
Биологическое значение чередования поколений растений и животных реферат
Обновлено: 02.07.2024
Чередование поколений – это тип жизненного цикла у наземных растений, а некоторые водоросли в котором последующие поколения людей чередуются между гаплоидный а также диплоид организмы. Это можно противопоставить половое размножение у животных, у которых в каждом поколении обнаружены как гаплоидные, так и диплоидные клетки. Смена поколений имеет несколько отличительных черт, и эти черты могут быть слегка изменены между вид, Как правило, поколения чередуются между спорофитами, способными создавать споры, и гаметофитами, способными создавать гамет.
Чередование жизненного цикла поколений
Спорофит
Затем, когда спорофит достигает зрелости, наступает ключевой момент смены поколений. У спорофита развиваются органы, известные как спорангия. Эти специализированные репродуктивные органы используются для создания одноклеточных гаплоидных спор. Эти клетки будут выпущены в воздух или воду и унесены. Когда они достигнут подходящей среды, они начнут процесс превращения в гаметофит.
гаметофита
Это представляет следующее поколение в чередовании поколений, поскольку гаплоидная спора создана. Спора технически новый организм, и имеет только половину ДНК в качестве родительского организма. Эта спора подвергнется последовательным митозам, чтобы сформировать нового многоклеточного индивидуума – гаметофит. Там, где поколение спорофитов создает споры, поколение гаметофитов создает гаметы. Гамет вырабатываются специальными органами на гаметофите – гаметангии. Эти гаметы затем передаются в окружающую среду или передаются между растениями.
Когда они находят противоположность гамета, они начинают процесс слияния, чтобы сформировать другую зиготу. Эта зигота в конечном итоге станет спорофитом, и чередование поколений продолжит вращаться. Хотя это упрощенная версия смены поколений, существует много сложностей, которые будут обсуждаться ниже. Из-за этих сложностей и из-за того, что все растения подвергаются некоторой версии смены поколений, ученые предпочитают ссылаться на другие аспекты своих репродуктивных циклов для определения вида.
Самая простая форма смены поколений находится в папоротнике, как показано ниже. Как видно из диаграммы, гаметофит и спорофит – это явно разные люди. Это не всегда так.

Осложнения с чередованием поколений
Половые гаметы
Есть много дополнительных факторов, которые могут усложнить эту основную тему смены поколений. Наиболее заметным осложнением является пол. Как спорофиты, так и гаметофиты могут иметь пол. Кроме того, спорофит или гаметофит могут не быть полностью независимыми от другого поколения. Для примера рассмотрим цветущие растения. Некоторые цветы содержат как мужские, так и женские гаметофиты. У других видов цветов мужские и женские цветки разделены, но могут присутствовать на одном и том же растение, В других цветах отдельные спорофиты дают только определенный пол цветка.
Термины моногенный и диоидный используются для описания гаметофитов. Если оба пола содержатся в одном и том же особе, этот вид моноусен. Если они разделены, то чередование поколений двоякое. Это относится только к гаметофиту.
В спорофите используемые термины являются однодомными и двудомными. Здесь они описывают, будет ли спорофит продуцировать оба пола, или же пол разделен на разные спорофиты. Важно отметить, что многие растения однодомные, но двудомные. Это означает, что один спорофит может производить как мужские, так и женские цветы. Многие другие комбинации этих различий в чередовании поколений можно наблюдать у различных видов растений и водорослей.
Различия в спорофите и гаметофите
При смене поколений у некоторых видов наблюдается неравномерное распределение по размеру или доминированию либо спорофита, либо гаметофита. У разных видов это можно изменить. В печеночники гаметофит является доминирующим поколением. Печеночник – гаметофитное растение. У папоротников верно обратное, и спорофит – более крупный и доминирующий особь. Это можно назвать спорофитным растением.
У других видов размер и форма спорофита и гаметофита практически неразличимы. Разница лишь в количестве ДНК, которую они несут, и в том, будут ли они производить споры или гаметы. Хотя может показаться, что они на самом деле ничего не делают, смена поколений все же позволяет половому размножению происходить. Это сильно смешивает гены и повышает адаптивность вида.
Спорофиты и гаметофиты разных видов также продуцируют разные типы спор и гамет. Если гаметы одинаковы, это называется изогамия. Некоторые виды зеленых водорослей производят только один тип гаметы. Они оба жгутиковые и плавают в открытой воде, пока не найдут друг друга. Эта схема не особенно полезна для наземных растений. Большинство наземных растений показывают анизогамию или разницу в их гаметах. Это чаще всего рассматривается как разница между подвижной мужской гаметой и более крупной, неподвижной женской гаметой, или спермой и ооцитами. В других случаях это рассматривается только как разница в размерах.
Ниже приведено графическое изображение двудомного двудомного анизогаметного растения. Это означает, что у растения есть отдельные особи как гаметофитов, так и спорофитов, и что производимые им гаметы имеют разные размеры. Это было бы верно для дерева падуба или ивы, которое производит отдельные мужские и женские цветы на отдельных отдельных спорофитах.

Эволюция смены поколений
Ископаемые свидетельства и существование многих водорослей с упрощенным чередованием жизненных циклов поколений намекают на то, что эволюционное преимущество полового размножения посредством смены поколений – это то, что сделало земные растения достаточно приспособленными для колонизации земли. Чередование поколений учитывает как динамичный и изменчивый акт полового размножения, так и устойчивый и последовательный акт бесполое размножение.
Когда спорофит создает споры, клетки подвергаются мейоз что позволяет поколению гаметофитов рекомбинировать генетика подарок. Это допускает большое разнообразие. Поскольку растения колонизировали землю, они были изначально изоморфны, или гаметофиты и спорофиты выглядели и действовали примерно одинаково. Со временем большинство растений сочли целесообразным сократить один из этих жизненных циклов. У большинства цветковых растений в настоящее время значительно сокращен жизненный цикл гаметофита, в то время как печеночники и мхи пошли другим путем, предпочитая уменьшить цикл спорофитов. В большом разнообразии растений, живущих сегодня, есть почти все возможные варианты смены поколений.
викторина
1. Каково одно преимущество смены поколений перед бесполым видом?A. Бесполых видов очень мало генетическая рекомбинация B. Нет никаких преимуществC. Легче воспроизвести
Ответ на вопрос № 1
верно. В то время как размножение намного сложнее, чередование поколений позволяет рекомбинировать и тестировать гены так, как это позволяют немногие системы размножения.
2. В чем разница между сменой поколений у растений и половым размножением у млекопитающих?A. Млекопитающие чередуются только с каждым другим поколениемB. Млекопитающие не чередуют поколенияC. Млекопитающие производят сперму и яйца, где растения не
Ответ на вопрос № 2
В верно. Млекопитающие одинаковы для каждого поколения и не различаются между гаплоидными и диплоидными государствами. Некоторые животные, такие как пчелы, очень похожи между гаплоидом и диплоидом, но это определяет их пол в поколении и не меняется в течение нескольких поколений. У растений эта линия становится размытой по мере того, как гаметофит становится зависимым от спорофита, как это видно у многих цветковых растений.
3. Лучше ли иметь более доминирующий спорофит или более доминирующий гаметофит?A. СпорофитB. гаметофитаC. Ни!
Ответ на вопрос № 3
С верно. Доминирование определенного поколения полностью зависит от успехов и неудач прошлых поколений вида. Если спорофит имеет тенденцию быть слабее или более ранним, он станет менее заметным признаком в репродуктивном цикле. То же самое относится и к гаметофиту. Каждый вид сформировал жизненный цикл, который позволил ему дожить до настоящего времени, поэтому все по существу равны.
Воспроизведение — это способность организмов образовывать себе подобных. Воспроизведение является одним из важнейших свойств жизни и возможно благодаря общей способности организмов производить потомство. Однако не всегда непосредственные потомки подобны родительским особям. Например, из спор папоротника вырастает многочисленное потомство, представленное заростками, не похожими на материнское спороносное растение. На заростке, в свою очередь, возникает непохожее на него растение — спорофит. Такое явление получило название чередование поколений.
Если образование потомства сопровождается увеличением числа особей данного вида, то такой процесс представляет собой размножение. Размножение — это воспроизведение генетически сходных особей данного вида, которое характеризуется увеличением числа особей в дочернем поколении по сравнению с поколением родителей.
При размножении обеспечивается преемственность и непрерывность жизни. Преемственность заключается в том, что в процессе воспроизведения передается вся генетическая информация, заложенная в родительском поколении, дочернему поколению. Благодаря смене поколений определенные виды и их популяции могут существовать неограниченно долго, так как снижение их численности вследствие естественной гибели особей компенсируется за счет постоянного воспроизведения организмов и замещения умерших родившимися(непрерывность жизни).
Виды организмов, будучи представлены смертными особями, благодаря смене поколений не только сохраняют и передают потомкам основные черты своего строения и функционирования, но и изменяются. Наследственные изменения организмов в размножения наблюдается у некоторых водорослей и простейших (фораминифер, споровиков).
Бесполое размножение многоклеточных организмов. Вегетативное размножение— форма бесполого размножения у растений, при котором начало новому организму дают вегетативные органы — корень, стебель,лист, либо специализированные видоизмененные побеги — клубни, луковицы, корневища, выводковые почки и т. п.
В основе фрагментации, как и в случае вегетативного размножения, лежит способность организма восстанавливать недостающие органы и части тела (регенерация). При этом способе размножения новые особи возникают из фрагментов материнского организма. Фрагментацией могут размножаться, например, нитчатые водоросли, грибы, некоторые плоские (ресничные) и кольчатые черви.
Почкование характерно для губок, некоторых кишечнополостных (гидры) и оболочников (асцидии), у которых за счет размножения группы клеток на теле образуются выпячивания (почки). Почка увеличивается в размерах, затем у нее появляются зачатки всех структур и органов, характерных для материнского организма. Потом происходит отделение (отпочковывание) дочерней особи, которая растет и достигает размеров материнского организма. Если дочерние особи не отделяются от материнской, то формируются колонии (коралловые полипы).
У некоторых кишечнополостных встречается размножение стробиляцией. При этом полип интенсивно растет, а затем в верхней части делится поперечными перетяжками на дочерние особи (стробилы). В это время полип напоминает стопку тарелок. Образовавшиеся дочерние особи — медузы — отрываются от материнской и начинают самостоятельное существование.
Бесполое размножение одноклеточных организмов. Бактерии и простейшие (амебы, эвглены, инфузории и др.) размножаются делением клетки надвое. Бактерии делятся простым бинарным делением; простейшие — митозом. После деления дочерние клетки растут и, достигнув величины материнского организма, снова делятся.
В природе наблюдаются случаи, когда клетки делятся не на равные части. В этом случае меньшая клетка как бы отпочковывается от большой. Подобный тип деления (гетеротомия) встречается у дрожжей и некоторых бактерий, и его называют почкованием.
Множественное деление (шизогония) характеризуется тем, что при таком размножении наблюдается многократное деление ядра без деления цитоплазмы. Далее вокруг каждого из ядер обособляется небольшой участок цитоплазмы, и деление клетки завершается образованием множества дочерних особей. Такой тип ряду поколений приводят к изменению вида или к возникновению новых видов.
Обычно различают два основных типа размножения: бесполое и половое. Половое размножение связано с образованием половых клеток — гамет, их слиянием (оплодотворением), образованием зиготы и дальнейшим ее развитием. Бесполое размножение не связано с образованием гамет.
Половое размножение
Половой процесс. Половое размножение отличается наличием полового процесса, который обеспечивает обмен наследственной информацией и создает условия для возникновения наследственной изменчивости. В нем, как правило, участвуют две особи — женская и мужская, которые образуют гаплоидные женские и мужские половые клетки — гаметы. В результате оплодотворения, т. е. слияния женской и мужской гамет, образуется диплоидная зигота с новой комбинацией наследственных признаков, которая и становится родоначальницей нового организма.
Половое размножение по сравнению с бесполым обеспечивает появление наследственно более разнообразного потомства. Формами полового процесса являются конъюгация и копуляция.
Конъюгация — своеобразная форма полового процесса, при которой оплодотворение происходит путем взаимного обмена мигрирующими ядрами, перемещающимися из одной клетки в другую по цитоплазматическому мостику, образуемому двумя особями. При конъюгации обычно не происходит увеличения количества особей, но происходит обмен генетическим материалом между клетками, что обеспечивает перекомбинацию наследственных свойств. Конъюгация типична для ресничных простейших (например, инфузорий), некоторых водорослей (спирогиры).
Копуляция (гаметогамия) — форма полового процесса, при которой две различающиеся по полу клетки — гаметы — сливаются и образуют зиготу. При этом ядра гамет образуют одно ядро зиготы.
Различают следующие основные формы гаметогамии: изогамия, анизогамия и оогамия.
При анизогамии (гетерогамии) формируются подвижные, различающиеся морфологически и физиологически гаметы. Такой тип полового процесса характерен для многих водорослей.
В случае оогамии гаметы сильно отличаются друг от друга. Женская гамета — крупная неподвижная яйцеклетка, содержащая большой запас питательных веществ. Мужские гаметы — сперматозоиды —- мелкие, чаще всего подвижные клетки, которые перемещаются с помощью одного или нескольких жгутиков. У семенных растений мужские гаметы — спермии — не имеют жгутиков и доставляются к яйцеклетке с помощью пыльцевой трубки. Оогамия характерна для животных, высших растений и многих грибов.
Гаметогенез. Процесс образования и развития гамет называется гаметогенезом. У многоклеточных водорослей, многих грибов и высших споровых растений формирование гамет происходит в специальных органах полового размножения — гаметангиях. У высших споровых растений женские гаметангии называются архегониями, мужские — антеридиями. У животных гаметогенез протекает в специальных половых железах —гонадах. Однако, например, у губок и кишечнополостных половые железы отсутствуют и гаметы возникают из различных соматических клеток.
Особую форму полового размножения представляет собой партеногенез, или девственное размножение, — развитие организма из неоплодотворенной яйцеклетки. Данная форма размножения характерна главным образом для видов, обладающих коротким жизненным циклом с выраженными сезонными изменениями.
Партеногенез бывает гаплоидным и диплоидным.
При гаплоидном (генеративном) партеногенезе новый организм развивается из гаплоидной яйцеклетки. Получаемые при этом особи могут быть только мужскими, только женскими или теми и другими. Это зависит от хромосомного определения пола. Например, у пчел, паразитических ос, муравьев в результате партеногенеза из неоплодотворенных яиц появляются самцы, в то время как из оплодотворенных —диплоидные самки, что приводит к возникновению различных каст организмов. Такой механизм размножения у общественных насекомых позволяет регулировать численность потомков каждого типа.
У тлей, дафний, коловраток, некоторых ящериц наблюдается диплоидный (соматический) партеногенез, при котором ооци-ты самки формируют диплоидные яйцеклетки. Например, у дафний самки диплоидны, а самцы гаплоидны. В благоприятных условиях у дафний не происходит мейоза: диплоидные яйцеклетки развиваются без оплодотворения и дают начало самкам. У скальных ящериц перед мейозом происходит митотическое увеличение числа хромосом в клетках половых желез. Далее клетки проходят нормальный цикл мейоза, и в результате образуются диплоидные яйцеклетки, которые без оплодотворения дают начало новому поколению, состоящему только из самок. Это позволяет поддерживать численность особей в условиях, когда затруднена встреча особей разного пола.
Установлено существование естественного партеногенеза у птиц. У одной из пород индеек яйца часто развиваются партено-генетически, причем из них появляются только самцы.
У многих видов партеногенез носит циклический характер. У тлей, дафний, коловраток в летнее время существуют лишь самки, развивающиеся партеногенетически, а осенью партеногенез сменяется размножением с оплодотворением. Циклический партеногенез исторически развивался у видов животных, которые погибают в большом количестве. Широко распространён партеногенез у паразитических червей, находящихся на личиночных стадиях развития, что обеспечивает их интенсивное размножение и выживание, несмотря на массовую гибель на различных этапах жизненного цикла.
Партеногенез можно вызвать искусственно. Советский Учёный Б. Л. Астауров опытным путем (раздражая разными способами поверхность яиц тутового шелкопряда: механически поглаживая кисточкой или делая уколы иглой, химически помещая яйца в различные кислоты, термически — нагревая яйца) добился эффекта дробления яиц без оплодотворения. В дальнейшем американец Грегори Пинкус из неоплодотворен ных яиц партеногенетически получил взрослых лягушек и кроликов-
В природе естественный партеногнез встречается у ряда растений (одуванчик, ястребинка и др.) и носит название апомиксис Под апомиксисом обычно понимают либо развитие из неопло-дотворенной яйцеклетки, либо возникновение зародыша вообще не из гамет (например, у цветковых растений зародыш может развиваться из различных клеток зародышевого мешка)
Чередование поколений.
У преобладающего большинства как высших, так и низших растений существует 2 способа размножения: бесполое и половое. Причем у каждого растения полный жизненный цикл его развития возможен только при наличии обоих способов размножения, которые осуществляются в определенной очередности: один способ размножения чередуется с другим, и, следовательно, происходит определенное чередование, или смена, поколений — бесполого и полового.
Сущность чередования поколений в цикле развития одного растения заключается в том, что одно поколение образует органы полового размножения, а другое поколение — органы бесполого размножения. Поколение, образующее органы полового размножения, в которых формируются половые клетки — гаметы, называется половым поколением, или гаметофитом. Поколение, на котором образуются органы бесполого размножения, с развивающимися в них спорами, называется бесполым, или спорофитом.
В процессе эволюции у разных групп растений гаметофит и спорофит формировались неодинаково, а поэтому в растительном мире существуют большие различия в морфологическом строении таких поколений. У одних растений оба поколения развиты одинаково, внешне довольно сходны и живут самостоятельно (многие водоросли). У других растений гаметофит и спорофит внешне сильно различаются. Так, например, у мхов гаметофит развит сильнее и морфологически более дифференцирован, чем спорофит, но существуют они не самостоятельно: спорофит живет (паразитирует) на гаметофите (мох кукушкин лен).
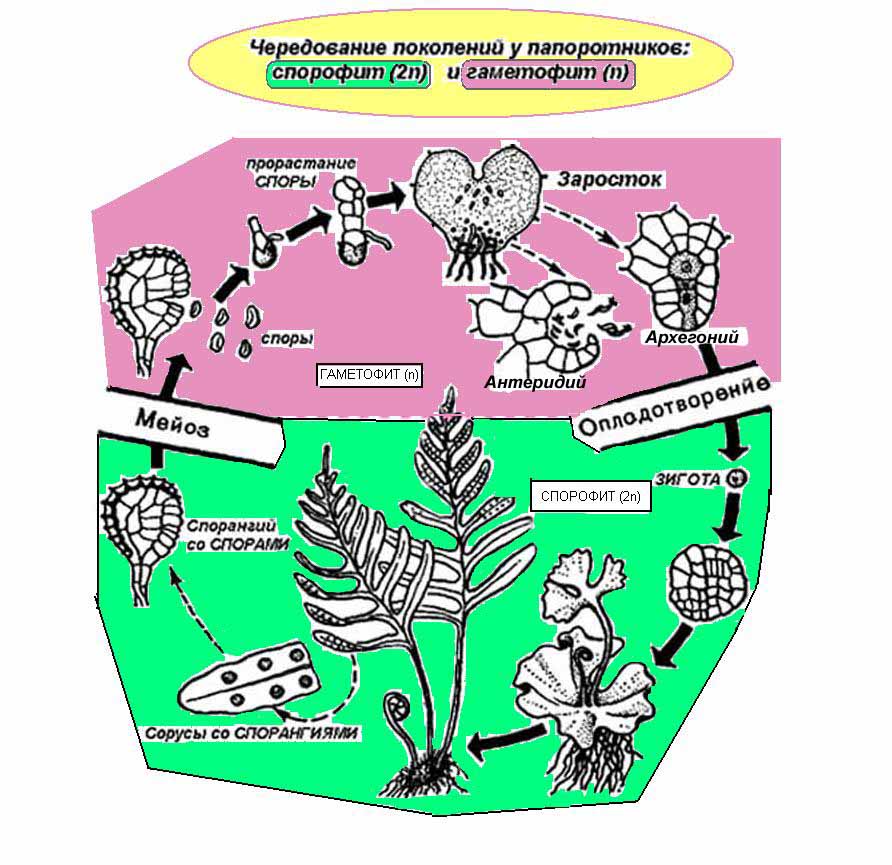
Очень ясно выражено развитие двух поколений у папоротников. У них оба поколения живут самостоятельно, причем у некоторых папоротников спорофит по размерам резко отличается от гаметофита (мужской папоротник). Бесполое поколение у этого папоротника представляет собой крупное растение, хорошо дпфферинцированное на отдельные органы, достигающие 80. 100 см и больше, а половое поколение (гаметофит) — очень маленькое растеньице в виде зеленой пластинки с 10-копеечную монету.
Процесс чередования поколений мужского папоротника проходит следующим образом. На нижней стороне листа спорофита образуются специальные органы (спорангии), в которых формируются споры. Созревшие споры высыпаются и, попав в благоприятные условия, прорастают.
Из споры вырастает маленькая зеленая пластинка, которая и представляет собой половое поколение, или гаметофит, папоротника. В данном случае это половое поколение носит специальное название — заросток. С нижней стороны заростка образуются мужские (антеридии) и женские (архегонии) половые органы. В антеридиях формируются мужские гаметы -сперматозоиды, в архегониях образуются женские гаметы — яйце –клетки.
После оплодотворения из образовавшейся зиготы развивается бесполое поколение папоротника — спорофит, т. е. вырастает обычное растение папоротника, на листьях которого снова образуются споры. Жизненный цикл папоротника начинается снова.
Таким образом, у мужского папоротника бесполое и половое поколения растут отдельно и питаются самостоятельно.
У цветковых, которые эволюционно стоят выше рассмотренных растений, чередование поколений также существует, но оно выражено менее наглядно, так как у этих растений гаметофиты сильно редуцированы. У цветковых растений имеется 2 гаметофита — мужской (двуклеточное пыльцевое зерно) и женский (зародышевый мешок, содержащий 7 клеток). У этих растений гаметофит живет на бесполом поколении, которое представляет собой целое растение, достигающее у некоторых представителей огромных размеров (береза, дуб и др.), тогда как женский и мужской гаметофиты у цветковых растений имеют микроскопически малую величину.
Одновременно с чередованием поколений происходит смена ядерных фаз. Осуществляется это следующим образом. На бесполом поколении споры образуются в спорангиях из спорогенной ткани. При образовании спор происходит редукционное деление, и споры, следовательно, имеют гаплоидный набор хромосом. Образовавшийся из споры гаметофит — половое поколение и сформировавшиеся на нем гаметы также гаплоидны. При слиянии гаплоидных гамет на половом поколении образуется зигота, которая несет уже диплоидный набор хромосом. Гаплоидная фаза ядра сменилась диплоидной фазой. Из зиготы развивается бесполое поколение — спорофит — с диплоидным набором хромосом. Затем при образовании спор число хромосом снова уменьшается вдвое. Таким образом, гаметофит и спорофит различаются не только внешне, по и цитологически: они имеют различное число хромосом.
У более древних примитивных растений в цикле развития преобладает половое поколение — гаметофит (гаплоидная фаза). Более высокоорганизованным растениям, наоборот, свойственно при чередовании поколений преобладание бесполого поколения — спорофита (диплоидной фазы), что особенно ясно выражено у цветковых растений, у которых половое поколение претерпело сильную редукцию.

Рис. Чередование поколений у папоротника щитовника мужского:
/ - спорофит; 2 — часть листа с сорусами; 3.— сорус в разрезе; 4 — клетка, спорогенной ткани; 5 — редукционное деление; 6 — 2-е деление; 7 — тетрада; 8— споры; 9 — раскрывшийся спорангий; 10 — прорастающая спора; // — заросток (гаметофит); 12 — архегоний; /Л — антеридий; 14 —сперматозоид; 15 — проникновение сперматозоида в архегоний; 16 — деление зиготы; 17 — заросток с проростком молодого папоротника
Чередование поколений имеет большое биологическое значение, так как в нем сочетается 2 способа размножения: бесполое, дающее большое число особей, и половое, способствующее обогащению наследственности потомства.

По названию этой статьи не всем даже понятно о чем пойдет речь, не правда ли?
К тому же, судя по ответам учащихся на ЕГЭ, именно этот вопрос остается вовсе без ответа.
Есть ли чередование поколений у растений
Давайте вспомним, как называются эти стадии. Спорофит и гаметофит. Почему они так называются?
Теперь, относительно того, что надо знать про эту другую часть жизненного цикла растения, названную гаметофитом.
Таким образом, у растений не только гаметы (половые клетки), являются гаплоидными (n), но и бесполые клетки — споры , тоже являются гаплоидными.
Почему же тогда споры — это бесполые клетки, а гаметы — половые клетки
Каждая гаплоидная спора (одна) не сливаясь ни с какой другой клеткой, то есть сама по себе, прорастая, образует новый организм (вернее другую жизненную стадию организма), генетически идентичную наследственному аппарату этой одной споры.
Таким образом, спора, являясь продуктом спорофита, сама образует будущий гаметофит. Такое размножение и называется бесполым.
Ткани гаметофита гаплоидные (они же развились из гаплоидных спор), из них формируются гаметы. Каждая гаплоидная гамета не образует новый организм . Только после стадии оплодотворения её другой гаметой, после объединения генетического материала (n) женской и (n) мужской гамет, образуется диплоидная (2n) зигота. Именно эта диплоидная зигота и даст начало новому будущему диплоидному организму (спорофиту).
Таким образом, гаметы, являясь продуктом гаплоидного гаметофита, только сливаясь попарно (мужские с женскими) обеспечат дальнейшее развитие организма. Поэтому такое размножение, в котором участвуют два партнера называется половым.
Что же является спорофитом и гаметофитом у споровых (водоросли, мхи и папоротниковидные) и семенных растений (голосеменные и покрытосеменные)
Мы подошли к ответу на вопрос, который вызывает наибольшую путаницу. Так вот, у водорослей и мхов, основным (доминирующим) поколением в цикле развития является гаметофит. А у папоротниковидных (хотя они тоже относятся к споровым растениям ) и всех семенных растений основным поколением является спорофит.
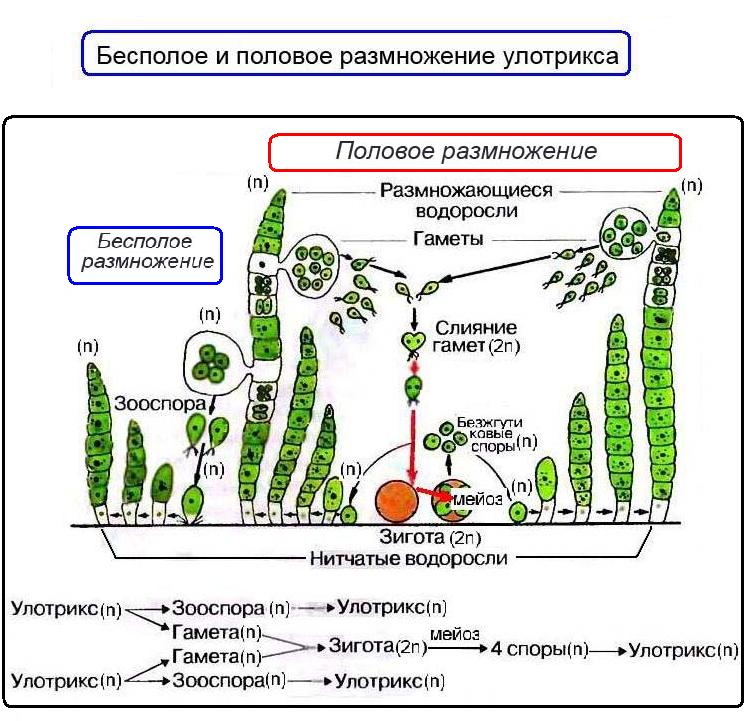
Цикл чередования поколений у водорослей разберем на примере нитчатой зеленой водоросли улотрикса. На рисунке из школьного учебника мы видим, что улотрикс может размножаться как бесполым, так и половым путем. Значит взрослое растение улотрикса можно считать спорогаметофитом. При благоприятных условиях улотрикс (n) размножается бесполым путем четырехжгутиковыми зооспорами (n) . При неблагоприятных условиях улотрикс (n) размножается половым путем, образуя двухжгутиковые гаметы (n) . После копуляции (слияния) гамет образуется четырехжгутиковая зигота (2n) .
Зигота вначале плавает, затем оседает на дно, теряет жгутики, вырабатывает плотную оболочку и слизистую ножку, которой прикрепляется к субстрату. Это покоящийся спорофит.
После периода покоя происходит редукционное деление ядра зиготы (мейоз) и в ней образуются или безжгутиковые споры (n), или зооспоры (n) , что зависит от вида улотрикса (а их 25 видов), Из этих спор (или зооспор) снова формируются взрослые растения улотрикса — спорогаметофиты.

У мха кукушкин лен ,
взрослое вегетирующее растение является гаметофитом (n), образующимся из зеленой нити — протонемы (предростка) — (n).
Кукушкин лен — раздельнополое растение. На рисунке показано, что после оплодотворения (n + n), на женском гаметофите формируются коробочки со спорами (2n).
Коробочка на ножке — это стадия спорофита в цикле развития кукушкина льна. Споры в коробочках формируются в результате мейоза. Затем уже гаплоидные споры (n) высыпаются из коробочки наружу и из них образуется зеленая нить — протонема (п) .
Таким образом, мы видим, что у мхов как и у водорослей в цикле чередования поколений стадия гаметофита является преобладающей над спорофитом.
А у папоротников и всех семенных растений их основной жизненной формой, самим вегетирующим растением является спорофит
На рисунке ниже показана схема изменения соотношения гаметофита (n) и спорофита (2n) в процессе эволюции растений, Красная линия, разделяет изображения спорофитов (выше линии) и гаметофитов (ниже линии) у разных групп растений.
На рисунке мы видим, что только у водорослей и мхов стадия гаметофита (n) является преобладающей. У папоротников гаметофит представлен маленьким заростком, а у голосеменных и покрытосеменных вообще редуцирован до микроскопических размеров.
Казалось бы, поскольку папоротники как и мхи споровые растения, то у них чередование поколений должно происходить сходным со мхами образом. Но оказывается все наоборот: у споровых папоротников цикл чередования поколений (имеется в виду, какая форма представляет собой само взрослое вегетирующее растение) сходен с циклом чередования поколений у семенных растений.
Что бы этот факт легче запомнился, следует указать, что мхи — тупиковая ветвь эволюции царства растений. И, что именно от папоротниковидных произошли все современные семенные растения (только семенные растения произошли не от ныне живущих споровых папоротников, а от вымерших папоротников, у которых уже было семенное размножение).
Есть ли чередование поколений у животных
Да, есть. Но, если чередование поколений характерно почти для всех представителей царства растений, то в царстве животных это скорее исключение, чем правило.
Из курса школьной программы по биологии надо помнить, что чередование поколений есть у некоторых паразитических простейших (например, у малярийного плазмодия — тип споровики), многих кишечнополостных, паразитических червей (тип плоские черви) и некоторых насекомых.
А у растений то, помните, взрослое растение — спорофит, хотя и тоже 2n, но представляет собой бесполое поколение).
Половые клетки (n) после оплодотворения образуют зиготу-6 снова (2n), развивающуюся в личинку — плавающую планулу-7.
Планула оседает на дно и из нее образуется новый организм, совершенно не похожий на медузу — сидячий полип-1,2 (тоже как и планула и медуза 2n).
Основной и промежуточный хозяин
В цикле чередования поколений паразитарных животных (то есть живущих внутри других организмов) смена их жизненных фаз сопровождается сменой хозяев.
Например, у печеночного сосальщика, то животное, в котором происходит обычное половое размножение паразита, называется основным хозяином (человек, парнокопытные). А животное, в котором осуществляется партеногенетическое размножение паразита называется промежуточным хозяином (моллюск — малый прудовик).
В заключение хочется еще раз подчеркнуть
* у животных организмов, для которых характерно развитие с чередованием поколений, оба поколения и половое, и бесполое состоят из клеток с двойным набором хромосом (2n);
* у высших же растений (кроме мхов) их взрослая вегетирующая форма, являющаяся спорофитом — бесполым поколением, тоже содержит в своих клетках двойной набор хромосом (2n), а гаметофит — половое поколение — всегда гаплоиден (n).
Недавно в комментариях прозвучал вопрос от Александра:
Ответ на этот интересный вопрос посчитал уместным разместить в конце этой статьи.
Известно, что чередование поколений зависит от условий среды. При благоприятных условиях размножение происходит, как правило, бесполыми способами – делением, почкованием, вегетативно. При неблагоприятных условиях бесполое поколение сменяется половым.
Так произошло исторически, что эволюция размножения шла от бесполого размножения, свойственного одноклеточным, к половому размножению. От организмов с гаплоидным числом хромосом в клетках – к организмам с диплоидным набором хромосом.
Согласитесь, что диплоидность – это возможность обладать более разнообразной генетической информацией. Помимо мутационной изменчивости обладать возможностями комбинативной изменчивости, а значит и возможностью иметь эволюционные преимущества.
Примитивные формы, размножаются только бесполым путем, а у более сложных форм бесполое размножение чередуется с половым (в основном в царстве растений). В процессе эволюции в цикле развития организмов закономерно уменьшается роль (продолжительность существования и размеры) гаплоидной фазы и увеличивается роль диплоидной фазы.
У кого есть вопросы по статье к репетитору ЕГЭ по биологии по Скайпу, замечания, пожелания — прошу в комментарии.
У меня на блоге вы можете приобрести ответы на все тесты ОБЗ ФИПИ за все годы проведения экзаменов по ЕГЭ и ОГЭ (ГИА).
Чередование поколений — закономерная смена у организмов поколений, различающихся способом размножения. Организмы многих видов могут размножаться как бесполым, так и половым путем. В связи с этим говорят о бесполом и половом поколениях данного вида.
Чередование этих поколений у растений и животных имеет много общих черт. Граница, разделяющая половое и бесполое поколения в цикле развития, — процесс оплодотворения (рис. 1).

Рис. 1. Эволюция размножения. А — только бесполое размножение; Б, В, Г — чередование бесполого и полового поколений; Д — только половое размножение.
При этом в результате слияния гаплоидных (т. е. содержащих одинарный набор хромосом) гамет появляется диплоидная (т. е. содержащая двойной набор хромосом) зигота, и половое поколение переходит в бесполое. И бесполое, и половое поколения могут иметь как одинарный, так и двойной набор хромосом: в зависимости от того, на какой стадии жизненного цикла происходит мейоз. При мейозе число хромосом уменьшается вдвое и диплоидный их набор переходит в гаплоидный. Мейоз и оплодотворение — это две вехи, разделяющие гаплоидную и диплоидную фазы в цикле развития.
В процессе эволюции в цикле развития закономерно уменьшается роль (продолжительность существования и размеры) гаплоидной фазы и увеличивается роль диплоидной фазы.
У споровиков и жгутиковых, многих водорослей и некоторых грибов диплоидная фаза представлена только зиготой, которая сразу претерпевает мейоз, образуя гаплоидные клетки (рис. 1, б и 2). У всех высших и некоторых низших форм (отдельные водоросли и грибы, инфузории) зигота делится путем митоза, поэтому бесполое поколение у них так же, как и зигота, диплоидное.

Рис. 2. Зеленая водоросль улотрикс: 1 — вегетативная нить; 2 — бесполое размножение водоросли при помощи зооспор; 3 — половое размножение путем образования гамет и слияния двух гамет в зиготу; 4 — зооспоры; 5 — гаметы; 6 — слияние гамет; 7 — зигота.
Так, у фораминифер из зиготы вырастает диплоидное бесполое поколение. В результате мейоза из него образуются гаплоидные клетки, из которых вырастает также гаплоидное половое поколение. Половое поколение в результате многократного деления ядра образует гаметы, которые, сливаясь попарно, дают зиготу (рис. 1, в). Процесс бесполого размножения у мохообразных, папоротникообразных и некоторых других растений происходит в результате рассеивания гаплоидных спор, возникающих при мейозе (рис. 1, в и 3). У таких видов процесс мейоза отделяет бесполое поколение (спорофит) от полового (гаметофит).

Рис. 3. Цикл развития мохообразных: 1 — гаметофит; 2 — спорофит; 3 — спорангий; 4 — спора; 5 — молодой гаметофит; 6 — антеридий; 7 — архегоний; 8 — яйцеклетка; 9 — сперматозоид.
Споры делятся митотически, образуя гаплоидное половое поколение.
У кишечнополостных и других многоклеточных животных происходит дальнейшее подавление гаплоидной фазы (рис. 1, г). У них диплоидно как бесполое, так и половое поколение, которое образуется из бесполого путем митотического деления его клеток. Мейоз происходит только в процессе образования гамет, которые являются единственной гаплоидной фазой у таких организмов. Например, гидроидные полипы представляют собой бесполое поколение. Почкуясь, они образуют колонии, на которых развиваются медузы с семенниками и с яичниками (диплоидное половое поколение).
Рис. 4. Цикл развития покрытосеменных: 1 — мужской гаметофит; 2 — женский гаметофит; 3 — яйцеклетка; 4 — пыльцевое зерно; 5 — молодой спорофит; 6 — эндосперм; 7 — семядоля; 8 — мейоз; 9 — микроспоры; 10 — макроспоры.
Медузы свободно плавают в воде и размножаются половым путем. В результате опять возникают полипы (рис. 5).

Рис. 5. Жизненный цикл гидроидного полипа: 1 — гидроиды й полип; 2 — образование медуз с семенниками и яичниками путем бесполого размножения — почкования; 3 — яйцеклетки и спермин; 4 — зигота; 5 — развитие новой колонии полипов.
У животных различают первичное и вторичное чередование поколений. При первичном чередуется бесполое и половое размножение. Так бывает у многих простейших. К вторичному чередованию поколений относят метагенез и гетерогонию. При метагенезе, который характерен для оболочников и кишечнополостных, чередуется половое и вегетативное размножение. При гетерогонии, которая характерна для трематод, некоторых круглых червей и коловраток, ряда членистоногих, чередуются нормальное половое размножение с партеногенезом.
Чередование поколений зависит от условий среды. При благоприятных условиях размножение происходит, как правило, бесполыми способами — делением, почкованием, вегетативно или партеногенетически. При неблагоприятных условиях бесполое поколение сменяется половым.
Эволюция размножения шла от бесполого, свойственного одноклеточным, к половому. Примитивные формы размножаются только бесполым путем, у более сложных форм бесполое размножение чередуется с половым. Наиболее прогрессивные виды размножаются только половым путем (рис. 1).
Чередование поколений у растений. Типичное чередование поколений характерно для растений, у которых многоклеточны как диплоидная фаза (диплонт), так и гаплоидная (гаплонт). Диплонт образует спорангии, в которых в результате мейоза возникают споры (поэтому диплонт называют также спорофитом или бесполым поколением). Гаплонт образует гаметангии, в которых без редукционного деления — мейоза образуются половые клетки — гаметы (гаплонт называют также гаметофитом или половым поколением).
Спорофит развивается из зиготы, возникающей в результате оплодотворения, т. е. слияния двух гамет, а гаметофит — из споры. У немногих растений (например, у некоторых зелен«ых и бурых водорослей) спорофит и гаметофит развиты одинаково, а у большинства растений в циклах развития преобладает либо гаметофит (например, у мохообразных), либо спорофит — бурая водоросль ламинария, папоротникообразные и семенные растения (рис. 6).
У многих зеленых водорослей (хламидомонада, улотрикс, спирогира и др.) диплоидны только зиготы, при прорастании которых происходит мейоз (рис. 6). А у сифоновых, диатомовых и некоторых бурых водорослей, как и у огромного большинства животных, гаплоидны только гаметы, возникающие в результате мейоза. Поэтому у этих растений фактически чередования поколений нет, хотя«смена ядерных фаз происходит.

Рис. 6. Смена ядерных фаз у растений: 1 — бурая водоросль диктиота (гаметофит и спорофит развиты одинаково); 2 — мох кукушкин лен (доминирует гаметофит); 3 — папоротник (доминирует спорофит); 4 — зеленая водоросль спирогира (диплоидны лишь зиготы); 5 — бурая водоросль фукус (гаплоидны лишь гаметы). ГАМ — гаметы, ОПЛ — оплодотворение, ЗИГ — зигота, МЕЯ — мейоз; МСП — мейоспоры, СПФ — спорофит, СП Г — спорогоний (спорофит мохообразных), ГМФ — гаметофит, ЗАР — заросток (гаметофит папоротникообразных и семенных растений).
Фазы в циклах развития высших растений имеют особые названия: спорофиты мохообразных называют спорогониями (они развиваются на гаметофитах), а гаметофиты остальных высших растений — заростками (рис. 6). У папоротникообразных они существуют самостоятельно, а у семенных развиваются на спорофитах. Заростки равноспоровых растений (см. Споры) обоеполы, а разноспоровых — раздельнополы и более редуцированы (особенно мужские), чем заростки равноспоровых. Так, например, у покрытосеменных растений мужской заросток — это развивающееся из микроспоры пыльцевое зерно, а женский заросток — зародышевый мешок, развивающийся из мегаспоры.
Читайте также: