
Экстрапирамидная система мозжечок реферат
Обновлено: 05.07.2024
Экстрапирамидная система (лат.: extra — вне, снаружи, в стороне + pyramis, греч.: πϋραμίς — пирамида) – совокупность подкорковых и стволовых образований, моторных путей, которые не проходят через пирамиды продолговатого мозга. Данная система, наряду с кортикальной, участвует в управлении движениями, причем, являясь наиболее филогенетически древней, играет значительную роль в построении и контроле движений, не требующих активации внимания.
Впервые мысль о том, что на состояние двигательных функций оказывает влияние не только образования, входящие в состав пирамидной системы, высказал английский невролог С. Вильсон в 1908г. в процессе изучения заболевания, известного теперь как болезнь Вильсона-Коновалова. С тех пор все структуры головного мозга, которые влияют на состояние поперечнополосатых мышц и участвующие в обеспечении движений стали именовать экстрапирамидными.
По предложению Р. Гранита (Granit R., 1973) структуры пирамидных путей, от которых зависят активные движения тела и его частей, были названы фазическими. Экстрапирамидные структуры, влияющие на двигательные акты, положение, поддержание равновесия тела и его позу - тоническими.
Н.К. Боголепов отмечал, что экстрапирамидная система участвует также в выполнении выразительных, мимических и эмоциональных реакций.
В 1973 г. ведущий американский физиолог П. Милнер (Milner P.) высказал сомнение относительно существования отдельных экстрапирамидной и пирамидной систем. Однако на данный момент, после тщательного изучения функций той и другой системы, их обособленность неоспорима.
Э. система по сравнении с пирамидной является филогенетически более древней (особенно ее паллидарная часть). С развитием пирамидной системы экстрапирамидная система переходит в соподчиненное положение.
Экстрапирамидная система состоит из следующих структур головного мозга:
базальные ганглии (включая полосатое тело), красное ядро, интерстициальное ядро, тектум, чёрная субстанция, ретикулярная формация ствола мозга, ядра вестибулярного комплекса, мозжечок, нижняя олива продолговатого мозга.
Nucleus basales – скопления серого вещества в толще полушарий большого мозга.
Corpus striatum ( полосатое тело ) состоит из n.caudatus и n.lentiformis.
Хвостатое ядро-имеет форму запятой, расположенной в сагиттальной плоскости. Состоит из трех частей: caput, corpus и cauda. Передняя часть головки сращена с белым веществом лобной доли, своими свободными поверхностями (верхней и медиальной) тело хвостатого ядра в височной доле образует дно центральной части бокового желудочка. Хвост направлен в височную долю полушария, где достигает миндалевидного тела. Участвует в организации двигательной активности, формировании условных рефлексов и механизмов памяти.
Чечевицеобразное ядро-залегает латерально от хвостатого ядра и зрительного бугра и состоит из двух частей – латеральной, большей по размеру и медиальной, скорлупы (putamen) и бледного шара (globus pallidus). Функцией чечевицеобразного ядра являются регуляция мышечного тонуса.
Подталамическое ядро (Льюиса) – скопление серого вещества с пронизывающими его волокнами. Прилегает к поверхности внутренней капсулы, отделяющей его от бледного шара, с которым ядро связано большим числом волокон, проходящих в составе субталамического пучка.
Черное вещество- ядро э.с., лежащее в основании ножек мозга, участвует в распределении мышечного тонуса, необходимого для установки тела в определенном положении.
Красное ядро – серое вещество покрышки среднего мозга, является первым интегративным центром управления конечностями.
Промежуточное ядро ( Кахаля ) – содержится в ретикулярной формации среднего мозга, дает начало медиальному продольному пучку, который является важным ассоциативным путем, связующим различные ядра нервов глазных мышц между собой, чем обусловливаются сочетанные движения глаз при отклонении их в ту или другую сторону. Функция его связана также с движениями глаз и головы, возникающими при раздражении аппарата равновесия.
Пластинка четверохолмия – часть среднего мозга, являющаяся рефлекторным центром различного рода движений, возникающих, г.о. под влиянием зрительных и слуховых раздражений.
Нижняя олива- овальное возвышение, находящееся по бокам от каждой пирамиды продолговатого мозга. Отделена от последней переднелатеральной бороздой. Играет важную роль в желудочно-ободочном рефлексе, проводит волокна из медиального спинно-мозжечкового тракта
Cerebellum - часть заднего мозга, отвечающий за регуляцию позы и мышечного тонуса, сенсомоторную координацию позных и целенаправленных движений, координацию быстрых целенаправленных движений.
Часть структур э.с. участвует в формировании ее высшего отдела, так называемой стриопаллидарной системы, состоящей из pallidum (более филогенетически древнего образования, состоящего из бледного шара, ядра Льюиса, красного ядра, черного вещества) и striatum (более молодой части, состоящей из скорлупы и ограды).
Несмотря на объединение стриатума и паллидума в единую систему, в функциональном плане эти структуры отличаются друг от друга. Паллидум представляет собой двигательное ядро, которое оказывает активизирующее воздействие на подкорковые образования. Бледный шар получает афферентные импульсы по волокнам, идущим от стриатума и субталамического ядра. Часть волокон следуют к таламическим ядрам, нейронам красного и покрышечных ядер. Бледный шар, будучи связан эффекторно с центрами среднего и заднего мозга, регулирует и координирует их работу. Одной из функций паллидума считают торможение нижележащих ядер, главным образом красного ядра среднего мозга, поэтому при повреждении бледного шара наблюдается сильное увеличение тонуса скелетной мускулатуры – гипертонус вследствие освобождения красного ядра от тормозящего влияния бледного шара.
Стриатум же оказывает тормозящее действие на подкорковые структуры. Стриатум получает афферентные импульсы главным образом от больших полушарий и таламуса и посылает эфферентные импульсы главным образом к бледному шару. Часть эфферентов не переключаясь идет к черной субстанции. Незначительное число волокон направляется к субталамическому и красному ядру, нижней оливе, голубому пятну, ядрам шва. Полосатое тело рассматривают как эффекторное ядро, не имеющее самостоятельных двигательных функций, но контролирующее функции паллидума. Полосатое тело тормозит деятельность бледного шара, то есть действует на него таким же образом, как он сам действует на красное ядро.
-nucleus basales (corpus striatum, n,amygdaloideum)
Акинетико-ригидный синдром (гипомимия,
Баллизм и гемибаллизм
-substantia grisea, nucleus ruber, n.intersticieles,lamina tecti,
Гипотонически-гиперкинетический синдром (гиперкинезы, атетоз)
-oliva inferior, nucleus vestibularis
Formation reticularis trunci encephali
Функционирование э.с. осуществляется с помощью многочисленных специализированных проводящих путей:
Эфферентные tractus rubrospinalis, tractus tectospinalis, tractus vestibulospinalis , tractus reticulospinalis , tractus olivispinalis.
Рубро-спинальный (путь Монакова) тракт начинается от красного ядра среднего мозга, после выхода этот путь вновь перекрещивается (перекрёст Фореля) расположен в боковых канатиках и оканчивается в V-VII пластинах спинного мозга (промежуточная зона). Прямых связей с мотонейронами нет. Участвует в организации движения всей мускулатуры туловища и конечностей (ходьба, бег), контролирует тонус сгибателей, делая эти движения пластичными, способствуя сохранению определенной позы на протяжении длительного времени.
Вестибуло-спинальные тракты (латеральный и вентральный) активируют моторные системы, связанные с равновесием тела и направлением движения. Они начинаются от крупных вестибулярных ядер продолговатого мозга и моста, оливы продолговатого мозга и оканчивается в VII - IX пластинах спинного мозга (область переднего рога). Латеральный тракт (проходящий в боковом канатике) заканчивается в области шейных отделов, а вентральный (проходящий в переднем канатике) в поясничных отделах. Контролируют и организуют перераспределение мышечного тонуса при любом изменении положения тела и головы в пространстве, контролируют тонус разгибателей.
Ретикуло-спинальные тракты представлены аксонами нейронов ретикулярной формации моста и продолговатого мозга. Они проходят в переднем канатике, образуя два пути: медиальный и латеральный, и заканчиваются в области моторных ядер IX пластины, а также в области VI - VIII пластин. Важное значение этого тракта в механизме поддержания и распределения мышечного тонуса и регуляции поз, участие в диффузном торможении всей мускулатуры необходимо в некоторых поведенческих актах, например, при перегревании организма. Этот путь опосредует активирующее влияние дыхательного и сосудистодвигательного центров.
Текто-спиннальный тракт берет начало в глубоких слоях бугров четверохолмия среднего мозга, проходит в составе вентральных канатиков и оканчивается на интернейронах VII — VIII пластин спинного мозга (область переднего рога). Функционально текто-спинальные пути связаны с координацией движений головы и туловища в ответ на зрительные и звуковые сигналы и обеспечивают осуществления сторожевого рефлекса у человека.
Оливо-спиннальный тракт берет начало от нижнего ядра оливы продолговатого мозга, оно имеет непосредственные связи с корой полушарий лобной доли(корково-оливный путь), с красным ядром(красноядерно-оливный путь) и корой полушарий мозжечка(оливо-мозжечковый путь). Аксоны клеток нижнего ядра собираются в пучок, который прослеживается в передне-медиальном отделе бокового канатика и заканчивается в двигательных ядрах передних рогов спинного мозга. Обеспечивает безусловно-рефлекторное поддержание тонуса мышц шеи, головы и двигательные акты, направленные на сохранение равновесия тела.
Импульсы из экстрапирамидной системы, равно как и из мозжечка и из пирамидной системы, притекают, следовательно, к клеткам переднего рога, где и заканчиваются все только что перечисленные проводники. Окончательный путь к мышце проходит через периферический двигательный нейрон.
Ассоциативные (связи нейронов экстрапирамидной системы между собой)
Экстрапирамидные расстройства можно разделить на два клинических синдрома: акинетико-ригидный и гиперкинетико-гипотонический
Вариантами снижения двигательной активности являются: акинезия - отсутствие движений, брадикинезия - замедленность движений, олигокинезия -бедность движений, гипокинезия - недостаточность двигательной активности.
Ригидность - постоянное пребывание мышц в состоянии тонического напряжения, которое свойственно как мышцамагонистам, так и мышцам-антагонистам, в связи с чем и проявляется пластический характер повышения мышечного тонуса.
При акинетико-ригидном синдроме в начальной стадии его развития мышечная ригидность при болезни Паркинсона обычно асимметрична, может проявляться в какой-либо одной части тела, однако в дальнейшем, по мере прогрессирования заболевания, она становится все более распространенной и со временем генерализованной.
Гипотонически-гиперкинетический синдром развивается при поражении хвостатого ядра и скорлупы (в этих образованиях отмечается избыток медиаторов возбуждения — дофамина и др.). Различают:
атетоз — медленные вычурные тонического характера движения, преимущественно в дистальных отделах конечностей, иногда и в мускулатуре лица (выпячивание губ и др)
хореический гиперкинез (наиболее распространенная форма) — быстрые неритмичные движения в дистальных и проксимальных отделах конечностей, лице, туловище.
Итак, функционально э.с. очень тесно связана с более филогенетически молодой, пирамидной. Но в тоже время ей присущ определенная роль, заключающаяся в 1. Регуляции мышечного тонуса в комплексе с другими структурами.
2. Регуляции темпа, ритма и пластики любого произвольного двигательного акта.
3. Обеспечении двигательного компонента в регуляции безусловных рефлексов (половой, оборонительный, старт-рефлекс и др.).
4. Обеспечении последовательности двигательного акта.
5. Обеспечении моторного компонента эмоциональной сферы.
6. Регуляции высокоспециализированных движений человека, которые достигли уровня автоматизмов
Бурд, Гусев, Коновалов - Неврология и нейрохирургия
Общая неврология А.С. Никифоров, Е.И. Гусев - 2007г. - 720 с
Пневмапсихасоматология человека, Трифонов Е.В. Русско-английская энциклопедия 2013
Нормальная анатомия человека, Гайворонский И.В. 2001г.
Топический диагноз в неврологии, Петтер Дуус
Тут вы можете оставить комментарий к выбранному абзацу или сообщить об ошибке.
Экстрапирамидная система (от экстра. и греч. pyramís — пирамида), совокупность структур мозга, расположенных в больших полушариях и стволе головного мозга и участвующих в центр, управлении движениями, минуя кортикоспинальную, или пирамидную систему.
Экстрапирамидная система состоит из следующих основных структур: хвостатого ядра, скорлупы чечевицеобразного ядра, бледного шара, субталамического ядра, черного вещества и красного ядра. Уровень нижнего порядка этой системы – ретикулярная формация покрышки ствола мозга и спинной мозг.
Содержание
1. Экстрапирамидная система является более древней по сравнению с пирамидной системой. Основные структуры экстрапирамидной системы.
2. Функции экстрапирамидной системы.
3. Экстрапирамидные нарушения.
4. Диагностика патологии.
5. Лечение.
6. Литература.
Прикрепленные файлы: 1 файл
Анатомия ЦНС.doc
1. Экстрапирамидная система является более древней по сравнению с пирамидной системой. Основные структуры экстрапирамидной системы.
2. Функции экстрапирамидной системы.
3. Экстрапирамидные нарушения.
4. Диагностика патологии.
Экстрапирамидная система (от экстра. и греч. pyramís — пирамида), совокупность структур мозга, расположенных в больших полушариях и стволе головного мозга и участвующих в центр, управлении движениями, минуя кортикоспинальную, или пирамидную систему.
1. Экстрапирамидная система является более древней по сравнению с пирамидной системой. Основные структуры экстрапирамидной системы.
Частью этой системы также являются те пучки, которые связывают кору большого мозга с экстрапирамидными серыми структурами: полосатым телом, красным ядром, черным веществом, мозжечком, ретикулярной формацией и ядрами покрышки ствола.
В этих структурах импульсы передаются на вставочные нервные клетки и затем спускаются как покрышечно-, красноядерно-спинномозговые, ретикулярно– и преддверно-спинномозговые и другие пути к мотонейронам передних рогов спинного мозга. Через эти пути экстрапирамидная система влияет на спинномозговую двигательную активность.
Пирамидный путь (через вставочные нейроны) и волокна экстрапирамидной системы в конечном итоге встречаются на мотонейронах переднего рога, на альфа– и гамма-клетках и влияют на них путем как активации, так и торможения.
Экстрапирамидная система является филогенетически более древней (особенно ее паллидарная часть) по сравнению с пирамидной системой. С развитием пирамидной системы экстрапирамидная система переходит в соподчиненное положение.
Экстрапирамидная система состоит из следующих основных структур: хвостатого ядра, скорлупы чечевицеобразного ядра, бледного шара, субталамического ядра, черного вещества и красного ядра. Уровень нижнего порядка этой системы – ретикулярная формация покрышки ствола мозга и спинной мозг.
С дальнейшим развитием животного мира палеостриатум (бледный шар) стал главенствовать над этими структурами. Затем у высших млекопитающих ведущую роль приобретает неостриатум (хвостатое ядро и скорлупа). Как правило, филогенетически более поздние центры доминируют над более ранними. Это означает, что у низших животных обеспечение иннервации движений принадлежит экстрапирамидной системе.
Полосатое тело – ведущий центр среди структур, составляющих экстрапирамидную систему. Он получает импульсы от различных областей коры большого мозга, особенно от лобной двигательной области коры, включающей поля 4 и 6. Эти афферентные волокна организованы в соматотопической проекции, идут ипсилатерально и являются ингибиторными (тормозящими) по своему действию.
Достигает полосатого тела и другая система афферентных волокон, идущих от таламуса. От хвостатого ядра и скорлупы чечевицеобразного ядра основные афферентные волокна направляются к латеральному и медиальному сегментам бледного шара, которые отделены друг от друга внутренней медуллярной пластинкой. Существуют связи, идущие от ипсилатеральной коры большого мозга к черному веществу, красному ядру, субталамическому ядру, ретикулярной формации.
Все другие эфферентные волокна полосатого тела проходят через медиальный сегмент бледного шара. Они образуют довольно толстые пучки волокон. Один из этих пучков называется лентикулярной петлей. Ее волокна начинаются в вентральной части медиального сегмента бледного ядра и идут вентромедиально вокруг задней ножки внутренней капсулы к таламусу и гипоталамусу, а также реципрокно к субталамическому ядру.
После перекреста они соединяются с ретикулярной формацией среднего мозга, от которой цепь нейронов формирует ретикулярно-спинномозговой путь (нисходящая ретикулярная система), заканчивающийся в клетках передних рогов спинного мозга.
Основная часть эфферентных волокон бледного шара идет к таламусу. Это паллидоталамический пучок, или поле Фореля Н1. Большинство его волокон заканчивается в передних ядрах таламуса, которые проецируются на кортикальное поле 6. Волокна, начинающиеся в зубчатом ядре мозжечка, заканчиваются в заднем ядре таламуса, которое проецируется на кортикальное поле 4.
Все эти таламокортикальные соединения передают импульсы в обоих направлениях. В коре таламокортикальные пути образуют синапсы с кортикостриарными нейронами и формируют кольца обратной связи. Реципрокные (сопряженные) таламокортикальные соединения облегчают или ингибируют активность кортикальных двигательных полей.
Волокна базальных ядер, которые спускаются к спинному мозгу, сравнительно немногочисленны и достигают спинного мозга только через цепь нейронов. Этот характер соединений позволяет предположить, что основная функция базальных ядер – контроль и регулирование активности моторных и премоторных кортикальных полей, поэтому произвольные движения могут быть выполнены плавно, непрерывно.
Пирамидный путь начинается в сенсомоторной области коры большого мозга (поля 4, 1,2, 3). Это в то же время поля, в которых начинаются экстрапирамидные двигательные пути, которые включают кортикостриарные, кортикорубральные, кортиконигральные и кортикоретикулярные волокна, идущие к двигательным ядрам черепных нервов и к спинномозговым двигательным нервным клеткам через нисходящие цепи нейронов.
Большинство этих связей коры проходит через внутреннюю капсулу. Следовательно, повреждение внутренней капсулы прерывает не только волокна пирамидного пути, но и экстрапирамидные волокна. Этот перерыв является причиной мышечной спастичности.
2. Функции экстрапирамидной системы.
Основными физиологическими функциями экстрапирамидной системы являются координация движений, регуляция мышечного тонуса и поддержание позы, организация двигательных проявлений эмоций. Из-за сложного строения экстрапирамидной системы, обширных связей ее структур с различными образованиями головного мозга трудно понять физиологические механизмы экстрапирамидной регуляции движений. Экстрапирамидная система не разделяется на отдельные пути, она является сложной системой двигательных ядер и связей между ними, а также путей двигательных центров головного мозга с эфферентными нейронами спинного мозга и ядрами черепно-мозговых нервов через многочисленные подкорковые и стволовые структуры. В спинном мозге импульсы, проходящие по нисходящим пирамидному тракту и волокнам экстрапирамидной системы, взаимодействуют с возбуждениями, передающимися по афферентным путям от про-приоцепторов. Процесс передачи возбуждений на уровне спинного мозга играет важную роль в механизме произвольных и непроизвольных движений.
Началом экстрапирамидной системы являются аксоны нейронов коркового двигательного поля 4 и нейроны, находящиеся в соматосенсорной коре. Наряду с тем многие волокна экстрапирамидной системы берут свое начало и в других сенсорных областях коры (слуховой, зрительной, вкусовой) и в ассоциативных зонах лобной, теменной и височной долей мозга. От клеток коры головного мозга импульсы по экстрапирамидным путям стремятся к нейронам таламуса, гипоталамуса, моста головного мозга, красных ядер, черного вещества и ретикулярной формации. От поля 4 экстрапирамидные волокна вместе с волокнами пирамидного тракта направляются в составе внутренней капсулы к наиболее древним группам двигательных ядер – базальным ядрам. Эти ядра – главные среди структур экстрапирамидной системы, так как являются высшим надсегментарным аппаратом, выполняющим регуляцию двигательных актов с участием различных мышечных групп. Благодаря базальным ядрам выполняются все синергии, входящие в состав таких сложных движений, как бег, плавание, ходьба и другое. При участии экстрапирамидной системы обеспечивается плавность движений и устанавливается исходная поза, необходимая для их выполнения. Экспериментальные исследования и клинические наблюдения отмечают многообразие форм и в то же время однозначность двигательных реакций при разрушении или раздражении экстрапирамидных образований. Повреждения структур экстрапирамидной системы вызывают различные двигательные нарушения. При повреждении внутренней капсулы в глубине полушария головного мозга, где проходят пирамидные и экстрапирамидные волокна, проявляется спастическое повышение тонуса мышц парализованных конечностей, которое вызвано разрушением именно экстрапирамидных проводников, обеспечивающих в норме тормозное влияние на ретикулярную формацию. Корковое торможение бывает двух видов – торможение мышечного тонуса и торможение совершающегося движения. Последний вид торможения свойственен для нисходящих корковых экстрапирамидных влияний. Они выполняют основную роль в регуляции физиологической активности двигательных экстрапирамидных ядер мозгового ствола, от которых к двигательным центрам спинного мозга идет мощный поток возбуждений. Экстрапирамидные движения, вызываемые вследствие стимуляции различных участков коры, более медленны и стереотипны. Если изолировать тормозные влияния коры головного мозга, то движения станут судорожными мышечными сокращениями.
Функции экстрапирамидной системы. Стриатум экстрапирамидной системы считается высшим подкорковым регуляторно-координационным центром организации движений, а каллидум, действуя на нейроны спинного мозга через структуры среднего и продолговатого мозга, координирует тонус и фазовую двигательную активность мышц. Деятельность стриопаллидарных образований связана с выполнением медленных сложных движений, например, медленная ходьба, вдевание нитки в иголку и т.д.
Пирамидная и экстрапирамидная системы успевают в момент выполнения движения внести исправления в непрерывный поток двигательных возбуждений. Если разрушить стриатум, то образуются непроизвольные хаотические движения отдельных конечностей – гиперкинезы. При нарушениях пирамидной и экстрапирамидной регуляции произвольной и непроизвольной двигательной активности мимических мышц возникает неадекватное внешнее выражение эмоций, непроизвольный смех и плач или полное отсутствие мимического выражения (маскообразное лицо).
Одна из функций бледного шара характеризуется торможением нижележащих ядер среднего мозга. После повреждения бледного шара проявляется увеличение тонуса скелетной мускулатуры в результате освобождения красного ядра среднего мозга от тормозящего влияния паллидума. Раздражение бледного шара ведет к повышению тонуса мышц и тремору конечностей, а также к ограничению и скованности движений. Эти явления исчезают при разрушении паллидума. При раздражении полосатого тела происходит торможение движений. Такие эффекты подавления движений наблюдаются при раздражении тормозных зон экстрапирамидной системы (поясная кора головного мозга, части моторной коры, хвостатое ядро, мозжечок, ретикулярная формация).
В стриатуме берут начало многие двигательные пути экстрапирамидной системы, среди которых выделяют эффекторный путь, ведущий к паллидуму, а затем через красное ядро и рубро-спинальный тракт – к спинному мозгу. Стриатум довольно хорошо морфологически и нейрофизиологически изучен. Множество внутренних связей считаются особенностью его строения, однако небольшое число нейронов направляют свои аксоны к ядрам таламуса и гипоталамуса.
Экспериментальные физиологические исследования с помощью микроэлектродов доказали, что стриатум вдвойне влияет на нейроны бледного шара – возбуждающе и тормозя. Предполагают, что морфологическим субстратом подобных тормозных воздействий оказываются прямые восходящие пути перечисленных структур с моторной и соматической корой, а также таламическими ядрами и срединным центром.
Головка хвостатого ядра является важной в организации предпусковых процессов, которые включают перестройку позы, предшествующей произвольному двигательному акту.
Большинство образований экстрапирамидной системы не образует прямых выходов к мотонейронам спинного мозга, их влияние на мотонейроны опосредованно через ретикулоспинальный тракт, который является как бы общим конечным путем экстрапирамидной системы.
Экстрапирамидные воздействия на спинной мозг и ядра черепно-мозговых нервов выполняются через трункоспинальные пути, проводящие импульсы от ядер промежуточного мозга, среднего мозга и продолговатого мозга. К ним относят руброспинальный, вестибулоспинальный и оливоспинальный пути. Через эти же пути воздействует часть мозжечковых влияний на мотонейроны спинного мозга. Общность эффектов рассматриваемых воздействий с экстрапирамидной регуляцией разрешает отнести эти структуры к экстрапирамидной системе.
Все супраспинальные структуры, входящие в состав экстрапирамидной системы, направляют свои действия к у-мотонейронам спинного мозга. Гамма-мотонейроны регулируют поток проприоцептивных афферентных импульсов, идущих в спинной мозг от мышечных веретен. Эти афферентные импульсы воздействуют на возбудимость а-мотонейронов, активность которых характеризует рабочее состояние мышц. В этом процессе существует прямое управление мотонейронами с помощью сигналов, идущих от первичных проприорецепторов. Нисходящие воздействия со стороны образований экстрапирамидной системы облегчают или подавляют рефлекс растяжения, что наблюдается при децеребрационной ригидности, когда крайне усиливаются миостатические рефлексы. Усиливающим фактором считается увеличение под действием нисходящих воздействий активности у-мотонейронов, которая одновременно ведет к возрастанию разрядов рецепторов растяжения и соответствующей интенсификации моносиноптического возбуждения а-мотонейронов. Быстропроводящие волокна, начинающиеся из медиальной части ретикулярной формации продолговатого мозга и варониева моста, а также из материального вестибулярного ядра Дейтерса, тоже моносинаптически возбуждают а-мотонейроны спинного мозга и обеспечивают быстрое движение. Медленнопроводящий нисходящий путь покрышки среднего мозга осуществляет регуляцию тонических реакций. Таким образом, если нисходящее воздействие пирамидной системы, воздействуя непосредственно на а-мотонейроны, увеличивает их функциональную активность при осуществлении фазных и тонических двигательных реакций организма, то регулирующее воздействие экстрапирамидной системы на у-мотонейроны обуславливает нужную коррекцию выполняемых движений и является дополнительным механизмом влияния на познотоническую и двигательную активность. Тормозные регулирующие воздействия, ведущие от двигательных центров коры и стриопаллидарных структур, корректирует степень напряжения скелетных мышц.
Экстрапирамидная система (лат.: extra — вне, снаружи, в стороне + pyramis, греч.: πϋραμίς
— пирамида) – совокупность подкорковых и стволовых образований, моторных путей, которые не проходят через пирамиды продолговатого мозга. Данная система, наряду с кортикальной, участвует в управлении движениями, причем, являясь наиболее филогенетически древней, играет значительную роль в построении и контроле движений, не требующих активации внимания.
История изучения.
Впервые мысль о том, что на состояние двигательных функций оказывает влияние не только образования, входящие в состав пирамидной системы, высказал английский невролог С. Вильсон в 1908г. в процессе изучения заболевания, известного теперь как болезнь Вильсона-Коновалова. С тех пор все структуры головного мозга, которые влияют на состояние поперечнополосатых мышц и участвующие в обеспечении движений стали именовать экстрапирамидными.
По предложению Р. Гранита (Granit R., 1973) структуры пирамидных путей, от которых зависят активные движения тела и его частей, были названы фазическими.
Экстрапирамидные структуры, влияющие на двигательные акты, положение, поддержание равновесия тела и его позу - тоническими.
Н.К. Боголепов отмечал, что экстрапирамидная система участвует также в выполнении выразительных, мимических и эмоциональных реакций.
В 1973 г. ведущий американский физиолог П. Милнер (Milner P.) высказал сомнение относительно существования отдельных экстрапирамидной и пирамидной систем. Однако на данный момент, после тщательного изучения функций той и другой системы, их обособленность неоспорима.
Описание
Экстрапирамидная система по сравнении с пирамидной является филогенетически более древней (особенно ее паллидарная часть). С развитием пирамидной системы экстрапирамидная система переходит в соподчиненное положение.
Экстрапирамидная система состоит из следующих структур головного мозга: базальные ганглии (включая полосатое тело), красное ядро, интерстициальное ядро, тектум, чёрная субстанция, ретикулярная формация ствола мозга, ядра вестибулярного комплекса, мозжечок, нижняя олива продолговатого мозга.
Nucleus basales – скопления серого вещества в толще полушарий большого мозга.
Corpus striatum ( полосатое тело ) состоит из n.caudatus и n.lentiformis.
Хвостатое ядро-имеет форму запятой, расположенной в сагиттальной плоскости.
Состоит из трех частей: caput, corpus и cauda. Передняя часть головки сращена с белым веществом лобной доли, своими свободными поверхностями (верхней и медиальной) тело хвостатого ядра в височной доле образует дно центральной части бокового желудочка.
Хвост направлен в височную долю полушария, где достигает миндалевидного тела.
Участвует в организации двигательной активности, формировании условных рефлексов и механизмов памяти.
Чечевицеобразное ядро-залегает латерально от хвостатого ядра и зрительного бугра и состоит из двух частей – латеральной, большей по размеру и медиальной, скорлупы
(putamen) и бледного шара (globus pallidus). Функцией чечевицеобразного ядра являются регуляция мышечного тонуса.
Подталамическое ядро (Льюиса) – скопление серого вещества с пронизывающими его волокнами. Прилегает к поверхности внутренней капсулы, отделяющей его от бледного шара, с которым ядро связано большим числом волокон, проходящих в составе субталамического пучка.
Черное вещество - ядро э.с., лежащее в основании ножек мозга, участвует в распределении мышечного тонуса, необходимого для установки тела в определенном положении.
Красное ядро – серое вещество покрышки среднего мозга, является первым интегративным центром управления конечностями.
Промежуточное ядро ( Кахаля ) – содержится в ретикулярной формации среднего мозга, дает начало медиальному продольному пучку, который является важным ассоциативным путем, связующим различные ядра нервов глазных мышц между собой, чем обусловливаются сочетанные движения глаз при отклонении их в ту или другую сторону.
Функция его связана также с движениями глаз и головы, возникающими при раздражении аппарата равновесия.
Пластинка четверохолмия – часть среднего мозга, являющаяся рефлекторным центром различного рода движений, возникающих, г.о. под влиянием зрительных и слуховых раздражений.
Нижняя олива- овальное возвышение, находящееся по бокам от каждой пирамиды продолговатого мозга. Отделена от последней переднелатеральной бороздой. Играет важную роль в желудочно-ободочном рефлексе, проводит волокна из медиального спинно-мозжечкового тракта
Cerebellum - часть заднего мозга, отвечающий за регуляцию позы и мышечного тонуса, сенсомоторную координацию позных и целенаправленных движений, координацию быстрых целенаправленных движений.
Часть структур э.с. участвует в формировании ее высшего отдела, так называемой стриопаллидарной системы, состоящей из pallidum (более филогенетически древнего образования, состоящего из бледного шара, ядра Льюиса, красного ядра, черного вещества) и striatum (более молодой части, состоящей из скорлупы и ограды).
Несмотря на объединение стриатума и паллидума в единую систему, в функциональном плане эти структуры отличаются друг от друга. Паллидум представляет собой двигательное ядро, которое оказывает активизирующее воздействие на подкорковые образования. Бледный шар получает афферентные импульсы по волокнам, идущим от стриатума и субталамического ядра. Часть волокон следуют к таламическим ядрам, нейронам красного и покрышечных ядер. Бледный шар, будучи связан эффекторно с центрами среднего и заднего мозга, регулирует и координирует их работу. Одной из
функций паллидума считают торможение нижележащих ядер, главным образом красного ядра среднего мозга, поэтому при повреждении бледного шара наблюдается сильное увеличение тонуса скелетной мускулатуры – гипертонус вследствие освобождения красного ядра от тормозящего влияния бледного шара.
Стриатум же оказывает тормозящее действие на подкорковые структуры. Стриатум получает афферентные импульсы главным образом от больших полушарий и таламуса и посылает эфферентные импульсы главным образом к бледному шару. Часть эфферентов не переключаясь идет к черной субстанции. Незначительное число волокон направляется к субталамическому и красному ядру, нижней оливе, голубому пятну, ядрам шва.
Полосатое тело рассматривают как эффекторное ядро, не имеющее самостоятельных двигательных функций, но контролирующее функции паллидума. Полосатое тело тормозит деятельность бледного шара, то есть действует на него таким же образом, как он сам действует на красное ядро.
Классификация
Topography
Рас-ва
Functio
Рас-ва
Telencephalon
(substantia grisea)
-nucleus basales (corpus striatum, n,amygdaloideum)
Хорея
Activatio- pallidum
Акинетико-ригидный синдром (гипомимия, брадикинезия)
Diencephalon
- nucleus subthalamicus
Баллизм и гемибаллизм
Mesencephalon
-substantia grisea, nucleus ruber, n.intersticieles,lamina tecti,
Паркинсонизм Reactivatio- striatum
Гипотонически- гиперкинетический синдром (гиперкинезы, атетоз)
Medulla oblongata
-oliva inferior, nucleus vestibularis
Cerebellum
Formation reticularis trunci encephali
Функционирование экстрапирамидной системы осуществляется с помощью
многочисленных специализированных проводящих путей:
Афферентные корковые пути берут свое начало от многочисленных отделов коры больших полушарий, особенно из двигательных зон лобной области (предцентральная извилина, парацентральная долька). Вероятнее всего данные волокна являются тормозными. Другая система афферентных волокон, которая, по-видимому является активирующей, достигает полосатого тела из таламического центромедианного ядра.
Кроме этого выделяется система корковых волокон, обеспечивающих двусторонние связи между черной субстанцией и стриатумом. Дофаминэргические (афферентные нигростициальные) волокна – уменьшают тормозную функцию стриатума (аксонами нейронов этих путей выделяется около 80% дофамина, (нигростриарная система,
Импульсы из экстрапирамидной системы, равно как и из мозжечка и из пирамидной системы, притекают, следовательно, к клеткам переднего рога, где и заканчиваются все только что перечисленные проводники. Окончательный путь к мышце проходит через периферический двигательный нейрон.
Ассоциативные (связи нейронов экстрапирамидной системы между собой)
Экстрапирамидные расстройства можно разделить на два клинических синдрома: акинетико-ригидный и гиперкинетико-гипотонический
Вариантами снижения двигательной активности являются: акинезия - отсутствие движений, брадикинезия - замедленность движений, олигокинезия -бедность движений, гипокинезия - недостаточность двигательной активности.
Ригидность - постоянное пребывание мышц в состоянии тонического напряжения, которое свойственно как мышцамагонистам, так и мышцам-антагонистам, в связи с чем и проявляется пластический характер повышения мышечного тонуса.
При акинетико-ригидном синдроме в начальной стадии его развития мышечная ригидность при болезни Паркинсона обычно асимметрична, может проявляться в какой- либо одной части тела, однако в дальнейшем, по мере прогрессирования заболевания, она становится все более распространенной и со временем генерализованной.
Гипотонически-гиперкинетический синдром развивается при поражении хвостатого ядра и скорлупы (в этих образованиях отмечается избыток медиаторов возбуждения — дофамина и др.). Различают:
• атетоз — медленные вычурные тонического характера движения, преимущественно в дистальных отделах конечностей, иногда и в мускулатуре лица
(выпячивание губ и др)
• хореический гиперкинез (наиболее распространенная форма) — быстрые неритмичные движения в дистальных и проксимальных отделах конечностей, лице, туловище.
Итак, функционально эктрапирамидная система очень тесно связана с более
филогенетически молодой, пирамидной. Но в тоже время ей присущ определенная роль,
заключающаяся в
1. Регуляции мышечного тонуса в комплексе с другими структурами.
2. Регуляции темпа, ритма и пластики любого произвольного двигательного акта.
3. Обеспечении двигательного компонента в регуляции безусловных рефлексов (половой, оборонительный, старт-рефлекс и др.).
4. Обеспечении последовательности двигательного акта.
5. Обеспечении моторного компонента эмоциональной сферы.
6. Регуляции высокоспециализированных движений человека, которые достигли уровня автоматизмов
короткие, молниеносные клонические подергивания мышц и групп мышечных волокон
(чаще в проксимальных отделах конечностей, не вызывая движения конечности)
Тики - быстрые клонические подергивания ограниченной группы мышц стереотипного характера, имитирующие произвольные движения
Тремор - стереотипный ритмический клонический гиперкинез. Различают крупноразмашистый (рубральный) тремор, интенционный тремор (возникающий при выполнении целенаправленных движений), статический тремор (тремор покоя, уменьшающийся при выполнении произвольных движений)
Торсионная дистония - судорожные штопорообразные переразгибания позвоночника в поясничном и шейном отделе с формированием вычурных поз, проявляющиеся при произвольных движениях, сколиоз, гиперлордоз. В начальной стадии проявлением торсионной дистонии может быть спастическая кривошея - судорожные сокращения мышц шеи с поворотом головы в сторону, к плечу, откидыванием головы назад и пожиманием плечами.
Мозжечок
Располагается в области задней черепной ямки, позади моста, под затылочными долями.
Строение:
В области белого вещества располагается серое вещество, образуя ядра:
- более древние – ядро шатра, шаровидное и пробковидное ядро – участвуют в осуществлении движения туловища
- более молодое – зубчатое ядро
Серое вещество располагается и на поверхности – кора трехслойная
- зернистый слой – глубокий, содержит большое количество мелких клеток, короткие дендриты, длинные аксоны, грушевидные клетки Гольджи
- ганглиозный – промежуточный слой, содержит клетки Пуркинье, дендриты которых оканчиваются в молекулярном слое.
- молекулярный – содержит корзинчатые летки, которые контактируют с клетками
Пуркинье.
Мозжечок имеет связи с другими отделами нервной системы за счет трех пар ножек:
- верхние ножки содержат афферентные пути - передний спиномозжечковый (от задних рогов спинного мозга до зубчатого ядра) – и эфферентные – верхний спиномозжечковый(связь с вестибулярным центром и красным ядром)
- средние ножки содержат многочисленные волокна от собственных ядер моста
- нижние ножки содержат пути: задний спиномозжечковый(Флексига), от проводников глубокой чувствительности, от ядер вестибулярного аппарата, от ядер олив, от экстрапирамидной системы.
Два основных спиномозжечковых пути – пути Флексига и Говерса.
Путь Говерса – предний спиномозжечковый. Первый нейрон пути находится в спинномозговом ганглии. Его дендриты идут к рецепторам на связках, сухожилиях, надкостнице. Его аксон идет ко второму нейрону, который располагается в задниз рогах спинного мозга, формируя столбы Кларка. Волокна этих нейронов переходят на противоположную сторону и идут вверх в боковых канатиках кпереди от заднего пучка, образуя передний спиномозжечковый путь. Путь продолжается до ствола мозга, проходит через продолговатый мозг, мост, на уровне переднего мозгового паруса переходит на противоположную сторону и в составе верхней ножки мозжечка достигает ядер мозжечка(3ий нейрон в черве, 4ый – грушевидные клетки, 5ый – зубчатое ядро).
Путь Флексига – задний спиномозжечковый. Первый нейрон пути находится в спинномозговом ганглии. Его дендриты идут к рецепторам мышц, связок, сухожилий, надкостницы. Его аксон идет ко второму нейрону – колонке Кларка. Волокна вторых нейронов идут вверх по своей стороне, на уровне продолговатого мозга проходят в нижних ножках и оканчиваются в клетках коры червя. Там заложен третий нейрон, аксон которого идут к грушевидным клеткам (4-ый нейрон), которые связаны с зубчатым ядром
(5ый нейрон).
Основной путь афферентных импульсов от коры головного мозга – tractusfrontocerebellaris. Первые нейроны находятся в области верхней лобной извилины, их аксоны идут через лучистый венец, далее через переднюю ножку внутренней капсулы и в области моста достигают вторых нейронов – собственных ядер моста. Далее образуют перекрест, проходят в средних ножках и оканчиваются в коре полушарий мозжечка.
Другой путь – лобно-мосто-мозжечковый. Превые нейроны располагаются диффузно в переднем отделе верхней и средней лобных извилин. Аксоны первых нейронов проходят через лучистый венец, далее в передней ножке внутренней капсулы, проходят в основании ножки мозга и достигают вторых нейронов – ядер моста. Аксоны вторых нейронов образуют перекрест и через кору мозжечка достигают зубчатого ядра.
Аксоны зубчатого ядра проходят через верхние ножки, где делятся на восходящие (пути к покрышке мозга, к вестибулярным ядрам, к крое больших полушарий и таламусу) и нисходящие – к красному ядру противоположной стороны (перекрест Вернекинка). От красного ядра начинается эфферентный пучок Монакова. Аксоны совершают в покрышке вентральный подъядерный перекрест Фореля.
Мозжечок оказывает влияние на малые альфа-мотонейроны и гамма-мотонейроны передних рогов спинного мозга через вестибулярные ядра и ретикулярную фармацию, тем самым регулируя мышечный тонус.
Симптомы нарушения функции мозжечка
Мозжечковая атаксия – нарушение координации движений, в связи с чем они становятся неловкими и несоразмерными. При этом наблюдается расстройство целенаправленных движений – ходьбы (атаксическая походка). Больной при ходьбе широко расставляет ноги, идет зигзагообразно, что напоминает походку пьяного человека, часто падает; ему трудно стоять, особенно на одной ноге. При попытке взять какой-нибудь предмет больной промахивается, протягивая руку слишком далеко или не доводя ее до предмета.
При закрытых глазах моторика существенно не ухудшается. Одновременно может наблюдаться с начало мелкое, а при приближении руки к предмету все более крупное дрожание (интенционное дрожание). Речь расстраивается, теряет плавность. Ударения расставляются больным не по смыслу, а разделены равномерными интервалами
(скандированная речь).
Сенситивная атаксия обусловлена нарушением глубокой чувствительности с выпадением афферентных импульсов, направляющихся по задним канатикам спинного мозга. Под контролем зрения движения улучшаются. При закрытых глазах сенситивная атаксия усиливается.
При лабиринтной атаксии расстройства координации сочетаются с вестибулярными симптомами – головокружением, горизонтальным нистагмом, отклонением при ходьбе в сторону пораженного лабиринта. Дифференциальную диагностику облегчают специальные исследования.
Корковая атаксия возникает при поражении верхних отделов лобной доли и нижних отделов затылочной и височных долей, а также корково-мозжечковых путей, соединяющих эти отделы КГМ с корой мозжечка. В отличие от мозжечковой, сенситивной и лабиринтной атаксии, нарушение координации движений наблюдаются на стороне противоположной стороне поражения.
Атаксия может быть ведущим симптомом заболевания. Семейная атаксия Фридрейха развивается в детском возрасте; основными симптомами являются расстройства координации движений – мозжечковая атаксия с элементами сенситивной атаксии, а также отсутствие сухожильных рефлексов. Наследственная мозжечковая атаксия развивается позднее, чем атаксия Фридрейха. Отмечаются пирамидные симптомы и прогрессирующее снижение интеллекта в сочетании с ведущим признаком болезни – мозжечковой атаксией.
При инфекционных болезнях (пневмония, брюшной и сыпной тифы, дифтерия и др.) может развиваться острая атаксия. Нарушение координации движений в этих случаях появляются на высоте инфекционных процессов.
Симптомы атаксии могут возникать также при различных интоксикациях (алкоголь, пищевые отравления, ртуть и др.), а также при нарушениях мозгового кровообращения и при опухолях мозга.
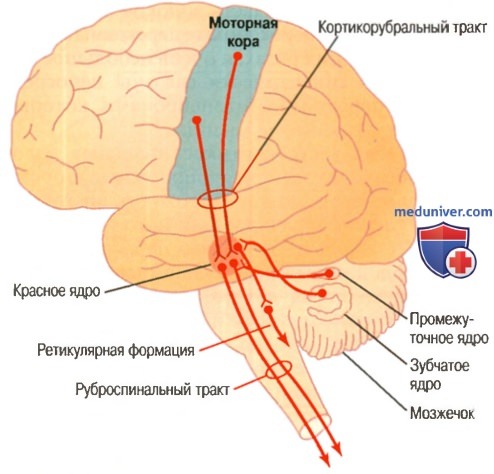
Кортикоруброспинальный путь двигательного контроля; показана также связь этого пути с мозжечком
а) Возбуждение двигательных областей спинного мозга первичной моторной корой и красным ядром. Нейроны моторной коры организованы в вертикальные колонки. В отдельных статьях на сайте (просим вас пользоваться формой поиска выше) указывалось, что клетки соматосенсорной и зрительной коры организованы в вертикальные колонки. Клетки моторной коры также собраны в вертикальные колонки, диаметр которых составляет долю миллиметра; одна колонка включает тысячи нейронов.
Каждая колонка клеток функционирует как единое целое, обычно стимулируя группу мышц-синергистов, а иногда лишь одну мышцу. Кроме того, как и вся кора большого мозга, колонка имеет 6 отдельных слоев клеток. Все пирамидные клетки, дающие начало кортикоспинальным волокнам, лежат в 5 слое клеток от поверхности коры, а сигналы входят в колонку через 2-4 слои; 6 слой дает начало основной части волокон, которые связывают колонку с другими регионами самой коры большого мозга.
Это важно, поскольку стимуляция одиночной пирамидной клетки редко может возбудить мышцу. Обычно для вызова сокращения определенной мышцы нужно, чтобы одновременно или в быстрой последовательности возбудились 50-100 пирамидных клеток.
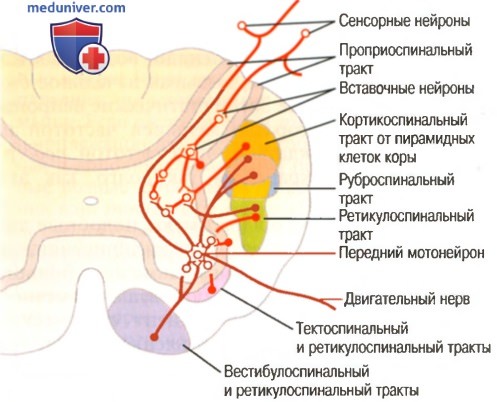
Конвергенция различных двигательных регуляторных путей на передних мотонейронах
в) Динамические и статические сигналы, передаваемые пирамидными нейронами. Если для запуска быстрого сокращения к мышце посылается сильный сигнал, то дальнейшее длительное поддержание сокращения может обеспечить гораздо более слабый продолжительный сигнал. Это обычный характер возбуждения, обеспечивающий мышечные сокращения.
Для этого каждая колонка клеток возбуждает две популяции пирамидных нейронов, одну из которых называют динамическими нейронами, а другую — статическими нейронами. В течение короткого периода в начале сокращения интенсивно возбуждаются динамические нейроны, вызывая начальное быстрое разбитие силы. Затем статические нейроны возбуждаются с гораздо меньшей частотой и, продолжая возбуждаться с этой частотой, поддерживают силу сокращения так долго, как это необходимо.
Нейроны красного ядра имеют подобные динамические и статические характеристики, за исключением того, что в красном ядре больше процент динамических нейронов, а в первичной моторной коре больше процент статических нейронов. Возможно, это объясняется тем, что красное ядро тесно связано с мозжечком, а мозжечок играет важную роль в быстрой инициации мышечного сокращения.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Читайте также: