулярной) двигательной области от теменной сенсорной. Дугообразная борозда отделяет лобную область. Теменная доля четко отделяется от прецентральной,

Рис. IV. 1. Цитоархитектонические карты мозга полуобезьян и низших обезьян. 1 — перохвостая тупайя, 2 — серый гапалемур, 3 — зеленая мартышка
Условные обозначения
Sу. — Сильвиева борозда, с.е. — Центральная борозда, ip. — Межтеменная борозда, lun. — Лунная борозда, рг. — Принципиальная главная борозда, rh. — Ринальная борозда, t.s. — Верхняя височная борозда, arc. — Дугообразная борозда

Рис. IV. 2. Цитоархитектоническая карта мозга обезьян Нового Света и высших человекообразных обезьян. 1 — коата, 2 — капуцин, 3 — орангутан, 4 — гиббон, 5 — шимпанзе, 6 — горилла
Условные обозначения к рис. IV.1, IV2, IV.3, IV.4, IV5, IV.6 Цифры обозначают цнтоархитектонические поля коры по карте Бродмана; 17,18 — поля, связанные с осуществлением зрительных рецепций; 19 — ассоциативное зрительное поле; 4, 6 — двигательные поля; 1-3 — тактильная чувствительность; 41, 42 — слуховые поля; 22 — ассоциативно-слуховое; 43 — вкусовое; 39, 40 — нижнетеменные поля, связанные со стереоскопией и стереогнозией, праксис и манипулирование; 37 — зрительно-гностические функции; 44, 45 — речедвигателъные поля; 8 — глазодвигательное поле; 9, 10, 47 — поля лобной области, связанные с прогнозированием и высшими формами психической деятельности; 23, 24 — лимбические поля, связанные с эмоциями, пищевым, питьевым и половым поведением. Крупно-зернистая штриховка означает наиболее прогрессивно развивающиеся поля неокортекса
Выделяется мозг руконожки мадагаскарской. Она обгоняет других полуобезьян по развитию центральной борозды, которая у нее выражена даже лучше, чем у ряда обезьян. Вероятно, такое значительное развитие сенсомоторной зоны объясняется исключительно тонкой перкуссионной функцией третьего пальца кисти. Руконожки используют этот палец как молоточек, которым постукивают по коре дерева в поисках насекомых, живущих под корой, а также для доставания личинок из коры и древесины.
Своеобразен также мозг обезьян Нового Света. У капуцинов отмечается сходство с антропоморфными обезьянами по развитию филогенетически новой нижнетеменной области, новых подполей затылочной и дифференцировке подполей прецентральной области. С развитием этих областей связывают исключительную способность капуцинообразных к манипулированию предметами, и выраженной конструктивной деятельности. Капуцины отличаются также более сложными общими поведенческими реакциями по сравнению с другими представителями низших обезьян Нового и Старого Света.
У паукообразных обезьян развивается и распространяется в сторону затылочной задняя часть теменной доли. Все переходные извилины Грациоле открыты и заняты теменными формациями. В связи с прогрессивным развитием задней теменной, передней затылочной и височной областей становится разнообразным использование хватательных конечностей и цепкого хвоста. Согласованность мышечно-суставной чувствительности, вестибулярного и зрительного анализаторов достигает наивысшего уровня в специфической ориентации при подвешивании обезьян на ветвях в разных положениях — вверх и вниз головой. В результате у цепкохвостых обезьян тип строения мозга ближе к понгидному типу (тип высших человекообразных обезьян), нежели к типу низших обезьян Старого Света.
Таким образом, на третьем этапе особое значение имеет затылочный и центральный сенсомоторных комплекс полей. Он осуществляет регуляторные взаимодействия между гностическими, кинестетическими и зрительными функциями. Ведущую роль на третьем этапе играет стереоскопия и стереогнозия.
Четвертый этап (см. рис. IV. 2, 3-6) представляет мозг высших человекообразных обезьян — шимпанзе и гориллы. Он имеет следующие особенности: понгидный тип организации мозга; высший, приматный тип структуры борозд; затылочная покрышка уменьшается; теменно-затылочные извилины открыты и заняты теменными формациями; нижнетеменная область увеличена.
У самого примитивного представителя человекообразных обезьян — гиббона — мозг находится в состоянии, как бы переходном к антропоморфному типу. Для него характерно преобразование прецентральной моторной зоны. Нижняя вертикальная ветвь трехлучевой дугообразной борозды переходит в нижнюю прецентральную борозду. Принципальная борозда превращается в нижнюю лобную. Однако состояние нижнетеменной области остается на уровне низших обезьян. Особенности строения мозга соответствуют способам локомоции гиббонов, хотя они могут ходить по земле на двух ногах, основной способ их передвижения — это брахиация: уцепившись длинными руками за ветвь, они сильно раскачиваются, а затем мгновенно перебрасывают тело на большое расстояние на другую ветвь, затем — на следующую и т. д. Манипулирование развито слабо.
Для орангутана характерно дальнейшее развитие прецентральной области, с сильным ветвлением и своеобразием борозд. Вероятно, это связано с развитием брахиаторного типа локомоции. Заметно расширение нижнетеменных полей и зернистый тип цитоархитектоники, что, возможно, коррелирует с древесным образом жизни.
У шимпанзе рельеф борозд лобной области приближается к гоминидному типу. Это соответствует полувыпрямленному положению тела и полуназемному образу жизни. По дифференцировке полей нижнетеменной дольки, стратификации и ширине слоев, а также пирамидизации нейронов мозг шимпанзе приближается к человеческому типу. Эти черты соответствуют высокому уровню психических способностей шимпанзе.
Мозг гориллы характеризуется наибольшим усложнением борозд лобной доли при специфическом, понгидном типе их ветвления. По некоторым признакам, горилла стоит ближе других человекообразных обезьян к человеку.
Пятый этап нашел отражение в мозге ископаемых гоминид — австралопитека, питекантропа,неандертальца (рис. IV. 3). Это — гоминидный тип мозга с прогрессивным развитием филогенетически новых полей: нижнетеменной, верхневисочной и лобной областей. У сильвиевой борозды появляются две ветви — восходящая и горизонтальная, которые формируют комплекс речедвигательного центра. Из наплыва нижнелобной, теменной и височной коры образуется покрышка, погружающая островок. Заметно увеличение и усложнение интрасулькального компонента в составе неокортекса.
Для мозга австралопитека характерно формирование гоминидного типа полушария. Выявляются особые выпячивания в нижнетеменной, а именно в ангулярной (область зрительного гнозиса) и супрамаргинальной (зона мануального праксиса) извилинах. Имеются выпячивания также и в оперкулярной и лобной областях.
У питекантропов появляются выпуклости на эндокранах в нижнетеменной, задневисочной и нижней лобной областях, свидетельствующие о нарастании здесь массы мозга. По мере надвигания участков покрышек лобных, центральных и теменных долей, инсулярные формации оказываются полностью на дне сильвиевой ямы.
У неандертальцев разрастание нижнетеменной, верхней височной и нижнелобной областей усиливается. Это ведет к новому нарастанию покрышек островка и образуется зияющая сильвиева яма. На эндокранах очаги интенсивного роста в нижнетеменной, нижнелобной и височной областях расширяются и сливаются между собой. Рельеф полушария делается более обтекаемым и приближается по своим очертаниям к форме эндокрана у современных людей. Они не имеют больших выпячиваний и западений.

Рис. IV. 3. Топография корковых полей на эндокране австралопитека (А) и синантропа (Б)
Штриховкой обозначены места дефектов, пунктиром — отпечатки сосудов.
R — проекция центральной борозды; со — коронарный шов; L — лямбовидный шов; Ра — возвышение теменное угловое; Fi -возвышение лобное нижнее; М — возвышение прецентральное; Т — возвышение височное; f. Sy — сильвиева яма
Шестой этап — мозг современного человека не имеет себе равных по сложности и многообразию вариантов строения, форме и величине. Специфичность мозга человека проявляется в структурно-количественных соотношениях различных отделов, а также клеточной, волокнистой, нейронной и синаптической организации (рис. IV. 4).
Мозг человека отличается от мозга других приматов по следующим признакам:
1. Максимальное значение весового квадратного указателя (у грызунов — 0,19, у хищных — 1,14, у дельфинов — 6,27, у человекообразных обезьян — 7,35, у слона — 9,82, у человека — 32,0).
2. Максимальное преобладание головного мозга над спинным.
3. Максимальное преобладание новой коры над старой.
4. Резко увеличена площадь филогенетически более молодых отделов: например, площадь нижнетеменной коры в 37 раз выше, чем у обезьян.
5. В лобной доле у человека развиты две передние ветви сильвиевой борозды: восходящая и горизонтальная, разделяющие нижнюю лобную извилину на три: оперкулярную, триангулярную и орбитальную; на двух последних находятся рече-двигательные поля 44 и 45. В целом, размер площади коры и сложность ее микроскопической организации в филогенетически новых формациях достигают своих максимальных значений именно у человека.
6. Особая глубина и сложность ветвлений борозд полушария, появляющаяся после рождения и придающая мозгу индивидуальный рисунок. У человека до 75% коры находится в глубине борозд.
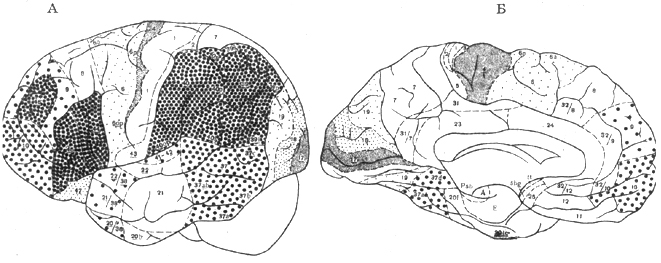
Рис. IV. 4. Цитоархитектоническая карта мозга современного человека (по Бродману) а — дорсолатеральная поверхность; Б — медиальная поверхность
8. Качественные отличия мозга человека от мозга животных, делающие его уникальной материей, имеют в своей основе морфологический субстрат — специфически человеческую морфофункциональную систему. Именно СЧМФС является морфологической основой высшей психической деятельности человека. Она представлена филогенетически новыми полями неокортекса, возникшими в процессе антропогенеза(поля лобной области, теменной, височно-теменно-затылочной подобласти), мощной системой пучков ассоциативных волокон и сосудами бассейна средней мозговой артерии, индуцирующими их развитие и питающими их. СЧМФС включает также специфическую для человека систему членораздельной речи, которая представлена 4 центрами: рече-двигательным, рече-слуховым, рече-зрительным и центром письменной речи.
9. Для коры мозга человека характерна наивысшая степень дифференцированности и сложности колончатой организации, особенно в специфических для него отделах.В процессе эволюции увеличилось количество ассоциативных корковых нейронов, степень ветвлений дендритов и число шипиков на них, число и пластичность синапсов, возросла эффективность медиаторных процессов и ферментных систем мозга.
Все это способствовало образованию новых межнейронных связей и создало предпосылки для совершенствования двигательной и манипуляторной деятельности, интенсификации механизмов памяти и обучения, развития интеллекта, прогнозирования будущих событий и др.
В результате мозг современного человека представляет собой уникальный орган, наивысший продукт эволюции.
В этот день:
Дни рождения 1961 Родился Владимир Роальдович Эрлих — советский и российский учёный-историк, доктор исторических наук, специалист по скифской эпохе Северного Кавказа.
Мозг приматов отличается от мозга других млекопитающих наибольшим развитием и дифференцированностью неокортекса (новой коры), составляющего у них 96% всей коры. Рисунок борозд характерный для приматов: вертикальная направленность центральных и тенденция к горизонтальному ходу других борозд.

Изучение строения мозга приматов в сравнительноанатомическом ряду позволило выявить два основные направления в его развитии. Первое отражает общую тенденцию развития от низших форм приматов к высшим. Оно выявляет закономерности усложнения мозга в соответствии с повышением уровня их общей организации и совершенствованием регуляторных механизмов.
Общее направление развития мозга включает шесть этапов.
Первый этап — мозг самых примитивных приматов (тупайя, галаго, лемуры) (рис. IV. 1). Характерный признак — ведущее развитие аллокортекса, т. е. древней старой и промежуточной коры). Слабо дифференцированный неокортекс расположен в дорсальной части и отделен от алло-кортекса обонятельной бороздой. Имеются четыре слабо дифференцированные и малообособленные цито-архитектонические области, соответствующие четырем долям мозга и корковым зонам (представительствам) сенсорных систем: зоны слуха, зрения, кожной чувствительности и движения. Значительное развитие аллокортекса сближает этих приматов с низшими представителями других млекопитающих. У более высокоорганизованных приматов аллокортекс уменьшается и вытесняется неокортексом с боковой поверхности мозга на базальную и медиальную.

Изменения черепа примата до формирования человека
Наиболее примитивен мозг тупайи: максимальное развитие аллокортекса и минимальное — неокортекса. Некоторые галаго имеют несколько прогрессивных черт, в частности, у них появляется сильвиева борозда. Среди лемуров встречается небольшое разрастание височной доли и зачаток сильвиевой борозды. У некоторых долгопят заметна еще одна прогрессивная особенность: височный полюс опускается ниже горизонтали остальной части полушария, затылочная область надвигается на мозжечок, образуется шпорная борозда. Второй этап — мозг некоторых представителей полуобезьян — лемуровых, а также игрунковых — представителей обезьян Нового Света. Для них характерен интенсивный рост височной доли, однако борозды в ней отсутствуют и дифференцировка на цитоархитектонические поля появляется лишь у отдельных представителей. Эта доля спереди четко отграничена сильвиевой бороздой от нижнелобной области, а сзади — теменно-затылочной зарубкой от затылочной. Вся височная доля выпячивается, расширяется и становится крупнее других. Основные поля располагаются по приматному типу. Появляются орименты филогенетически более молодых нижнетеменных полей. На медиальной поверхности полушарий мозга выражены теменно-затылочная и шпорная борозды. Затылочная доля сильно нависает и над мозжечком. У некоторых представителей появляются зачатки верхней височной, интрапариетальной, принципальной и фронтоорбитальной борозд. Появляется также дифференцировка полей в височной доле. Сильвиева борозда поднимается высоко и сливается с интрапариетальной. У некоторых представителей появляются, с одной стороны, отдельные черты более прогрессивной организации (образование лунной борозды в результате развития затылочной доли), с другой — выявляются признаки сходства с мозгом низшего ранга млекопитающих — хищных (фрагменты дугообразного расположения борозд вокруг сильвиевой борозды).
В целом, второй этап характеризуется разрастанием неокортекса по приматному типу (форма полушария, доли, области и поля), более четким делениям на области, разделяемые бороздами, и началом деления на цитоархитектонические поля. Тип организации второго этапа, вероятно, обусловлен адаптацией животных к ночному и сумеречному образу жизни, обитанию в тенистых лесах и др. В этих условиях увеличивается значение слухового и обонятельного анализаторов, которые начинают играть роль интегративного аппарата в организации целостных поведенческих реакций. Намечается также переход ведущей роли в нервной деятельности от височной к центральной сенсомоторной и затылочной областям.
Третий этап общего направления развития мозга приматов отражает мозг обезьян Нового Света — собственно лемуровых, ревунов, капуциновых, низших обезьян Старого Света (рис. IV. 1, 2). Для этого этапа характерно формирование питекоидного типа мозга с прогрессивным развитием центральных соматотопических сенсомоторных зон и затылочной области. Наблюдается отграничение центральной бороздой прецентральной (агранулярной) двигательной области от теменной сенсорной. Дугообразная борозда отделяет лобную область. Теменная доля четко отделяется от прецентральной,

Рис. IV. 1. Цитоархитектонические карты мозга полуобезьян и низших обезьян. 1 — перохвостая тупайя, 2 — серый гапалемур, 3 — зеленая мартышка
Условные обозначения
Sу. — Сильвиева борозда, с.е. — Центральная борозда, ip. — Межтеменная борозда, lun. — Лунная борозда, рг. — Принципиальная главная борозда, rh. — Ринальная борозда, t.s. — Верхняя височная борозда, arc. — Дугообразная борозда

Рис. IV. 2. Цитоархитектоническая карта мозга обезьян Нового Света и высших человекообразных обезьян. 1 — коата, 2 — капуцин, 3 — орангутан, 4 — гиббон, 5 — шимпанзе, 6 — горилла
Условные обозначения к рис. IV.1, IV2, IV.3, IV.4, IV5, IV.6 Цифры обозначают цнтоархитектонические поля коры по карте Бродмана; 17,18 — поля, связанные с осуществлением зрительных рецепций; 19 — ассоциативное зрительное поле; 4, 6 — двигательные поля; 1-3 — тактильная чувствительность; 41, 42 — слуховые поля; 22 — ассоциативно-слуховое; 43 — вкусовое; 39, 40 — нижнетеменные поля, связанные со стереоскопией и стереогнозией, праксис и манипулирование; 37 — зрительно-гностические функции; 44, 45 — речедвигателъные поля; 8 — глазодвигательное поле; 9, 10, 47 — поля лобной области, связанные с прогнозированием и высшими формами психической деятельности; 23, 24 — лимбические поля, связанные с эмоциями, пищевым, питьевым и половым поведением. Крупно-зернистая штриховка означает наиболее прогрессивно развивающиеся поля неокортекса
Выделяется мозг руконожки мадагаскарской. Она обгоняет других полуобезьян по развитию центральной борозды, которая у нее выражена даже лучше, чем у ряда обезьян. Вероятно, такое значительное развитие сенсомоторной зоны объясняется исключительно тонкой перкуссионной функцией третьего пальца кисти. Руконожки используют этот палец как молоточек, которым постукивают по коре дерева в поисках насекомых, живущих под корой, а также для доставания личинок из коры и древесины.

Своеобразен также мозг обезьян Нового Света. У капуцинов отмечается сходство с антропоморфными обезьянами по развитию филогенетически новой нижнетеменной области, новых подполей затылочной и дифференцировке подполей прецентральной области. С развитием этих областей связывают исключительную способность капуцинообразных к манипулированию предметами, и выраженной конструктивной деятельности. Капуцины отличаются также более сложными общими поведенческими реакциями по сравнению с другими представителями низших обезьян Нового и Старого Света.
У паукообразных обезьян развивается и распространяется в сторону затылочной задняя часть теменной доли. Все переходные извилины Грациоле открыты и заняты теменными формациями. В связи с прогрессивным развитием задней теменной, передней затылочной и височной областей становится разнообразным использование хватательных конечностей и цепкого хвоста. Согласованность мышечно-суставной чувствительности, вестибулярного и зрительного анализаторов достигает наивысшего уровня в специфической ориентации при подвешивании обезьян на ветвях в разных положениях — вверх и вниз головой. В результате у цепкохвостых обезьян тип строения мозга ближе к понгидному типу (тип высших человекообразных обезьян), нежели к типу низших обезьян Старого Света.
Таким образом, на третьем этапе особое значение имеет затылочный и центральный сенсомоторных комплекс полей. Он осуществляет регуляторные взаимодействия между гностическими, кинестетическими и зрительными функциями. Ведущую роль на третьем этапе играет стереоскопия и стереогнозия.
Четвертый этап (см. рис. IV. 2, 3-6) представляет мозг высших человекообразных обезьян — шимпанзе и гориллы. Он имеет следующие особенности: понгидный тип организации мозга; высший, приматный тип структуры борозд; затылочная покрышка уменьшается; теменно-затылочные извилины открыты и заняты теменными формациями; нижнетеменная область увеличена.
У самого примитивного представителя человекообразных обезьян — гиббона — мозг находится в состоянии, как бы переходном к антропоморфному типу. Для него характерно преобразование прецентральной моторной зоны. Нижняя вертикальная ветвь трехлучевой дугообразной борозды переходит в нижнюю прецентральную борозду. Принципальная борозда превращается в нижнюю лобную. Однако состояние нижнетеменной области остается на уровне низших обезьян. Особенности строения мозга соответствуют способам локомоции гиббонов, хотя они могут ходить по земле на двух ногах, основной способ их передвижения — это брахиация: уцепившись длинными руками за ветвь, они сильно раскачиваются, а затем мгновенно перебрасывают тело на большое расстояние на другую ветвь, затем — на следующую и т. д. Манипулирование развито слабо.
Для орангутана характерно дальнейшее развитие прецентральной области, с сильным ветвлением и своеобразием борозд. Вероятно, это связано с развитием брахиаторного типа локомоции. Заметно расширение нижнетеменных полей и зернистый тип цитоархитектоники, что, возможно, коррелирует с древесным образом жизни.
У шимпанзе рельеф борозд лобной области приближается к гоминидному типу. Это соответствует полувыпрямленному положению тела и полуназемному образу жизни. По дифференцировке полей нижнетеменной дольки, стратификации и ширине слоев, а также пирамидизации нейронов мозг шимпанзе приближается к человеческому типу. Эти черты соответствуют высокому уровню психических способностей шимпанзе.
Мозг гориллы характеризуется наибольшим усложнением борозд лобной доли при специфическом, понгидном типе их ветвления. По некоторым признакам, горилла стоит ближе других человекообразных обезьян к человеку.
Пятый этап нашел отражение в мозге ископаемых гоминид — австралопитека, питекантропа,неандертальца (рис. IV. 3). Это — гоминидный тип мозга с прогрессивным развитием филогенетически новых полей: нижнетеменной, верхневисочной и лобной областей. У сильвиевой борозды появляются две ветви — восходящая и горизонтальная, которые формируют комплекс речедвигательного центра. Из наплыва нижнелобной, теменной и височной коры образуется покрышка, погружающая островок. Заметно увеличение и усложнение интрасулькального компонента в составе неокортекса.
Для мозга австралопитека характерно формирование гоминидного типа полушария. Выявляются особые выпячивания в нижнетеменной, а именно в ангулярной (область зрительного гнозиса) и супрамаргинальной (зона мануального праксиса) извилинах. Имеются выпячивания также и в оперкулярной и лобной областях.
У питекантропов появляются выпуклости на эндокранах в нижнетеменной, задневисочной и нижней лобной областях, свидетельствующие о нарастании здесь массы мозга. По мере надвигания участков покрышек лобных, центральных и теменных долей, инсулярные формации оказываются полностью на дне сильвиевой ямы.
У неандертальцев разрастание нижнетеменной, верхней височной и нижнелобной областей усиливается. Это ведет к новому нарастанию покрышек островка и образуется зияющая сильвиева яма. На эндокранах очаги интенсивного роста в нижнетеменной, нижнелобной и височной областях расширяются и сливаются между собой. Рельеф полушария делается более обтекаемым и приближается по своим очертаниям к форме эндокрана у современных людей. Они не имеют больших выпячиваний и западений.

Рис. IV. 3. Топография корковых полей на эндокране австралопитека (А) и синантропа (Б)
Штриховкой обозначены места дефектов, пунктиром — отпечатки сосудов.
R — проекция центральной борозды; со — коронарный шов; L — лямбовидный шов; Ра — возвышение теменное угловое; Fi -возвышение лобное нижнее; М — возвышение прецентральное; Т — возвышение височное; f. Sy — сильвиева яма
Шестой этап — мозг современного человека не имеет себе равных по сложности и многообразию вариантов строения, форме и величине. Специфичность мозга человека проявляется в структурно-количественных соотношениях различных отделов, а также клеточной, волокнистой, нейронной и синаптической организации (рис. IV. 4).
Мозг человека отличается от мозга других приматов по следующим признакам:
1. Максимальное значение весового квадратного указателя (у грызунов — 0,19, у хищных — 1,14, у дельфинов — 6,27, у человекообразных обезьян — 7,35, у слона — 9,82, у человека — 32,0).
2. Максимальное преобладание головного мозга над спинным.
3. Максимальное преобладание новой коры над старой.
4. Резко увеличена площадь филогенетически более молодых отделов: например, площадь нижнетеменной коры в 37 раз выше, чем у обезьян.
5. В лобной доле у человека развиты две передние ветви сильвиевой борозды: восходящая и горизонтальная, разделяющие нижнюю лобную извилину на три: оперкулярную, триангулярную и орбитальную; на двух последних находятся рече-двигательные поля 44 и 45. В целом, размер площади коры и сложность ее микроскопической организации в филогенетически новых формациях достигают своих максимальных значений именно у человека.
6. Особая глубина и сложность ветвлений борозд полушария, появляющаяся после рождения и придающая мозгу индивидуальный рисунок. У человека до 75% коры находится в глубине борозд.

Рис. IV. 4. Цитоархитектоническая карта мозга современного человека (по Бродману) а — дорсолатеральная поверхность; Б — медиальная поверхность
8. Качественные отличия мозга человека от мозга животных, делающие его уникальной материей, имеют в своей основе морфологический субстрат — специфически человеческую морфофункциональную систему. Именно СЧМФС является морфологической основой высшей психической деятельности человека. Она представлена филогенетически новыми полями неокортекса, возникшими в процессе антропогенеза(поля лобной области, теменной, височно-теменно-затылочной подобласти), мощной системой пучков ассоциативных волокон и сосудами бассейна средней мозговой артерии, индуцирующими их развитие и питающими их. СЧМФС включает также специфическую для человека систему членораздельной речи, которая представлена 4 центрами: рече-двигательным, рече-слуховым, рече-зрительным и центром письменной речи.
9. Для коры мозга человека характерна наивысшая степень дифференцированности и сложности колончатой организации, особенно в специфических для него отделах.В процессе эволюции увеличилось количество ассоциативных корковых нейронов, степень ветвлений дендритов и число шипиков на них, число и пластичность синапсов, возросла эффективность медиаторных процессов и ферментных систем мозга.
Все это способствовало образованию новых межнейронных связей и создало предпосылки для совершенствования двигательной и манипуляторной деятельности, интенсификации механизмов памяти и обучения, развития интеллекта, прогнозирования будущих событий и др.
В результате мозг современного человека представляет собой уникальный орган, наивысший продукт эволюции.
Только после того, как последнее дерево будет срублено,
Только после того, как последняя река будет отравлена,
Только после того, как последняя рыба будет поймана,
Только тогда вы поймете, что деньги нельзя есть.
Приматы: эволюция мозга

Считается, что мозг, имеющийся сейчас у приматов, достался им в наследство от их далеких пращуров – насекомоядных предков. Так считает большинство исследователей. Они считают, что эволюция мозга происходила следующим образом: обонятельный отдел мозга уменьшался, появлялась новая кора, которая и оттесняла старую. У приматов в коре переднего мозга поля, где происходит обобщение сигналов от органов чувств. Это так называемые поля вторичного коркового анализа. Рост новой коры у полуобезьян происходил за счет увеличения объема, распрямления поверхности полушарий, а не за счет образования извилин и борозд. Мозг стал крупнее по сравнению с предками, но лишенным борозд.
Такой мозг есть у многих современных полуобезьян, в том числе и ископаемых видов лемуров и долгопятов. Он называется лиссэнцефалическим.
Следующим этапом эволюции мозга стало образование сложного узора из извилин и борозд. Это подтверждается наличием таких узоров у ископаемых и современных полуобезьян, особенно обитающих на территории старого света.
М. Вебер еще в 1936 году отмечал – все то, что характеризует мозг человека, присутствует и в строении мозга обезьяны. В том же году другой исследователь, Ю.Г.Шевченко, обнаружил у высших обезьян структуры, гомологичные полям коры мозга человека. Мозг человека и мозг человекообразной обезьяны практически идентичны по строению. Но, несмотря на это, функциональные возможности мозга обезьян намного ниже человеческого. Размер его также меньше в три раза.
В изучении эволюции мозга у приматов до сих пор остается много белых пятен, хотя некоторые исследователи и придают большое значение объему и площади коры головного мозга.
У человека помимо борозд и извилин первого и второго порядка имеются в наличии и борозды третьего порядка. Как нет двух одинаковых рисунков папиллярных линий на руке человека, так нет и двух одинаковых рисунков извилин и борозд мозга человека.
Читайте также: